
长三角地区粳稻种质的稻瘟病抗性基因鉴定及其抗性评价
2022-03-11 04:48周锋利张丽霞胡泽军曹黎明吴书俊
核农学报 2022年1期
闫 影 王 凯 周锋利 张丽霞 胡泽军 曹黎明,2 吴书俊,2,*
(1 上海市农业科学院作物育种栽培研究所,上海 201403;2 上海农产品保鲜加工工程技术研究中心, 上海 201403;3 上海市嘉定区农业技术推广服务中心,上海 201800)
稻瘟病是由子囊真菌(Magnaportheoryzae)引起的一种水稻真菌病害,是水稻生产中流行最广、危害最大的世界性水稻三大病害之一,素有“水稻癌症”之称[1-2]。依据稻瘟病感染发病部位的不同可将其分为苗瘟、叶瘟、节瘟、穗颈瘟、谷粒瘟等,其中穗颈瘟的危害最大,发病严重时会造成水稻减产40%~50%,甚至颗粒无收[3-4]。近年来我国稻瘟病年均发生面积在470万hm2次左右;2013年以后,发病情况更呈现上升趋势[3]。
当前全国各地在新品种审定时已实施对稻瘟病表现感病的“一票否决制”,对育成品种的稻瘟病抗性提出了更高要求[5]。研究表明,选育推广具有持久和广谱抗性的水稻新品种是防治稻瘟病最有效的方法之一,而抗病基因的发掘与利用是抗病育种的基础和核心[6]。20世纪60年代中期,日本率先开展了水稻品种抗稻瘟病基因分析的研究工作,鉴定出最初8个抗性位点上的14个基因,并建立了一套抗稻瘟病基因分析用的鉴别体系(japanse differential cultivars,JDCs),随后,国际水稻研究所和中国等产稻国也逐渐开展了水稻稻瘟病抗性遗传的系统性研究[7]。截至目前,已鉴定的抗稻瘟病质量性状基因有100多个,数量性状位点500多个[8],Pib[9]、Bsrd-1[10]、pi-21[11]、Pi2[12]、Pi9[13]、Piz-t[12]、Pigm[14]、Pi5/Pi3/Pii[15]、Pikh/Pi54[16]、Pikm[17]、Pik[18]和Pita[19]等37个抗稻瘟病基因获得克隆。其中,Piz-t、Pi9、Pi2和Pigm互为Piz位点上的等位基因[14];Pi-1、Pikh/Pi-54、Pik-p和Pikm互为Pik位点上的等位基因[18];Pi-25和Pid-3互为等位基因[20];PiCO39和Pia互为等位基因[21]。部分已克隆的抗病基因已在育种中广泛应用[22],例如李莹[23]利用分子标记辅助技术,将广谱、高抗的稻瘟病基因Pid2和Pid3导入到空育131的遗传背景中,以培育新的寒区水稻品系空育131(Pid2/Pid3);马作斌等[24]利用分子标记辅助选育出聚合基因Pi5和Pita的抗稻瘟病水稻新品系。
本研究利用8个抗稻瘟病基因功能标记对长三角地区52份粳稻资源进行抗稻瘟病基因检测,结合田间抗病评价,分析其对稻瘟病的抗性贡献,鉴定筛选出今后可以作为改良长三角地区优质粳稻稻瘟病抗性的主要基因,以便利用分子标记辅助选择技术,进一步有针对性地进行抗病基因聚合,提高育成材料对稻瘟病的持久抗性,并为同类研究提供参考依据。
1 材料与方法
1.1 试验材料
以52份长三角地区育种科研单位选育的粳稻种质作为试验材料,均为本研究前期选育优质抗稻瘟病粳稻品种的骨干亲本(表1),对照品种为苏御糯。于2019年夏季种植于上海市农业科学院庄行综合试验站,常规水肥管理。

表1 供试水稻种质Table 1 Rice germplasms used in this experiment
用于稻瘟病抗性鉴定的供试菌株由江苏省农业科学院植物保护研究所提供,为2019年分离得到的江苏省及周边地区稻瘟病菌优势代表菌株2019-746、2019-587、2019-924、2019-16-3、2019-522和2019-863的孢子混合液。
1.2 穗颈瘟人工鉴定和发病调查
在水稻孕穗初期向幼穗穗苞内注射接种上述稻瘟病菌菌株孢子混合液,每株选1个穗苞注射1 mL,每个种质注射10株。在水稻成熟期按国家标准GB/T 15790-2009稻瘟病测报调查规范[25]调查水稻穗颈瘟的发病情况,分级记载。
1.3 水稻基因组DNA提取
在水稻分蘖盛期采集幼嫩叶片,经液氮研磨,利用CTAB法提取水稻全基因组DNA[26]。
1.4 抗稻瘟病基因的检测
本研究检测Pi9、Pib、Pi5、Pita、Pi2、Pikh、Pikm和Pigm共8个抗稻瘟病基因的分子标记特异性引物均由生工生物工程(上海)股份有限公司合成,具体分子标记的引物序列及扩增多态性差异见表2。反应体系20 μL:DNA模版2 μL,2×Hieff PCR Master Mix(with dye)10 μL,引物等体积混合液1.6 μL,ddH2O 6.4 μL。所用试剂购自上海翊圣生物科技有限公司。反应程序为:94℃预变性5 min;94℃变性30 s,55~60℃退火30 s,72℃复性30 s,共33个循环;72℃延伸10 min,10℃保存。扩增产物经含有核酸染料的2%琼脂糖凝胶电泳分离,在紫外凝胶成像系统内观察多态性差异。

表2 抗稻瘟病基因相关分子标记信息Table 2 Molecular markers related to rice blast resistance genes
1.5 数据统计分析
根据8个抗稻瘟病基因在52份粳稻种质中的分布情况,对检测结果赋值,检出稻瘟病抗性等位基因记为1,检出感病等位基因记为0。利用软件NTsys 2.1计算52个种质间的遗传相似系数,并用非加权类平均法(unweighted pair-group method with arithmetic mean,UPGMA)对遗传相似性系数进行聚类分析,得到聚类树形图。
2 结果与分析
2.1 8个抗稻瘟病基因在52份长三角地区粳稻种质中的分布
本研究检测了52份长三角地区粳稻种质中Pi9、Pib、Pi5、Pita、Pi2、Pikh、Pikm和Pigm共8个抗稻瘟病基因的分布情况(部分检测结果见图1,具体结果见表3)。

注:基因Pi9检测图中,泳道从左至右依次为分子量标记、淮稻5号、丙11-107、扬粳50226、秀水121、沪软2123、秀水114、嘉07-34、嘉33、WDR129、宁9103、WDR35;等位基因Pib和pib检测图中,泳道从左至右依次为分子量标记、WDR48、沪香12-268、WDR56、空白泳道、光明糯1号、沪11-12、沪12-285、沪12-383、沪粳144、镇稻99、沪香12-258;基因Pi5检测图中,泳道从左至右依次为分子量标记、秀水121、WDR35、沪软2123、WDR48、WDR56、武3338、光明糯1号、沪11-12、沪12-285、沪12-383、沪粳144;基因Pi2检测图中,泳道从左至右依次为分子量标记、WDR56、嘉花1号、沪软12-386、沪香12-273、沪香粳106、沪香软386、宁9103、苏香粳100、苏12-838、秀水110、秀水123;等位基因Pita和pita检测图中,泳道从左至右依次为分子量标记、苏12-838、武香粳19、武运粳21、沪香12-268、秀水110、秀水123、秀水134、秀水519、沪香12-211、南粳5055、秀水994;基因Pikh检测图中,泳道从左至右依次为分子量标记、ZHG-1、南粳9108、沪12-380、银香38、镇稻99、南粳46、沪香12-258、苏御糯、沪10-139、沪香12-540;基因Pikm检测图中,泳道从左至右依次为分子量标记、沪软12-386、沪香 12-273、沪香粳106、沪香软386、嘉花1号、武香粳19、宁9103、空白泳道、苏香粳100、苏12-838、武运粳21、秀水123Note: In the picture of gene Pi9 genotype detection the lines from lefTTo right is DNA Marker (DL2000), Huaidao5, Bing11-107, Yanggeng50226, Xiushui121, Huruan2123, Xiushui114, Jia07-34, Jia33, WDR129, Ning9103 and WDR35. In the picture of gene Pib and pib genotype detection the lines from lefTTo right is DNA Marker (DL2000), WDR48, Huxiang12-268, WDR56, blank, Guangmingnuo1, Hu11-12, Hu12-258, Hu12-383, Hugeng144, Zhendao99 and Huxiang12-258. In the picture of gene Pi5 genotype detection the lines from lefTTo right is DNA Marker (DL2000), Xiushui121, WDR35, Huruan2123, WDR48, WDR56, Wu3338, Guangmingnuo1, Hu11-12, Hu12-258, Hu12-383 and Hugeng144. In the picture of gene Pi2 genotype detection the lines from lefTTo right is DNA Marker (DL2000), WDR56, Jiahua1, Huruan12-386, Huxiang12-273, Huxianggeng106, Huxiangruan386, Ning9103, Suxianggeng100, Su12-838, Xiushui110 and Xiushui123. In the picture of gene Pita and pita genotype detection the lines from lefTTo right is DNA Marker (DL2000), Su12-838, Wuxianggeng19, Wuyungeng21, Huxiang12-268, Xiushui110, Xiushui123, Xiushui134, Xiushui519, Huxiang12-211, Nangeng5055 and Xiushui994. In the picture of gene Pikh genotype detection the lines from lefTTo right is DNA Marker (DL2000), ZHG-1, Nangeng9108, Hu12-380, Yinxiang38, Zhendao99, Nangeng46, Huxiang12-258, Suyunuo, Hu10-139 and Huxiang12-540. In the picture of gene Pikm genotype detection the lines from lefTTo right is DNA Marker (DL2000), Huruan12-386, Huxiang12-273, Huxianggeng106, Huxiangruan386, Jiahua1, Wuxianggeng19, Ning9103, blank, Suxianggeng100, Su12-838, Wuyungeng21 and Xiushui123.图1 部分种质中7个抗稻瘟病基因检测Fig.1 Detection of 7 blast resistance genes in some rice germplasms
除基因Pigm以外,其他抗病基因在52份粳稻种质中均有不同分布频率。其中分布频率最高的是Pib,达到78.85%,其次是Pita,达到50.00%;其余基因分布频率依次为38.46%(Pikh),34.62%(Pi9),23.08%(Pikm),21.15%(Pi2),5.77%(Pi5)(图2)。除对照品种苏御糯外,其余种质均有抗稻瘟病基因分布。同时携带2个抗病基因的最多,占供试种质的38.46%(图3),有12个种质同时携带抗病基因达4个,分别是秀水121、秀水114、嘉07-34、丙11-107、嘉33、2123软、武香粳19、武运粳21、秀水123、秀水134、秀水519和常农粳5号,其中8个为浙江省嘉兴市农业科学研究院选育的品种。
2.2 聚类分析
为了进一步明确供试水稻种质稻瘟病抗性的遗传差异,利用8个抗稻瘟病基因的功能标记对52份水稻种质进行聚类分析。结果表明,检测水稻种质的遗传相似系数变化范围在0~1之间,在相似系数大于0.5的水平上,将52份水稻种质分为6类。第1类包括6个;第2类包括3个;第3类有1个;第4类包括29个;第5类包括12个;第6类有1个(图4)。

表3 52份粳稻种质中8个稻瘟病抗性基因分子检测结果Table 3 Molecular detection of 8 blast resistance genes in 52 japonica rice germplasms
2.3 52份长三角地区粳稻种质稻瘟病抗性接种鉴定
本研究利用2019-746、2019-587、2019-924、2019-16-3、2019-522和2019-863的菌株孢子混合液对52份供试粳稻种质进行田间接种鉴定,依照国家标准《GB/T 15790-2009稻瘟病测报调查规范》中穗颈瘟抗性分级指标[25]对各种质进行抗性评价。结果表明,52份粳稻种质中大部分表现抗病,达到61.5%;中抗材料占比9.6%;中感材料占比3.8%,感病及高感病材料占比25.0%(表4)。
2.4 长三角地区粳稻主要抗稻瘟病基因的分析
分析发现(表5),抗病种质中携带基因Pib的最多,达55.8%,其次为携带基因Pita的种质,占比46.2%,其余依次为Pikm、Pi2、Pikh、Pi9和Pi5。其中携带基因Pikm的种质田间全部表现抗病,未发现感病种质;携带Pita、Pi5和Pi2的种质分别只发现1株感病,其余种质均表现抗或中抗稻瘟病。虽然抗病种质中携带基因Pib的最多,但感病种质比例达15.4%;携带Pi9和Pikh的种质感病比例分别有17.2%和15.3%。因此,可以将Pita、Pi5、Pi2和Pikm作为首选抗稻瘟病基因,通过高效分子聚合育种技术选育出适合长三角地区种植推广的优质粳稻新品种。携带Pib、Pi9和Pikh的水稻种质感病比例相对较高,因此这3个抗稻瘟病基因不适宜应用于长三角地区优质抗稻瘟病新品种的选育。

表4 52份水稻种质稻瘟病抗性评价Table 4 Evaluation of blast resistance of 52 tested rice germplasms

图2 8个抗稻瘟病基因在供试种质的 分布情况Fig.2 The distribution of 8 blast resistance genes in the tested japonica rice germplasms

图3 携带不同数目抗稻瘟病基因的种质在 52份供试种质中的占比Fig.3 The proportion of germplasms with different number of blast resistance genes in 52 tested germplasms

注:材料编号与表1一致。Note: The numbers are consistent with Table 1.图4 52份供试水稻种质的遗传差异聚类分析Fig.4 ClusterinGAnalysis of genetic differences in 52 tested germplasms
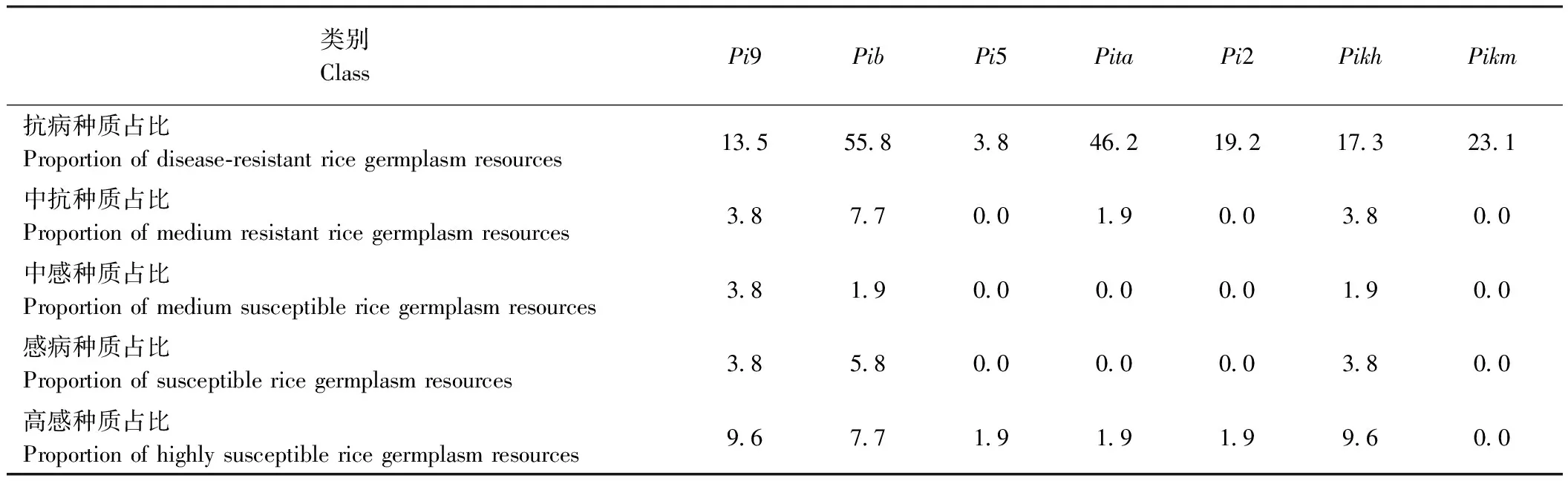
表5 抗稻瘟病基因在供试水稻种质中的抗性分布Table 5 Resistance distribution of blast resistance genes in tested rice germplasms /%
3 讨论
生产实践表明,通过抗稻瘟病基因分子聚合育种对防治稻瘟病效果显著,但由于稻瘟病菌生理小种在我国不同稻区的分布具有多样性,即使在同一稻区,不同年度间生理小种也存在差异;另一方面,稻瘟病菌的变异与分化速度快,新的生理小种层出不穷;随着新的生理小种的产生,只携带1个抗病基因的品种将会逐渐丧失抗病性,因此稻瘟病的危害持续发生。准确掌握本稻区稻瘟病菌群体组成及生理小种变化的动态趋势,有利于开展针对性的抗稻瘟病育种工作。研究发现,生理小种ZG1是江苏省的绝对优势小种,其次为ZB,并且年度间较为稳定[34-35]。近两年,江苏省农业科学院植保所在全省采集的样品中发现,ACE1、PWL1和Avr-Pita等共10个稻瘟病菌无毒基因在江苏省均有分布,频率最高的为Avr-Pib,连续5年扩增频率较高且稳定遗传的有Avr-Pib、Avr-Pik、Avr-Pizt[36]。 本研究利用该研究所2019年分离的6个代表性优势病菌小种对52份水稻种质资源进行穗颈瘟接种鉴定,结合分子标记检测结果发现,基因Pita对这6个小种表现的抗性最好,其次为Pikm、Pi2和Pi5。说明在这6个代表性小种流行的长三角地区,Pita、Pikm、Pi2和Pi5这4个抗稻瘟病基因发挥着主要作用,尤其Pita基因在供试种质资源中占50%,携带该基因的抗病种质资源占比达46.2%,充分说明了该基因对这6个小种的抗病优势。
Pigm是一个广谱且持久的稻瘟病抗性基因,来自籼稻品种谷梅4号,该基因含有2个典型的R基因,其中PigmR可导致水稻千粒重降低,产量下降,PigmS可提高水稻结实率,从而抵消PigmR对水稻产量的影响[37]。目前Pigm相关功能标记的开发及育种应用工作已逐步开展,陈涛等[38]开发了可以检测到该基因的特异性分子标记T-Pigm和K-pigm,并应用于育种实践;曾生元等[39]开发了该基因的特异分子标记Pigm-4, 并应用于粳稻穗颈瘟抗性育种;赖怡帆等[40]利用分子标记选择Pigm基因改良湘晚籼13号的稻瘟病抗性。Pigm基因来源于籼稻品种,长三角地区粳稻材料普遍不携带此基因,因此在本研究的供试种质中尚未鉴定出携带Pigm基因的材料,但是该基因具有广谱持久抗稻瘟病的特性,因此在今后长三角优质抗稻瘟病育种研究中,也需重点利用该基因。
利用8个抗稻瘟病基因功能标记对52份水稻种质进行聚类分析的结果表明,第4类中第Ⅰ小类的12份种质均含有3~4个抗病基因,接种抗性鉴定均达到R级;该类中8个种质均为浙江省嘉兴市农业科学研究院在长期通过稻瘟病菌高压筛选策略下培育成功的粳稻品种或衍生品系,试验结果与该类品种在生产上表现高抗稻瘟病的实际相吻合。由此可见,在今后抗稻瘟病育种的杂交亲本选择配置的过程中,可根据本研究中抗稻瘟病基因鉴定和聚类分析的结果,并按照抗稻瘟病基因互补的原则进行;此外还应注重开展Pita、Pi5、Pi2、Pikm和Pigm这5个基因中的双基因或三基因间的聚合育种研究,使育成材料具有广谱持久的稻瘟病抗性。
水稻与稻瘟病菌互作系统是植物-微生物互作研究的模式系统之一,植物抗病基因与无毒基因的互作关系符合经典的基因对基因学说。目前在稻瘟病病原菌中鉴定了与抗病基因对应的24个无毒基因,其中已克隆12个[41]。但由于稻瘟病病原菌的遗传背景复杂,易发生变异[42],特别是在近两年长三角地区连续出现稻瘟病流行大发生的形势下,今后需要进一步根据长三角地区稻瘟病菌优势小种的鉴定结果,利用优势小种菌株持续对长三角地区育成的各类新种质进行接种鉴定并评价其抗性,针对病原菌的变化及时调整利用与其无毒基因互作的抗病基因,可在较大程度上避免因病原菌生理小种变化导致的水稻品种原有抗性的丧失。
4 结论
培育和推广种植抗病品种是控制水稻稻瘟病危害最经济、环保、有效的措施。本研究结果表明,抗稻瘟病基因Pita、Pi5、Pi2、Pikm和Pigm在长三角地区抗稻瘟病粳稻的育种中具有重要应用价值。后续需要从上述5个抗病基因中选择2~3个互作效果好的抗病基因进行聚合育种,不断增加接种稻瘟病生理小种的数量,及时调整接种鉴定的优势生理小种,从而进一步考察其抗病广谱性和持久性,以期为长三角地区优质粳稻的安全生产提供有力保障。
猜你喜欢
今日农业(2022年4期)2022-06-01
上海农业科技(2022年2期)2022-05-13
农民致富之友(2020年32期)2020-12-03
农村百事通(2020年11期)2020-06-27
中学时代(2019年3期)2019-11-12
发明与创新·大科技(2019年5期)2019-07-31
艺术评论(2018年1期)2018-05-09
农村百事通(2017年8期)2017-04-26
金山(2016年12期)2017-02-17
中学生物学(2008年3期)2008-06-03
