
基于林木竞争指数诠释小蓬竹生境中的群落组成
2022-04-07 12:32蒋明泽于佳豪陈远松周亚琦
广东蚕业 2022年2期
蒋明泽 于佳豪 陈远松 李 湘 周亚琦
基于林木竞争指数诠释小蓬竹生境中的群落组成
蒋明泽于佳豪陈远松李湘周亚琦
(贵州师范大学贵州贵阳550000)
文章从物种竞争角度对小蓬竹种间竞争强度进行定量分析,研究其在不同生境下的适应性。在喀斯特地貌上选取小蓬竹常见的3种小生境样地(Q1石槽为主,Q2石沟为主,Q3石隙为主),对每种样地进行每木检尺。基于Hegyi林木竞争指数,分别对小蓬竹进行胸径与距离加权。结果表明:胸径方面,小蓬竹(对象木)竞争指数随着胸径的增大而增大;距离方面,二者并非距离越近竞争越激烈,而是存在最佳共存距离1.112 m(常见树种与小蓬竹的平均距离)。试验结果显示,基于胸径测量的竞争指数成正比,基于距离测量的竞争指数存在距离极值。
小蓬竹;物种竞争;林木竞争指数;Voronoi图;生态位
小蓬竹()系竹亚科镰序竹属,其地下茎呈合轴型,秆柄种在喀斯特生境和小生境中有良好生态功能,其株上部呈廉蓄状,株形美观,具有很高的园林观赏价值。小蓬竹在《中国物种红色名录》中被列为极危种。经过文献阅读,笔者发现造成此情况的原因有三个:第一,从繁殖特性来看,小蓬竹属于无性繁殖,以克隆为主的繁殖方式使得其分布区很难扩大;第二,小蓬竹虽是喀斯特生境优质种群,但石沟、石槽等生境地形破碎,种间竞争异常等加剧了小蓬竹的适生难度;第三,人为的大面积砍伐,加上少有人工扩繁,使得小蓬竹在短时间内数量骤减。随着贵州省交通道路的完善,贵州旅游业呈现井喷式发展,小蓬竹作为喀斯特地貌适生性竹种[1],在当前贵州大生态的背景下,更体现其生态位的重要性。虽众多学者已对小蓬竹无性系构件繁殖、土壤理化性质、光合特性、种群适生特性等方面展开分析,但少有学者采用Hegyi简单林木竞争指数以及加权Voronoi图[2]来分析林木之间的生长关系与竞争效应。为此,本文通过分析小蓬竹群落上层主要种群空间位置特征,从物种竞争角度,利用林木竞争指数并加以胸径和距离的加权,解析小蓬竹在喀斯特小生境中群落物种竞争的生态适应性。曹靖夫等(2007)的研究表明,在竹类邻体干扰指数中,因竹笋出土后胸径基本成型,故不适合将胸径作为唯一判断竞争指数的指标[3]。但竹类植物的盖度依然受到胸径和高度的影响,竹类测量林木竞争指数已有科学测量方法[4],从而在研究单木竞争指数时可以引入胸径作为参考依据且信度较高。
1 材料与方法
1.1 研究区概况
1.1.1 地理概况
研究地点为贵州省南部边缘罗甸县董架乡。境内以山地为主,其中包括岩溶、丘陵、盆地及石炭岩喀斯特等地貌。董架乡位于东经106°53′~106°54′,北纬25°37′,平均海拔997 m。境内土壤质地偏中壤土,坡下部的土壤水分大于坡上部[5]。区域内属典型的亚热带温暖湿润季风气候,基本无严寒,冬季平均降水量17.3 mm,夏季平均降水量156.3 mm,年平均降水量1 100 mm~1 400 mm。
1.1.2 样地设置
在喀斯特地貌上选取小蓬竹常见的3种小生境样地:Q1石槽为主,Q2石沟为主,Q3石隙为主。每种生境选取3个10 m×10 m的样方,并对每个样方进行每木检尺,调查其株数、胸径、株高、冠幅,并采用网格法测量其空间坐标以及海拔、坡向、坡度等立木因子。样地内基岩裸露,生境破碎,是典型的喀斯特小生境特征,在小生境下研究植被竞争关系本身有较大难度,但侧面更能反映出种群适生性以及物种竞争指数,有更强信度。
1.1.3 植被特征
生境中植被以小蓬竹为主,其余植被类型主要包括壳斗科(Fagaceae)、樟科(Lauraceae)、山茶科(Theaceae),主要植物有黄连木()、朴树()、榆树()、蚊母树()、椤木石楠()、山核桃()、齿叶铁仔()、檵木()、绒毛石楠()等
2 研究内容
2.1 加权Hegyi的单木竞争指数分析
采用加权Hegyi单木竞争指数能更好地反映生境内的种间关系。对象木胸径大小一般与竞争指数成正比,在距离方面,一般认为竞争木和对象木距离越近,竞争指数越高。本研究以小蓬竹为对象木,其周边的树木为竞争木。通过加权胸径后,对小蓬竹周围竞争木进行加权CI值(竞争指数)分析,得到不同竞争木与对象木小蓬竹的单木竞争指数,以此说明物种竞争的空间位置关系。
2.2 样地信息
所选3种小生境样地的基本状况如表1。
表1生境基本信息表
生境小蓬竹立竹密度/株/hm2小蓬竹盖度/%海拔/m坡度/°坡位坡向 Q1(石槽为主)1 230~1 39043.8%1 001.732上坡位东北25° Q2(石沟为主)1 830~2 59054.3%79539中坡位西南28° Q3(石隙为主)1 280~2 31052.3%88025下坡位西北30°
2.3 确定竞争范围
2.3.1 竞争关系确定
林木竞争指数反映了林木所承受的压力,能够对物种之间的竞争关系量化表达。本文选用加权Hegyi林木竞争指数,结合Voronoi图使林木竞争因子更具说服力。采用固定半径确定竞争木的范围的方法,很容易把非直接竞争者划入竞争木范围,也可能会把不同方位存在直接竞争关系的物种排除在竞争结构单位外,同时还有可能将竞争木划在样地范围之外。而Voronoi图是根据坐标的离散分布来确定空间点的效应范围,从而反映空间各点坐标的邻近关系,有效避免了固定半径缺陷,详情见图1。

图1 3种生境的Voronoi图
2.3.2 基于Voronoi的胸径加权图
Voronoi图所确定的竞争单元应根据林木的相对位置结合林木现阶段的胸径、冠幅等进行改进,借助ArcGIS软件生成加权Voronoi图。本文结合郝月兰等(2012)的胸径加权Voronoi图[6]以及惠刚盈(2013)提出的最佳空间结构单元[将与对象木相邻且最近的4株作为竞争木(=4)][7]来确定胸径加权Voronoi图,提高了3种生境内林木竞争因子的准确性,见图2。
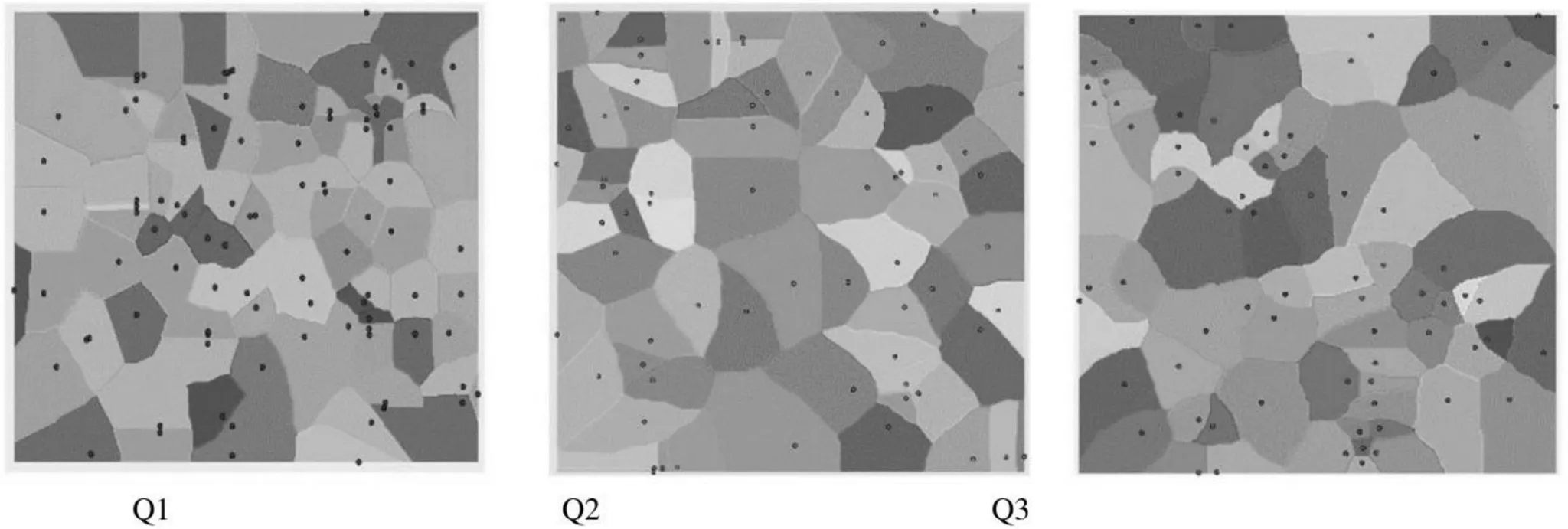
图2 3种生境的胸径加权Voronoi图
距离VN图与加权VN图只是在竞争边界发生偏移,但从图2~图4中不难看出3种生境下地形对植被的切割作用。其中Q1石槽植被聚集现象明显,竞争激烈;Q2石沟植被依然集中,竞争激烈;而Q3石隙植被分布较均匀,小蓬竹与竞争木处于理想竞争状态。加权Voronoi图反映出胸径越大,植株所覆盖的范围就越大,就会与周围更多树种产生竞争,图中深色部分为植株胸径与竞争木胸径产生竞争强烈的区域,该区域内可能会出现植被上部覆盖遮挡现象。
2.3.3 边缘消除
为消除边缘效应(竞争木与实测地之外的树种也被计算在内),需要进行边缘矫正。本文采用距离缓冲区法,把样地每条边向外3 m水平距离的范围作为缓冲区[8]。边缘效应为10 m×10 m外面外加3 m×3 m的正方形,每个样方之间都有3 m的间隔,以使数据更准确,如图3。

图3 边缘消除样图
2.3.4 林木竞争指数
采用改进的Hegyi林木竞争模型[9]:
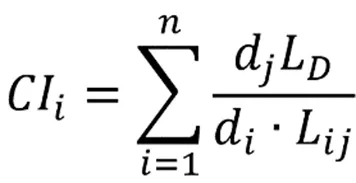
式中,CI为林木的竞争指数,L为对象木与竞争木之间的距离,L为林分生境空间单元中4棵竞争木与对象木的平均距离,d为对象木的胸径,d为竞争木的胸径,为竞争木的株数。CI值越大,表示对象木受到竞争木的压力越大,竞争木基于Voronoi图确定。最后,通过SPSS软件绘制一次线性回归图、幂函数图和指数函数。
3 结果与分析
3.1 小蓬竹胸径与竞争指数关系
不同生境中小蓬竹平均胸径与竞争指数的关系如图4~图6,函数模型如表2~表4。

图4 Q1中小蓬竹平均胸径与竞争指数的关系
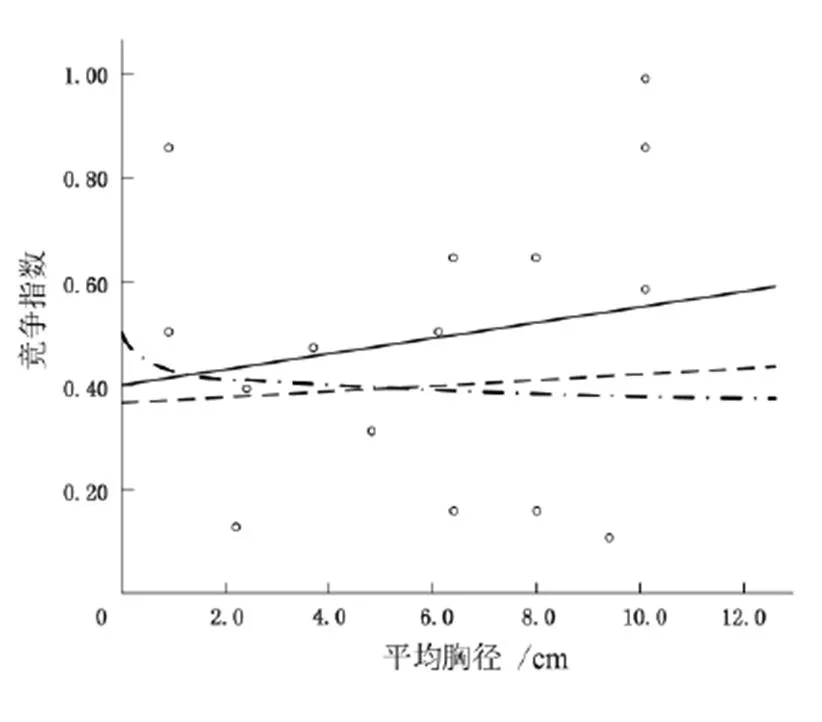
图5 Q2中小蓬竹平均胸径与竞争指数的关系
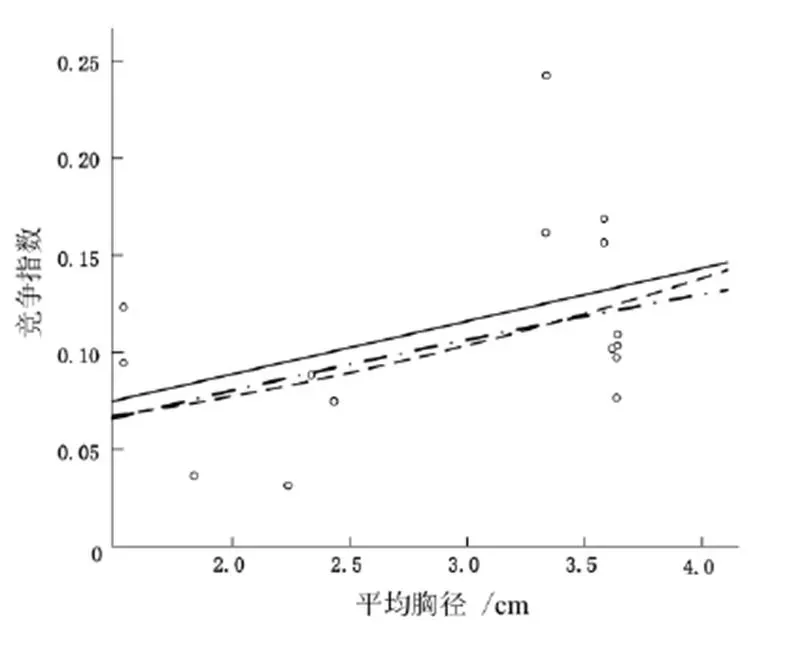
图6 Q3中小蓬竹平均胸径与竞争指数的关系
表2Q1函数模型
方程式R平方显著性 线性0.1460.178 次方0.2710.056 指数模式0.2700.053
表3Q2函数模型
方程式R平方显著性 线性0.0290.542 次方0.0030.848 指数模式0.2220.076
表4Q3函数模型
方程式R平方显著性 线性0.2290.542 次方0.1920.118 指数形式0.1780.076
图4~图6中每种生境共有3种方程,同时函数图像均呈正态分布,由此表明竞争木胸径越大,与对象木的竞争就越激烈。Q1和Q3中,竞争木(包括乔木、灌木层)平均胸径基本分布在4.5 cm以内,函数图像较好地以正态分布的形式呈现(Q1中线性方程为=0.014+0.400,2=0.146,置信水平>95%;Q2中的线性方程为=0.027+0.015 6,2=0.029,置信水平>95%;Q3中的线性方程为=0.024 7+0.043,2=0.229,置信水平>95%),这反映出3个生境内对象木胸径与竞争指数之间的正态关系。Q1与Q3中,乔灌层混交现象不明显且各树种分布分散。Q2的植被复杂系数增加,回归方程为=0.027+0.015 6,2=0.029,置信水平>95%,但幂函数为=0.4240.047,对象木的平均胸径与竞争指数成反比,但2为0.003,说明Q2中反比现象普遍存在,这并不合乎常理。Q2中当对象木胸径增大时,竞争木与对象木的竞争效应反而减小,很有可能是因为Q2中破碎的石沟、石槽等阻断了植被的扩繁范围,使该生境内植被复杂系数增加,导致该样地的植被未能像其他样地一样反映出正态关系,也有可能是实验数据的误差导致,或者样地太小,反映不出真实的竞争现状。以上结果表明:整体上来说,竞争木的胸径与竞争指数成正比,但因以石沟为主的Q2地形破碎,再加上样地面积相对较小,以及野外考察数据采集存在偏差,影响了实验结果。
3.2 种间竞争中距离与竞争指数的关系
表5为生境中的主要竞争木(乔木与灌木各3种)的胸径分布情况和平均竞争指数,竞争指数最高的是乔木层樟叶槭(平均CI=0.684),说明樟叶槭与小蓬竹存在较大的竞争效应,对光照与热量的需求大体一致。本文更加关注小蓬竹与竞争木之间距离与竞争指数的关系,从数据中挑选出具有代表性的几组数据并简要说明。3种生境中,樟叶槭与小蓬竹共存的有效半径为1.672 m,当胸径为10.7 cm时,樟叶槭的CI值达到最大(0.109),所以其与小蓬竹的最佳共存范围在1.93 m之外。朴树与小蓬竹共存的有效半径为1.291 m,当胸径为9.6 cm时,CI值达到最大(0.268),其与小蓬竹最佳共存范围在1.4 m之外。椤木石楠与小蓬竹共存的有效半径为0.905 m,当胸径为2.89 cm时,CI值达到最大(0.126),二者最佳共存范围在2.94 m之外。红花檵木与小蓬竹共存的有效半径为0.832 m,当胸径为3.74 cm时,CI值达到最大(0.98),同时与小蓬竹最佳共存范围在0.3 m之外。绒毛石楠与小蓬竹共存的有效半径0.564 m,当胸径为7.03 cm时,CI值达到最大(1.419 7),与小蓬竹产生激烈竞争,之后竞争指数下降,二者最佳共存范围在0.1 m之外。因此,主要竞争种群平均最佳共存距离为1.112 m。各生境中红花檵木种群数量多,占生境植物量(不包括草本)的22.9%,但从表5中可知红花檵木的CI值为0.087,说明红花檵木有可能是小蓬竹的优质伴生种,其在对光照、降水以及其他生境因子的需求方面可能与小蓬竹不相同或者相似度很低。综上可知,对象木与竞争木平均距离越近,二者竞争指数越大。也基本说明,竞争木胸径越大,与对象木竞争越激烈。
表5主要竞争木竞争指数及有效半径
主要竞争木拉丁名不同生境林分单木竞争指数(CI) Q1平均竞争指数Q2平均竞争指数Q3平均竞指数平均合计有效半径/m 朴树(CeltissinensisPers)0.5780.7900.0650.4771.291 香叶树(LinderacommunisHemsl.)0.3100.0570.0850.1501.027 樟叶槭(Acercinnamomifolium)0.8720.9830.1990.6841.672 椤木石楠(Photiniadavidsoniae)0.4720.8410.1140.4760.905 红花檵木(Loropetalumchinense)0.1200.0530.1990.0870.832 绒毛石楠(Photiniaschneideriana)0.1350.1230.0380.0980.564
4 结论与讨论
本文主要讨论小蓬竹与竞争木的胸径和距离关系,分析了竞争指数(CI)较高的六大树种与小蓬竹在3种生境中的竞争关系。结果显示,小蓬竹和竞争木胸径与CI值基本呈正相关关系(信度均大于95%),说明胸径越大,竞争越激烈。当进行距离加权后,发现二者产生了最优共存范围。也就是说,二者并非距离越近竞争越激烈,而是在CI值最大点所对应的距离处竞争相对激烈,且为良性竞争,二者位置拉近可能产生恶性竞争。值得注意的是,Q2函数图中,竞争木胸径与CI值成反比。推测可能原因是物种间关系复杂,交叉多,再加上小生境地形破碎且存在高差,单纯用二维距离作为竞争因子已难以判断二者的竞争关系。因此,根据小蓬竹的生长特点,需要加上冠幅、胸径、距离、高程,选取样地的范围要足够大,才能更好地反映出物种间的竞争关系。
本次实验亦存在缺点,如单纯将植株胸径与距离做对比而得出结论并不十分准确。基于ArcGIS软件来模拟加权胸径距离图有前提条件,即对象木周围的竞争木数量为4,这必然导致样地面积的扩大或者缩小(如样地1在VN图中面积为10 m×11 m)。同时计算机模拟加权胸径所得到的竞争关系是机械化的,忽视了众多生境因子,而且单纯将生境内相关性强的物种与对象木进行比较而抛弃其他细节,依然会导致实验数据偏差。冠幅更是重要因素,树冠之间会相互遮挡,尽管样地内仅朴树、香樟等乔木可能会遮挡小蓬竹,即遮挡因子是可控的,但盖度会直接影响植株被遮挡部位的光合作用以及其他更多不可控因子[10]。物种竞争与环境之间的关系受到多重因素的影响。为了更好地反映小蓬竹生境的物种竞争情况,除了胸径、距离,未来的研究中还可以加入冠幅与距离的加权,或者胸径、冠幅、距离加权,从而更好地反映出物种间的竞争关系[11-12]。
此外,本研究只考虑了植被地上芽部分,虽然胸径指数一部分可以说明地下芽与地上植被外貌特征的关系,但仍不完全,探究小蓬竹根系和土壤的理化性质与地上生境因素相结合也是未来研究小蓬竹群落物种间竞争关系的新方向,或许只有将土壤理化性质与实际立株距离结合,小蓬竹种间竞争关系研究才能打开新天窗。
[1]刘济明.贵州喀斯特地区小蓬竹生态特性研究[D].北京:北京林业大学,2010.
[2]陈志斌.台湾相思次生林胸径与竞争指数关系[J].绿色科技,2018(9):121-122.
[3]曹靖夫,蒙朝阳,周超,等.小蓬竹群落邻体干扰指数的研究[J].安徽农业科学,2007,35(25):7754-7755.
[4]曹永慧,萧江华,陈双林,等.竹阔混交林中阔叶树对毛竹生长的影响及竞争关系[J].浙江林学院学报,2006(1):35-40.
[5]汤孟平,陈永刚,施拥军,等.基于Voronoi图的群落优势树种种内种间竞争[J].生态学报,2007(11):4707-4716.
[6]郝月兰,张会儒,唐守正.基于空间结构优化的采伐木确定方法研究[J].西北林学院学报,2012,27(5):163-168.
[7]惠刚盈.基于相邻木关系的林分空间结构参数应用研究[J].北京林业大学学报,2013,35(4):1-9.
[8]曹永慧,萧江华,陈双林,等.竹阔混交林中阔叶树对毛竹生长的影响及竞争关系[J].浙江林学院学报,2006(1):35-40.
[9]李际平,房晓娜,封尧,等.基于加权Voronoi图的林木竞争指数[J].北京林业大学学报,2015,37(3):61-68.
[10]杨建宇,刘晓娟,杨晓杰.森林冠层结构与光环境及其对林下植被生长和格局的影响[J].高师理科学刊,2015,35(7):57-62.
[11]师静,李任波,卢萍.昆明地区针叶混交林林木竞争指标研究[J].内蒙古林业调查设计,2010,33(1):26-28,89.
[12]封磊,洪伟,吴承祯,等.杉木-观光木人工混交林林木竞争效应研究[J].热带亚热带植物学报,2008(6):505-510.
10.3969/j.issn.2095-1205.2022.02.09
Q948
A
2095-1205(2022)02-28-05
贵州师范大学大学生科研训练计划项目(S202010663022)。
蒋明泽(1998- ),男,汉族,山西大同人,本科,研究方向为地理科学等。
周亚琦(1987- ),男,汉族,河南漯河人,博士,副教授,研究方向为竹林培育。
猜你喜欢
绿色天府(2022年9期)2022-11-19
湖南林业科技(2022年4期)2022-08-29
农业与技术(2022年15期)2022-08-15
世界热带农业信息(2022年8期)2022-07-19
贵州畜牧兽医(2022年3期)2022-06-28
种子(2022年2期)2022-03-21
风景园林(2022年1期)2022-03-07
防护林科技(2021年3期)2021-09-12
中国科技纵横(2016年15期)2016-12-29
南方农业·下旬(2014年2期)2014-09-23
