
p300诱导的乙酰化修饰参与脂多糖诱导的炎症介质合成
2022-04-13 01:51曹湘玉李玉娴刘灵丽陈月富陈立军黄民江谭碧峰尹辉明
南方医科大学学报 2022年3期
在严重急性呼吸道综合征(SARS)和新型冠状病毒肺炎(COVID-19)2次重大呼吸道病毒感染爆发过程中,全身炎症反应综合症(SIRS)造成的多器官功能障碍综合征(MODS)是死亡的重要原因。目前临床处理SIRS时一般会应用激素,虽然在一定程度上可以降低死亡率,但带来的副作用同样不可忽视,亟待医学界探索出其他可行的方案。与炎症启动和发展关联的分子环节当中,经典以及非经典核因子(NF-kb)家族成员于目前仍占有核心地位。除此之外,丝裂原活化蛋白激酶(MAPKs)、Toll样受体(TLRs)、络氨酸激酶/信号转导和转录激活子(JAK/STAT3)、磷脂酰肌醇3-激酶(PI3K)、Hippo通路乃至细胞焦亡相关通路都被证实起到了积极作用。上述通路的相关小分子抑制剂多数在实验室条件下表现出一定的抑炎效果,但在成果转化方面仍少见报道,提示炎症发生发展的分子机制仍然未被充分挖掘。研究表明当碱基或组蛋白发生表观遗传修饰时,染色质构像将发生改变,从而影响转录因子与靶基因启动子区域的结合,使得基因表达发生变化。譬如辅助激活因子p300就是通过诱导组蛋白H3第27位赖氨酸(H3K27)发生乙酰化修饰,从而松弛染色质结构和激活相关基因表达。p300诱导的表观遗传修饰是否介入了脂多糖(LPS)诱导的炎症介质合成过程,现有研究还未见详细报道。
基于以上背景,我们采用有关实验技术路线,探究出LPS 刺激条件下RAW246.7 内p300 合成的变化规律,并且验证了p300与炎症基因启动子存在结合现象以及由此结合产生的乙酰化修饰作用在诱导炎症基因表达过程中的积极作用,提示干预p300与炎症基因启动子的结合能力或与之相关的乙酰化修饰活性,很可能成为逆转临床炎症失衡的全新策略。
1 材料和方法
1.1 细胞株及主要试剂
小鼠巨噬细胞、RPMI 1640培基由南京华奥生物技术有限公司提供。安捷伦G3 平台小鼠表达谱芯片(Agilent Sureprint G3 Mouse Gene Expression v2 8x60K Microarray)由Agilent和John Rinn 实验室共同设计,涵盖39 430条Entrez Gene RNAs以及16 251条lincRNA。兔抗小鼠RFX、Runt、Far1、Tmem176b、Ttc24、Tmem163、Csf3、Src、p65、p300、c-myb以及二抗均购自博大泰克公司。高表达p65、p300、c-myb蛋白的细胞提取物(以HEK293为载体,制备步骤详见参考文献[21])以及pEGFp-L1-p300和各干扰质粒均由武汉生工股份有限公司合成。
1.2 安捷伦G3小鼠表达谱芯片筛选与LPS刺激强度相关的转录因子
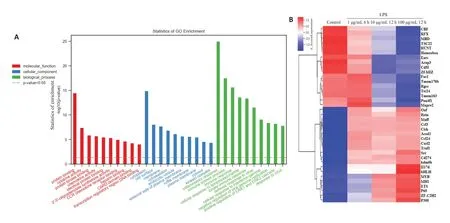
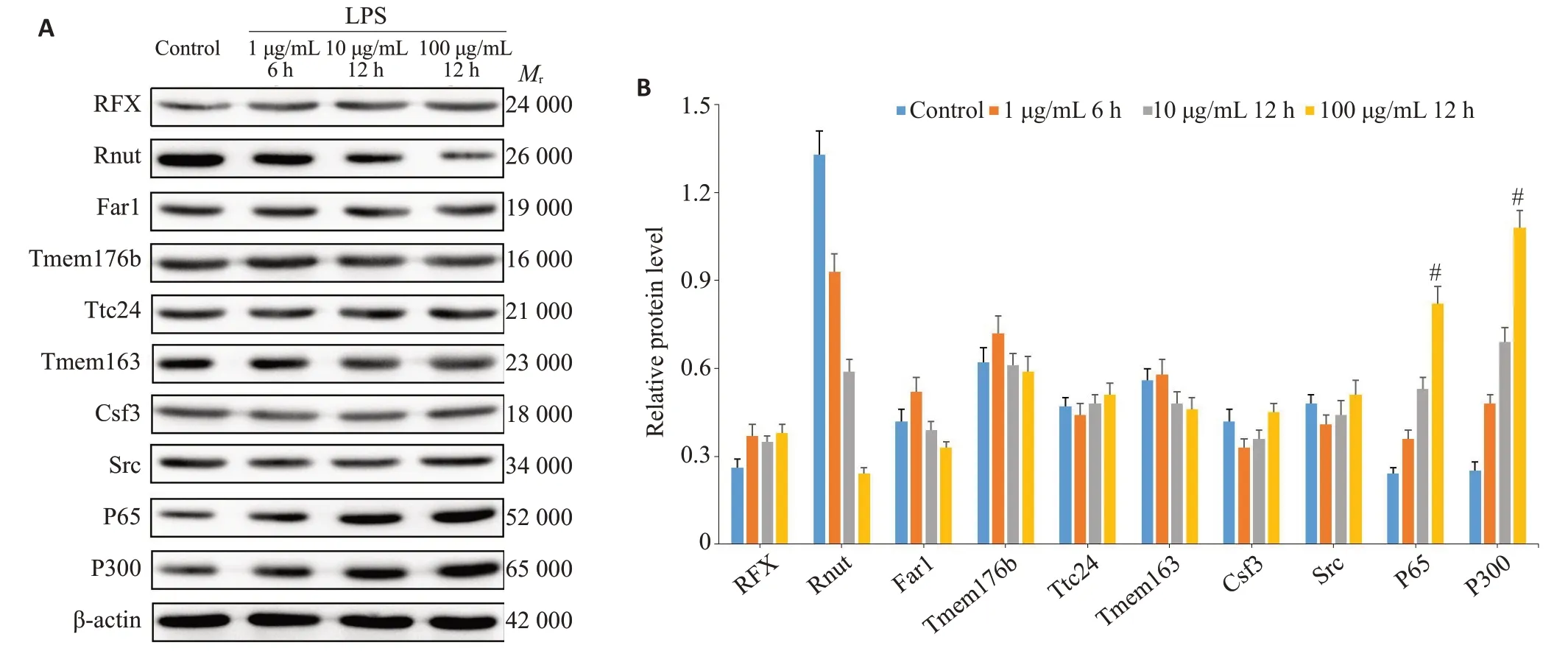
复苏后的Raw264.7细胞于RPMI 1640内培养,根据不同LPS刺激浓度以及时间分为以下4组:(1)1µg/mL LPS刺激6 h;(2)10µg/mL LPS刺激12 h;(3)100µg/mL LPS刺激12 h;(4)对照组:采用等体积培基加入。刺激结束后,MTT检测细胞活性,80%以上的对象进行微阵列芯片杂交实验,委托上海欧意生物医学科技有限公司完成。主要步骤为总RNA提取后进行纯化,后反转录成cDNA,再转录合成cRNA,样品片段化处理后进行荧光标记以及芯片杂交(65 ℃10 r/min滚动18 h)。结束后进行清洗及扫描,得到杂交图片。扫描后通过Feature Extraction 软件抽提数据,生成的原始数据经Genespring 标准化。将mRNA 表达差异倍数≥2,且<0.05的定义为差异表达基因(DEGs)并进行GO功能分析。筛选其中10个代表性基因的蛋白产物进行WB检测,以验证芯片结果的可靠性。
1.3 Raw264.7内p300与c-myb的免疫共沉淀检测
用含蛋白酶抑制剂的RIPA 裂解液提取Raw264.7蛋白,将蛋白样品分为免疫共沉淀、无关对照以及裂解液组(作为input)。于免疫共沉淀组加入兔抗小鼠多克隆c-myb抗体,无关对照组中加入兔抗小鼠IgG,混入蛋白A/G琼脂糖,4 ℃孵育过夜。清洗沉淀复合物以去除非特异结合的蛋白,将琼脂糖珠-抗原抗体复合物用2×Loading Buffer 重悬,95℃水浴5 min,100 g/L SDSpAGE 凝胶电泳,利用Western blot方法检测p300表达。
1.4 电泳迁移率实验(EMSA)确定鼠IL-6以及TNF-α基因结构上的转录因子结合位点
安捷伦G3平台小鼠表达谱芯片显示LPS刺激下RAW246.7内共有850个基因表达上调,676个基因表达下调(<0.05)。对差异表达基因进行GO功能分析,提示与LPS刺激有关的分子功能变化最为突出的是“蛋白结合”,而生物学过程改变则以“炎症反应”最为显著(图2A)。图2B为对照以及不同LPS刺激条件下芯片筛选的部分差异表达基因的热图。
作以边长为a的正方形ABCD,连接对角线BD,并以BD为边长作正方形BEFD,设其边长为b,如图所示,则有b2=2a2
1.5 Chip-qPCR法检测RAW246.7胞内相关分子与炎症基因启动子的结合
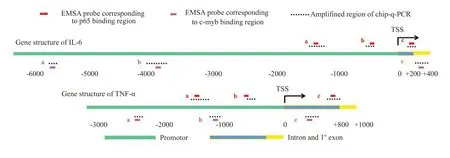
复苏成功的RAW264.7进行c-myb表达干预(质粒转染步骤详见1.6),同时设立分非干预组。搜集活力理想的细胞10个,Chip-qPCR 主要步骤见下:(1)蛋白-DNA交联:1%甲醛溶液固定,甘氨酸溶液终止反应;(2)超声断裂染色质:按100 W,10 s/次,间隔30 s的方式断裂细胞裂解液中的染色质,共5次;(3)免疫共沉淀:加入p65以及p300单克隆抗体(10 μg/管),4 ℃摇床孵育12 h,使用蛋白G-琼脂糖小珠吸附免疫沉淀复合物;(4)蛋白-DNA解交联:加入5 mol/L NaCl溶液,65 ℃水浴过夜,DNA纯化柱回收;(5)Real-time PCR分析结合启动子片段的相对含量:其中PCR引物应满足:(1)覆盖至少1个转录因子结合位点;(2)扩增区域位于EMSA实验寡聚核苷酸引物附近(分别对应图1中a、b区域),上下不超过200 bp范围。反应条件为94 ℃5 s,60 ℃30 s,共40个循环。
1.6 Western blotting法检测胞内p65、p300以及c-myb的含量
于培养板各孔准备RAW246.7 10个,按不同干预条件分为9 组:(1)pEGFp-L1-p300 转染组;(2)p65SiRNA转染组;(3)pEGFp-L1-p300+p65SiRNA联合转染组;(4)pEGFp-L1-p300+c-mybSiRNA转染组;(5)LPS 组(10 µg/mL 刺激12 h,以下同);(6)LPS+p65SiRNA联合干预组;(7)LPS+p300SiRNA联合干预组;(8)LPS+c-mybSiRNA联合干预组;(9)对照组:加入RPMI 1640,使培基总体积等于前面干预组。转染前将Lipofectamine 2000与Opti-MEM混合,静置5 min,再与经Opti-MEM稀释的过表达或干扰质粒混匀,静置20 min后缓慢加入RAW246.7,4 h后加入LPS,继续于37 ℃、5%CO条件下培养48~72 h。搜集活力理想细胞数量5×10个,根据《分子克隆实验指南》进行胞核蛋白提取、SDS-PAGE转膜以及封闭操作。分别加入兔抗小鼠p65和p300单克隆抗体(稀释度1∶1000)以及c-myb以及β-actin单克隆抗体(稀释度1∶2000),4 ℃孵育过夜;TBST洗膜5 min×3次,再加入HRP标记的二抗(稀释度1∶1000),室温孵育2 h;洗膜后ECL显影。AlphaEase FC Version 4 软件检测蛋白条带的吸光度()值,A目的蛋白/AGAPDH的比值视为表达水平。
1.7 染色质免疫沉淀-测序(Chip-seq)技术分析炎症基因相应区域H3K27乙酰化修饰以及转录因子的结合水平
各干预组选取5×10个RAW 264.7细胞(经MTT检测活力在80%以上),采用甲醛固定以及甘氨酸终止固定。室温摇晃5 min后,4 ℃离心并用预冷PBS清洗,后使用1 mL含有抑制剂的缓冲液裂解,冰上孵育10 min。冰上超声(功率设置为50 W,共计8次,每次1 min,间隔1 min)打断DNA 片段至100~500 bp 大小,4 ℃、14 000 r/min离心10 min,保留上清。均分成2管,其中1管加入2 μg IgG抗体作为Chip 样品,另1管加入2 μg兔抗小鼠p65、p300 或者乙酰化H3K27 单克隆抗体,4 ℃孵育过夜。之后4 ℃、14 000 r/min离心15 min,弃上清。elutionbuffer进行2次洗脱,振荡15min,14000r/min离心10 min后搜集上清。后利用DNA 快速PCR 纯化试剂盒进行DNA纯化,赛默飞200 超微量分光光度计检测DNA浓度。筛选出100~200 bp产物,3'端连接A碱基以及测序接头,委托上海欧意生物医学科技有限公司进行高通量测序。利用CisGenome v2.0软件将测序数据格式化为wig文件,上传IGV_2.9.4系统进行数据可视化。重点分析EMSA验证的结合位点的附近范围,对比基因组序列为mouse_mm10,错配不超过2个碱基的数据可被利用,最后得到p65、p300 以及乙酰化H3K27结合峰(peak)的图像结果。
1.8 RAW246.7分泌IL-6 以及TNF-α水平测定
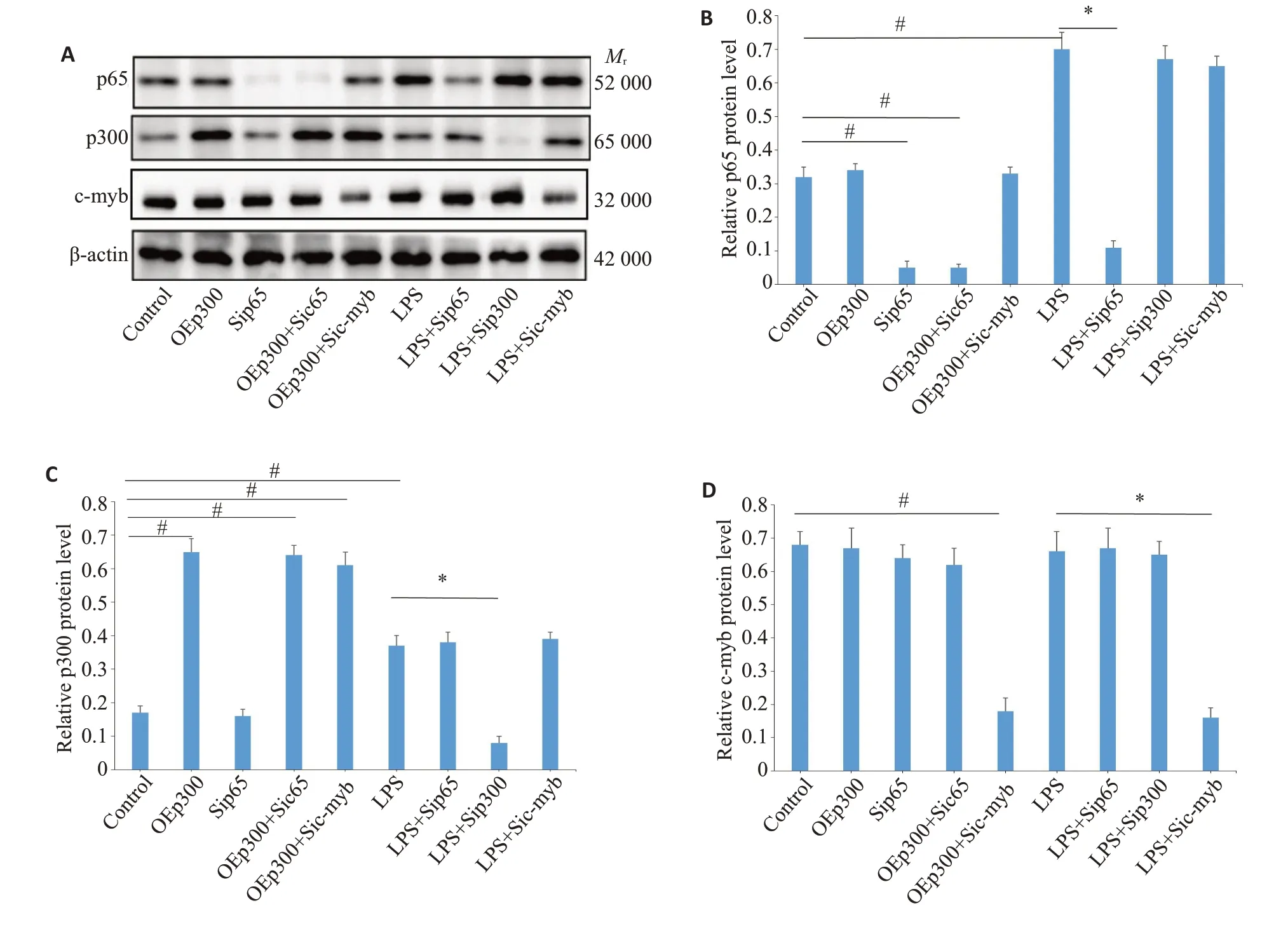
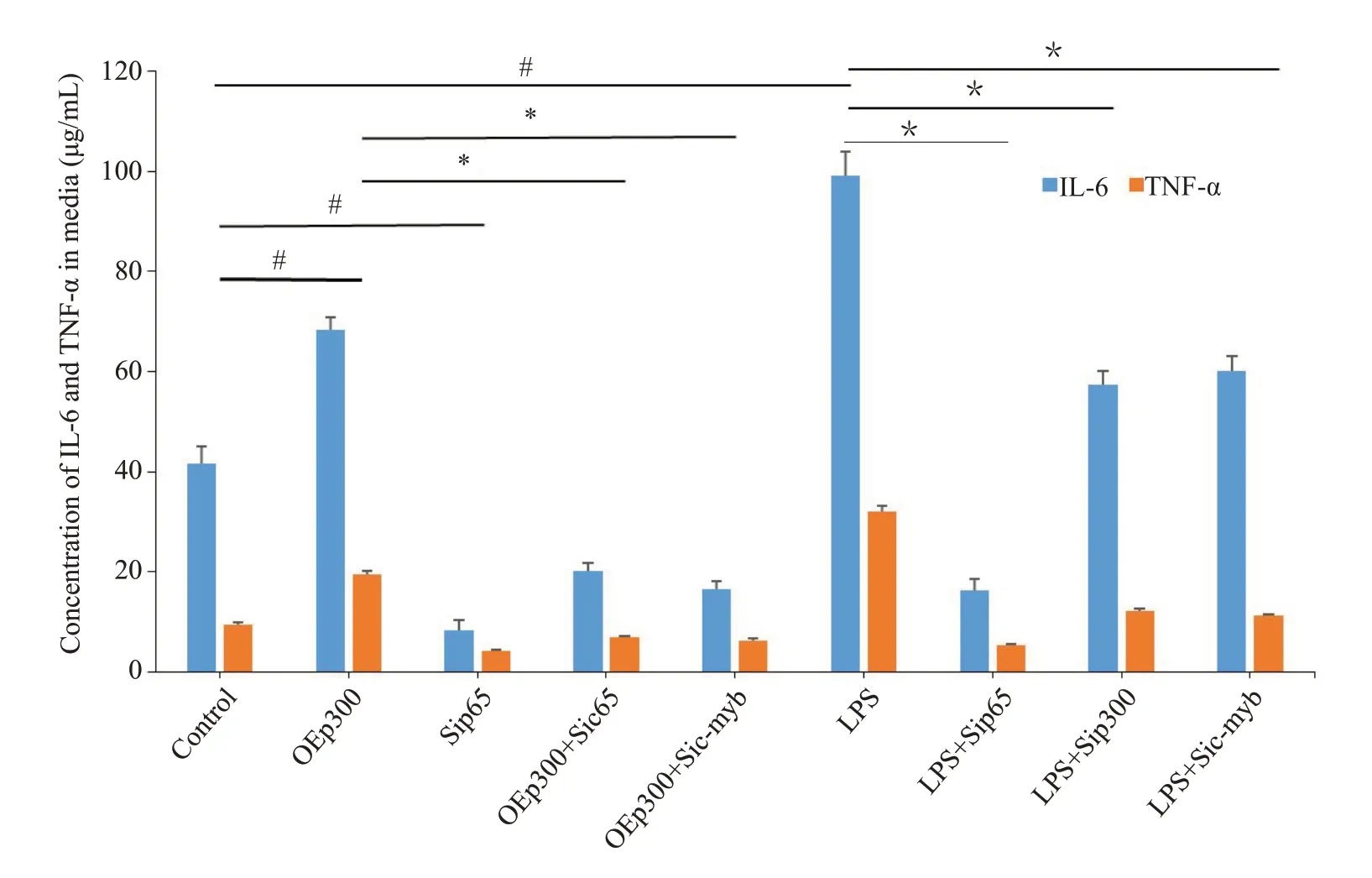
2.3.1 p65、p300以及c-myb在RAW246.7胞内的表达无相互影响 Western blot 对各干预组RAW246.7 胞内p65、p300 以及c-myb 的检测结果显示(图7A~D),pEGFp-L1-p300以及各种干扰质粒均可以上调或者遏制相应蛋白的表达(<0.05);然而p65、p300以及c-myb各自的表达均互不干涉。
1.9 统计学分析
而企业之间的合并也分多种,比如中国远洋和中国海运合并为中远海运集团,属于国企之间的合并重组,比如顺丰与夏辉合并为新夏辉,属于中外合资。
2 结果
2.1 RAW246.7胞内p300的表达与LPS刺激强度存在显著关联
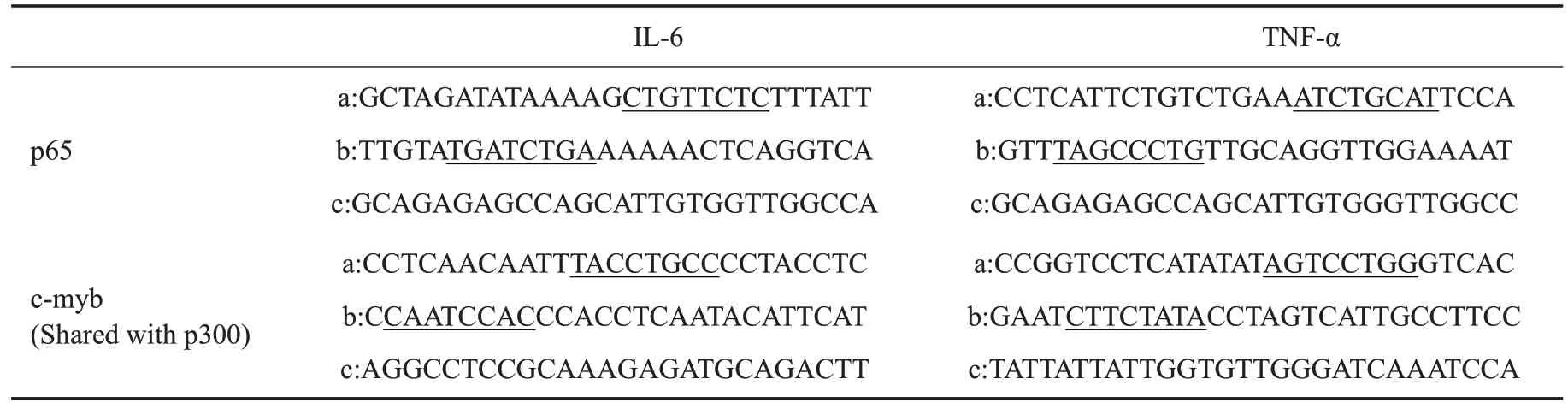
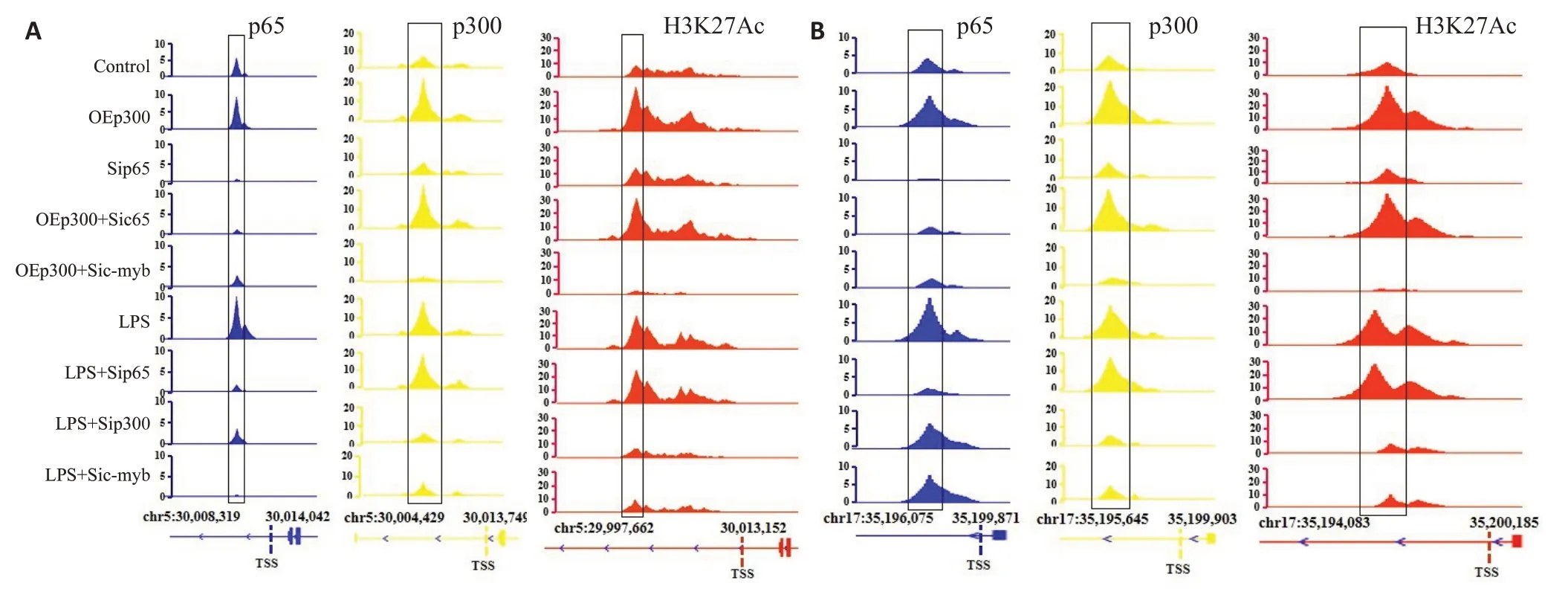
联合UCSC数据库(http://genome.ucsc.edu/)以及转录因子结合位点鉴别工具PROMO 网站(http://alggen.lsi.upc.es/cgi-bin/promo_v3/promo/promoinit.cgi?dirDB=TF_8.3)对鼠IL-6以及TNF-α的基因结构以及启动子部位的转录因子结合位点进行预测,显示存在较为密集的为核因子p65以及MYB类转录因子成员c-myb的结合位点,而未见p300结合位点(图1)。根据string生物信息学数据库以及免疫共沉淀检测结果,推测辅助激活因子p300是通过c-myb的介导才能够结合于IL-6以及TNF-α启动子,而并非直接结合。据此为p65以及c-myb各设计出2条寡聚核苷酸引物(分别对应图1中a、b区域,p300与c-myb共用相关寡聚核苷酸引物),覆盖了至少1个转录因子结合位点,同时根据转录起始位点(TSS)之后的内含子(对应图1中c区域)序列,设计1条大小相当但未覆盖转录因子结合位点的引物,另外还准备了标准阳性引物作为参照,全部序列信息见表1。配置好SDS-pAGE(50 g/L),按顺序加入寡聚核苷酸、标准阳性引物、近红外染料(IRDye-680)以及p65、p300或者c-myb纯蛋白提取物。室温下避光孵育30 min后点入凝胶点样孔中,80 V恒定电压下电泳1.5 h,待染料接近凝胶底部时停止,红外荧光成像系统扫描,选择650 nm波长读取结果。
随着社会的变化与发展,动物疫病的流行趋势也发生了一定的变化,这使得许多疫病以及病毒如果采用常规的方法,则难以检查出来,等到疫病爆发后,再采取相应的防治方法,那么就会对社会造成巨大的危害。而采用动物疫病监测,就能够对一些难以发现的疫病进行提前监测,使相关部门能够进行提前预防,以便对疫病的危害进行控制。
2.2 p300可在c-myb介导下结合于炎症基因启动子
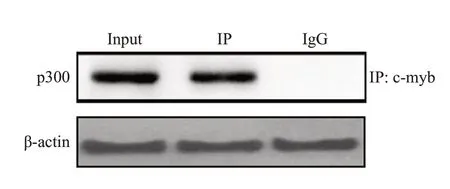
免疫共沉淀结果表明c-myb抗体能够提取到p300的蛋白成分,与input组结果类似,而IgG则无法获取,明确了p300与c-myb在RAW246.7核内的结合(图4)。
Western blotting法对10 个代表基因蛋白产物的表达水平验证结果显示与基因芯片检测结果吻合,即随着LPS刺激强度增加,p65与p300的表达逐步上升(图3A、B,<0.05)。
正态分布的定量资料均采用均数±标准差表示,多组间比较采用单因素方差分析,两两间比较采用最小显著差异法(LSD)。利用IBM SPSS Statistics 25软件进行分析,<0.05为差异有统计学意义。
Hermann Gretsch说:“我们不再提供那些让消费者购买一段时间后,就因为产品不实用、过时及退流行而弃置不用的产品”。他的设计观念在今天看来依旧如新。可见,实用功能是其设计的根本。无装饰是包豪斯的风格,因此陶瓷倾向简洁纯白。格罗佩斯生前最后设计委托案TAC 一号茶具,虽然线条柔和许多,仍属包豪斯风格,符合简洁几何外形、“形式追随功能”等原则。整套茶具以圆为主,握把像细长的门把,盖纽设计成钩子的造型,这并非是无用的装饰,极具功能性,使用时,用大拇指可以轻易地拉起壶盖和壶内的过滤器,避免倒水时,壶盖掉落得尴尬,使用非常方便。该设计造型简洁明快,没有任何多余的装饰,充分表达了包豪斯的精神。
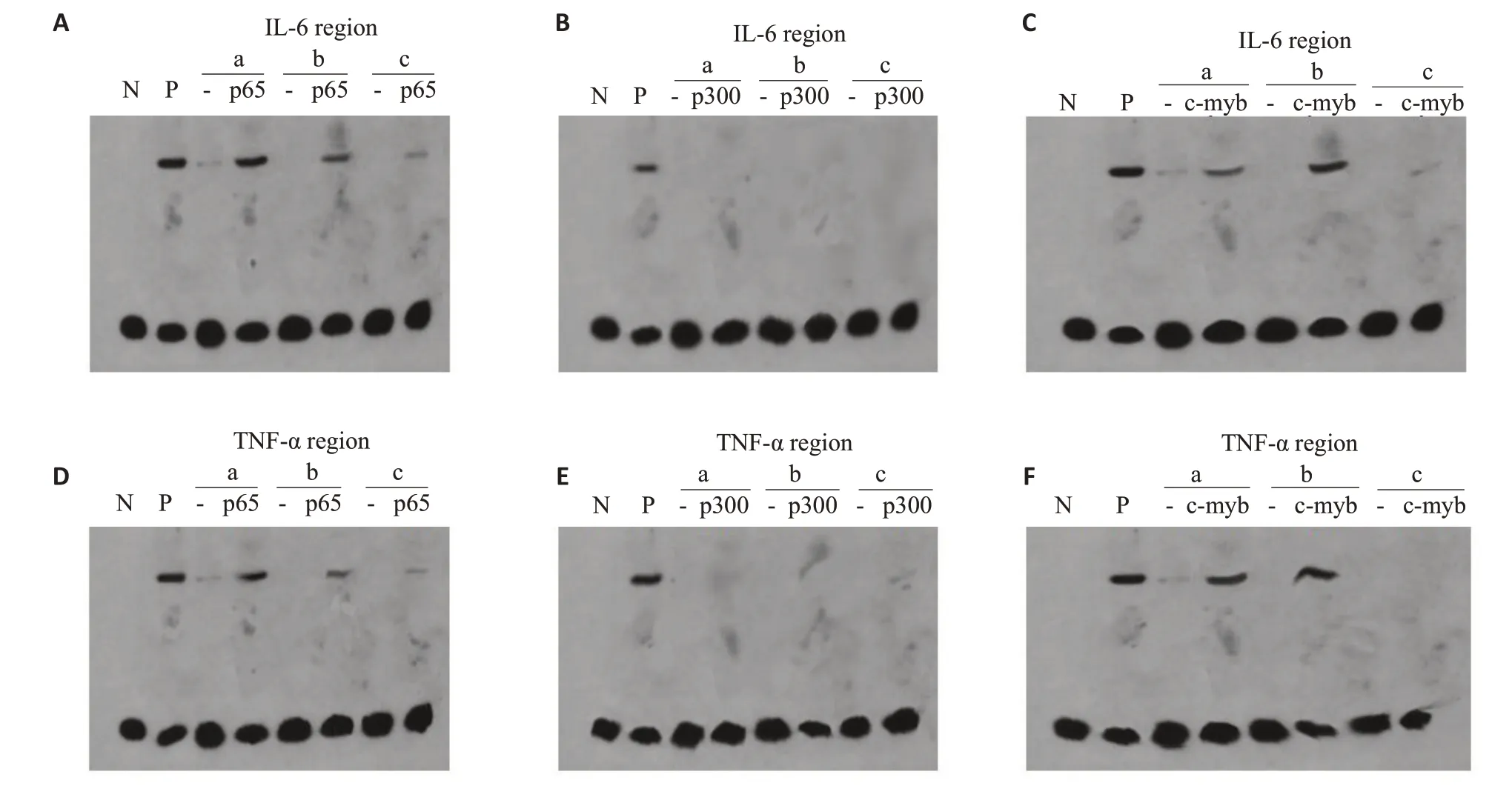
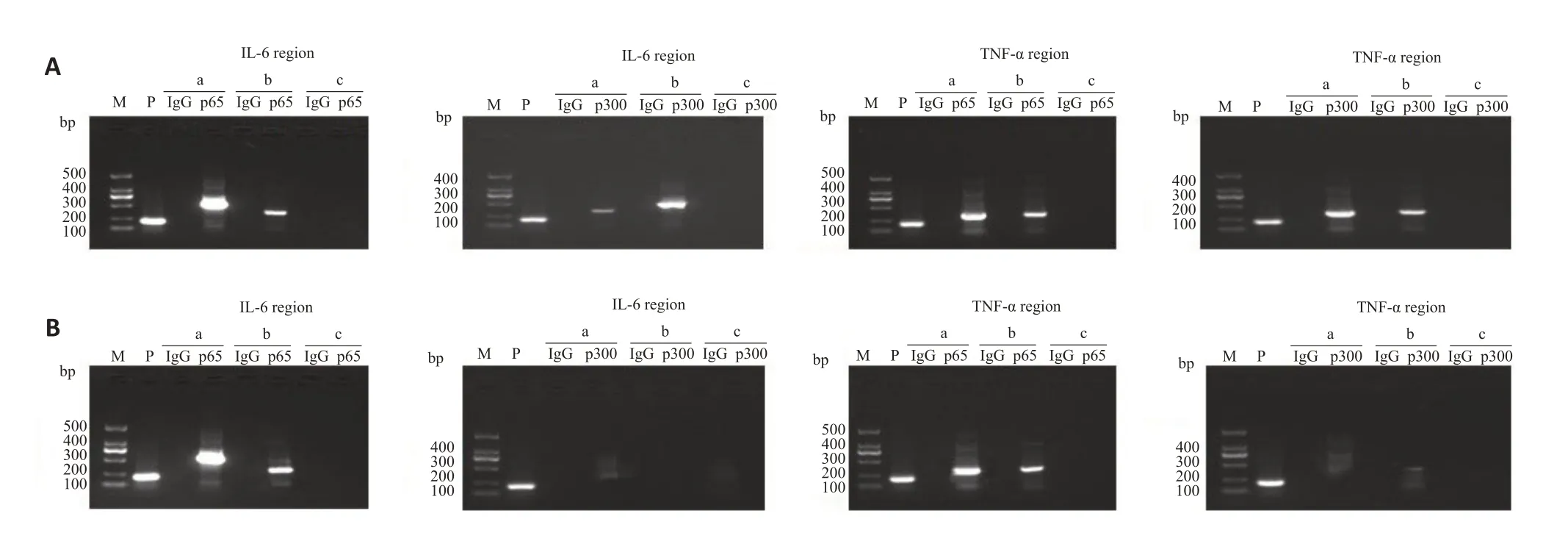
EMSA检测相应的p65、p300以及c-myb的纯蛋白提取物与IL-6以及TNF-α相应部位的结合情况,显示同阳性参照,以2基因启动子a,b区域为模板合成的有关探针与p65以及c-myb均能够结合,而内含子c区域则不能结合(图5A、C,图5D、F);且p300不能结合于上述a,b区域(图5B、E)。Chip-qPCR证实当c-myb存在时,p65以及p300抗体均能够沉淀和扩增得到炎症基因启动子a、b区域附近的片段(图6A,上排);c-myb被表达抑制时,p65抗体仍然能够沉淀和扩增而获得相应片段,但p300抗体不能获得(图6B,下排)。
2.3 P300诱导的乙酰化修饰参与LPS诱导的炎症介质合成
采用ELISA 法检测培基中IL-6 以及TNF-α质量浓度。在酶标包被板上设置标准品孔、待测样本孔,分别加入标准以及待测样本各10 μL,37 ℃下孵育30 min。酶标工作液洗板3 次后加入显色剂,37 ℃避光显色15 min,后加入终止液。采用酶标仪测定吸光度值,再根据标准曲线求得各待测孔的质量浓度。
2.3.2 p300可促进炎症基因启动子区域H3K27乙酰化,从而易化p65结合和激活转录 Chip-seq对各处理组炎症基因相应区域(IL-6为第5号染色体的30 003 000~30 015 000;TNF-α为第17 号染色体的35 195 000~35 200 000范围)p65、p300以及乙酰化H3K27结合水平的分析结果(图8A、B),ELISA对各组炎症介质合成水平的检测结果显示各干预组炎症基因启动子区域p65、p300和乙酰化H3K27的结合量以及IL-6和TNF-α表达水平的变化规律基本一致(图9)。相比于对照组,p300过表达质粒转染以及LPS刺激均可导致炎症基因启动子部位结合的p300和H3K27乙酰化修饰水平增加,从而促进p65与启动子的结合和炎症基因转录(<0.05)。然而c-myb 被表达抑制时,p300 过表达以及LPS刺激诱导的炎症基因启动子部位p65、p300以及乙酰化H3K27结合水平改变和炎症介质合成均被遏制(<0.05)。在p65干扰相关组别中,p65与启动子的结合以及炎症基因转录同样被遏制(<0.05),但未对p300的结合以及H3K27的乙酰化修饰水平造成影响。
随着越来越多的立法者加入其中,2015年国会要求NIH为阿尔茨海默病的研究准备一份“专业判断”的预算,这也是一份实现2025年目标所需要的愿望清单,该清单会绕过联邦预算程序,直接提交给总统和国会。此前,只有癌症和艾滋病的研究才能享受这种特殊待遇。
3 讨论
随着生物医学研究技术的不断进步,表观遗传修饰与人类疾病的关联被逐步揭示,很多不能够利用孟德尔遗传法则解释的发病现象,却可以被DNA甲基化以及组蛋白修饰等效应加以解释;且相关的修饰激动或者抑制策略,同样可能应用于临床疾病的干预,提示学术界需要更多从表观遗传修饰的角度来探讨相关疾病的发生机制。我们的前期研究发现,武陵山片区道地药材血藤果之中的提取物山姜素可激活组蛋白去乙酰化酶HDAC1以及DNA甲基转移酶DNMT3A,从而诱导炎症基因IL-6启动子部位的H3K9以及CpG二核苷酸分别发生去乙酰化和甲基化修饰,导致IL-6的合成抑制。以上发现表明相关表观遗传修饰介入了炎症介质表达的过程,因此在经典刺激物质LPS所造成的炎症激活过程中,表观遗传修饰可能同样扮演了重要角色。
在导致基因表达激活的表观遗传修饰当中,H3K27乙酰化修饰研究得比较深入,其多由辅助激活因子p300诱导。因为乙酰基团的加入干扰了核小体中碱性氨基酸与DNA之间的静电吸引力,使相邻核小体之间的聚集力减少,导致染色质开放,有利于转录因子结合于顺式作用元件,从而激活基因转录。本次研究中,我们利用LPS刺激RAW246.7,并采用小鼠表达谱芯片进行差异表达基因的“GO”分析,证实了LPS刺激可造成RAW246.7胞内炎症反应活化。联合WB技术,提示除了经典核因子p65,尚有p300的表达与LPS刺激强度呈显著关联,表明后者可能在LPS诱导的炎症介质合成过程中起到重要作用。
The azimuth DOA hk;l(t) and elevation DOA uk;l(t) can be expressed as
为了进一步揭示p300诱导炎症介质合成的分子机制,我们论证了p300 与炎症基因启动子存在结合现象。根据已有报道提示,p300独立存在时稳定性较差,多数情况下是与其它分子伴侣结合后才可定位于靶基因以发挥表达调节作用,其中报道较多的为CREB结合蛋白(CBP),据此推测p300/CBP复合物才是真正发挥乙酰化修饰作用的分子。还有研究报道在造血或肿瘤干细胞中,p300通过与辅助激活因子c-myb结合的方式以激活某些原癌基因表达,故p300/c-myb复合物可能与肿瘤的恶性分化有关。接下来我们利用PROMO数据库预测到IL-6以及TNF-α启动子部位均存在p65以及c-myb结合位点,但并无p300结合位点,故推测p300与炎症基因启动子的结合可能依赖于cmyb介导,而接下来的免疫共沉淀则恰好证实了p300与c-myb在RAW246.7内的结合。另外EMSA还证实了炎症基因启动子相关序列与p65以及c-myb蛋白提纯物存在结合现象,而p300 却不能结合;并且ChipqPCR亦证明当c-myb表达被抑制时,p300抗体不能沉淀和扩增得到相应的炎症基因启动子序列片段。综上,研究结果支持在RAW246.7内,p300通过与c-myb结合后再定位于炎症基因启动子部位,从而实现对基因转录的调控作用。
我们进一步采用过表达或干扰质粒转染策略以及WB和ELISA检测方法,联合热门的蛋白质-DNA互作实验技术-chip-seq,揭示了p300诱导的乙酰化修饰参与LPS 诱导的炎症介质合成过程。首先通过对RAW246.7转染p65、p300以及c-myb 过表达以及干扰质粒,证实了3者均不干涉对方表达。虽然转染p300过表达质粒不影响p65合成,但过表达的p300结合于炎症基因启动子部位并造成附近H3K27发生乙酰化修饰,从而易化p65与启动子结合和促进炎症介质合成,产生与LPS刺激类似的效果。同时发现当c-myb缺失时,p300与炎症基因启动子的结合、H3K27的乙酰化修饰以及p65的结合水平均被遏制,因此逆转了LPS刺激或p300过表达质粒对炎症介质合成的促进效果。另外在p65干扰相关组别中,p65与启动子的结合以及炎症基因合成均被抑制,但却未对p300的结合以及H3K27的乙酰化修饰水平造成影响。综上我们推测,LPS诱导的p65以及p300合成均参与了炎症介质的表达调控,但两者的作用机制存在较大差异:p65结合于炎症基因启动子部位后可直接发挥转录激活作用;而p300需要在cmyb介导下才能结合,进一步的转录激活效应则依赖于乙酰化H3K27后造成的染色质结构松弛,使p65更易结合而得以实现。
综上所述,本研究证实了p300在LPS诱导RAW246.7炎症介质合成过程中起到了积极作用,提示逆转p300与炎症基因启动子的结合或干预其乙酰化修饰能力,同样可能成为逆转与LPS关联的炎症失衡的有效方法。鉴于巨噬细胞并非人体内唯一的炎症细胞种类,下一步还应考虑在中性粒细胞以及肥大细胞等对象中开展类似研究,探讨p300是否通过同样机制介导LPS诱导的炎症介质合成;此外还应利用疾病动物模型来验证上述作用机制的可重复性,为证明p300在临床炎症紊乱领域的干预价值提供更多可靠依据。
猜你喜欢
中国农业科学(2022年16期)2022-09-19
中国农业科学(2022年15期)2022-08-09
装备维修技术(2022年7期)2022-07-01
中学生数理化·高一版(2021年12期)2021-09-05
电脑报(2020年40期)2020-11-06
理科考试研究·高中(2019年7期)2019-09-17
电脑知识与技术(2018年19期)2018-11-01
现代农业科技(2018年13期)2018-10-20
山东工业技术(2017年1期)2017-01-24
中国信息化周报(2014年4期)2014-02-17
