
兴安落叶松树轮宽度指数与归一化植被指数的关系研究
2022-04-26 03:24张楠郭雪梅张冬有
森林工程 2022年1期
张楠 郭雪梅 张冬有







摘 要:為探究兴安落叶松树木径向生长与归一化植被指数(Normalized Difference Vegetation Index,NDVI)的关系,从而了解树木生长动态,本文运用1973—2018年塔河采样点的兴安落叶松、1982—2018年GIMMS NDVI(Global Inventory Modelling and Mapping Studies, 全球植被指数变化研究)数据集与研究区的气候因子(降水量、月平均气温、月平均最低气温、月平均最高气温)进行相关分析。研究结果表明,树轮宽度指数与6月、7月、6—7月平均气温显著负相关(P<0.05),与6月、7月、6—7月平均最高气温显著负相关(P<0.05);NDVI与上年12月平均最低气温、月平均气温显著负相关(P<0.05),与5月平均气温、平均最高气温、7月平均气温显著正相关(P<0.05),NDVI与7月、8月、7—8月平均最高气温极显著正相关(P<0.01);树轮宽度指数与当年7月NDV相关关系不显著,与上年7月NDVI显著正相关。从而得出 ,6—7月平均最高气温是树木径向生长的主要控制因子,7—8月平均最高气温是树木冠层生长的主要控制因子,主干生长与冠层生长受控因子不同,冠层生长状况与主干生长状况不同。
关键词:NDVI;树轮宽度指数;气候因子;兴安落叶松;树木生长
中图分类号:S716.3 文献标识码:A 文章编号:1006-8023(2022)01-0001-08
A Study on the Relationship between Tree Ring Width Index
and NDVI of Larix gmelinii
ZHANG Nan, GUO Xuemei, ZHANG Dongyou*
(Heilongjiang Province Key Laboratory of Geographical Environment Monitoring and Spatial Information Service in Cold
Regions, Harbin Normal University, Harbin 150025, China)
Abstract:To explore the relationship between radial growth and NDVI (Normalized Difference Vegetation Index) of Larix gmelinii, and to provide reference for understanding tree growth dynamics, based on Larix gmelinii at Tahe sampling point from 1973 to 2018, GIMMS (Global Inventory Modelling and Mapping Studies) NDVI data set from 1982 to 2018 and climate factors (precipitation, monthly mean temperature, monthly mean minimum temperature, monthly mean maximum temperature) was analyzed in this paper. Results showed that the tree ring width index was significantly negatively correlated with the mean temperature in June, July and June-July (P<0.05), and significantly negatively correlated with the mean maximum temperature in June, July and June-July (P<0.05). NDVI was significantly negatively correlated with the mean minimum temperature and monthly mean temperature in December last year (P<0.05), significantly positively correlated with the mean temperature in May, the mean maximum temperature in May and the mean temperature in July (P<0.05), extremely positively correlated with the mean maximum temperature in July, August and July-August (P<0.01). There was no significant correlation between tree ring width index and NDV in July of that year, but a significant positive correlation between tree ring width index and NDVI in July of last year. It can be concluded that the mean maximum temperature from June to July was the main controlling factor of tree radial growth, and the mean maximum temperature from July to August was the main controlling factor of tree canopy growth. The control factors of trunk growth and canopy growth were different, and the growth status of canopy was different from that of trunk.
Keywords:NDVI; tree ring width index; climatic factor; Larix gmelinii; tree growth
0 引言
在全球气候变化下的大环境下,陆地生态系统如何变化及其对气候环境的响应作用是气候变化领域研究的核心问题之一[1]。植被是陆地生态系统重要组成部分[2],生长受到气候、地貌等因子的影响。研究植被冠层生长与径向生长间的关系,有利于明确气候变化背景下树木生长的动态变化,更好地了解树木的生理机制以及为气候变化提供参考依据[3]。归一化植被指数(Normalized Difference Vegetation Index, NDVI)是植物叶面由红光(植被吸收)和近红外(植被强烈反射)2个波段反射所合成的指数,能够反映冠层的繁茂程度和植被覆盖的动态变化[4]。且大量的研究结果显示,NDVI与植被覆盖度有较高程度的相关性,因此可用 NDVI来分析区域植被覆盖度[5-6]。
近年来已经有很多树轮宽度与NDVI的关系研究。闫平等[1]研究发现树木径向生长与冠层生长受控因子不同,夏季温度与树轮宽度序列的关系为显著正相关,上一年以及当年7月降水与NDVI序列显著正相关;柳凯茜等[7]研究发现树木的整轮宽度和早材宽度、晚材宽度与气候因子呈现不同的关系,与前一年NDVI呈显著的正相关关系,晚材宽度与当年4—5月NDVI呈显著的负相关关系;张旭等[8]研究发现小兴安岭树轮指数与当年7月NDVI显著正相关,并且重建了过去NDVI变化系列,为过去发生的干旱事件提供依据;郭艳飞等[9]研究发现森林上限与下限树轮宽度指数与生长季、非生长季NDVI的相关性不同;汪青春等[10]分析了青海湖布哈河流域NDVI与树轮宽度指数的关系,并重建了8月份NDVI的千年序列;贾飞飞等[11]研究腾格里沙漠南缘树轮宽度指数与NDVI的关系,重建1804—2009年NDVI序列,为过去发生的极端旱灾事件提供了参考依据;Vicente-Serrano等 [12]将西班牙不同环境条件下各种森林类型的树木年轮生长和净初级生产力结合起来,与NDVI数据相结合,探究不同森林生物群落的树木年轮生长和植被活动的变化;Carlon等[13]采取墨西哥米却肯州火山附近树木,并且评估了海洋大气指数和气候响应对树木年径向增长的影响;He 等[14]运用祁连圆柏的树轮年表,研究树轮宽度指数、NDVI和气候资料之间的关系,并且重建了德令哈过去一千年的8月NDVI。
采样点位于大兴安岭地区,已有很多学者对该地区的树木年轮与气候关系进行了研究[15-17]。这些研究大多是树轮宽度指数与气候因子的关系,该地区树轮宽度指数与归一化植被指数的关系研究较少,本文研究兴安落叶松树轮宽度指数、NDVI、气候因子的内在联系,为陆地生态系统对全球变化的响应及影响提供参考依据。
1 研究区概况和数据源
1.1 研究区概况
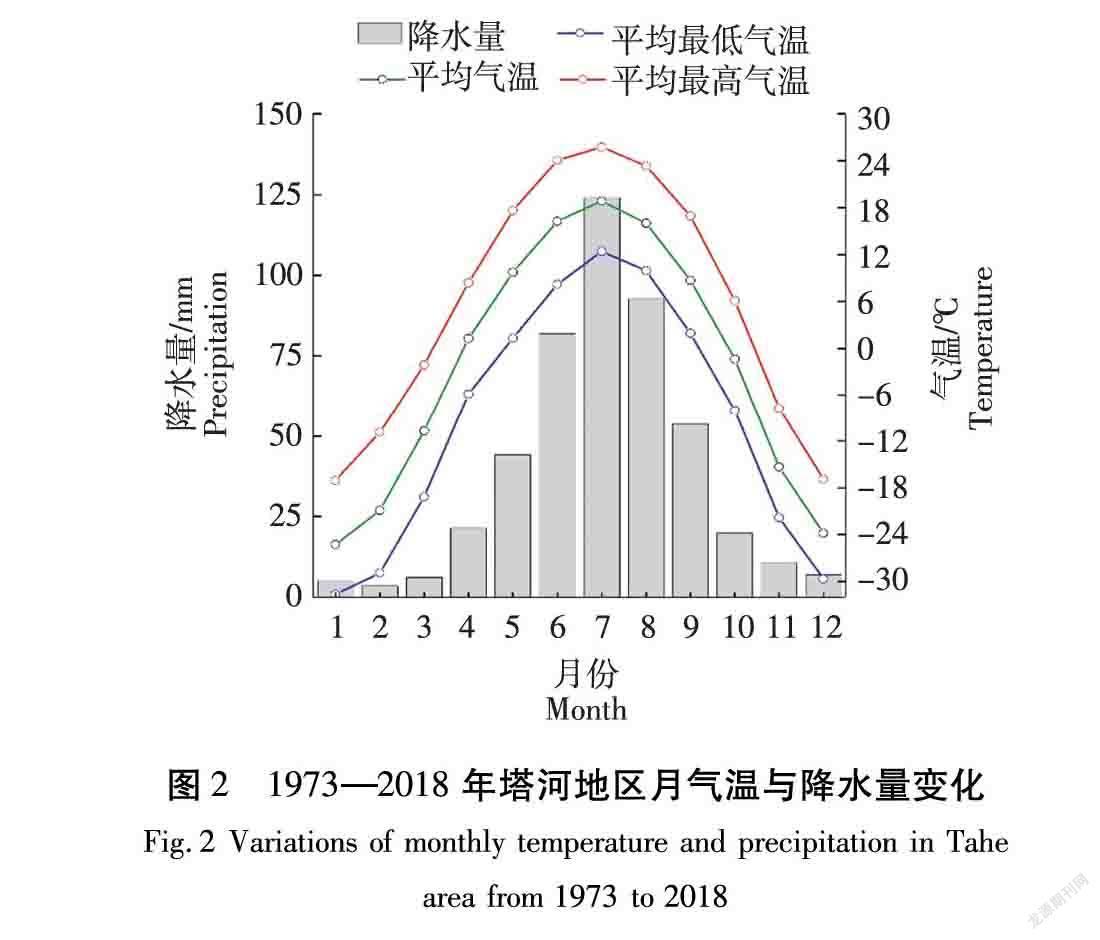
本研究选择区域是大兴安岭地区的塔河县,研究区域如图1所示。大兴安岭地处我国东北部,气候寒冷,是我国多年冻土主要发育的地区之一,由于大陆和海洋季风交替性影响,导致小气候变化多端,局部气候有显著差异,在极地大陆气团控制下,冬季气温极低、干燥而漫长。由于东北地区最近150年来的显著气候变暖和清朝开禁政策以来强烈的人为活动影响,东北地区冻土和寒区环境已经产生了显著变化[18]。塔河县位于黑龙江省北部、伊勒呼里山北麓,地理坐标为:123°19′~125°48′ E,52°09′~52°23′ N。塔河县地处北温带,属寒温带大陆性季风气候,年平均气温2.4 ℃,极端最高气温37.2 ℃(1992年),极端最低气温-45.8 ℃(1980年)。塔河县气候主要特征为:塔河县降水主要集中在7月、8月,7月、8月也是一年中最热的2个月,具有雨热同期的特点。一年中冬季的最低温度可达到-31.6 ℃,夏季的最高温度25.7 ℃,气温年较差大,月平均最高气温与月平均最低气温相差15 ℃左右。根据已有的气候信息,1973—2018年,塔河地区气温呈升高趋势。年平均降水量大于400 mm,属于半湿润地区,降水量最多的年份为1982年,年总降水量可达到715 mm,降水量最少的年份为1986年,年总降水量317 mm,气温、降水的变化并不是很稳定。为更直观地理解该地气候特征,制作该地区月均温与月降水量、年均温与年降水量示意图,分别如图2和图3所示。植被以兴安落叶松构成的明亮针叶林为主, 主要林型有杜香-兴安落叶松林、杜鹃-兴安落叶松林、草类-落叶松林、泥炭藓-真藓-落叶松林等[19-20]。主要植物有的白桦 (Betula platyphylla) 、山杨(Populus davidiana) 、杜香(Ledum palustre) 、兴安杜鹃 (Rhododendron dauricum)和越橘 (Vaccinium) 等[21] 。
1.2 数据的获取及处理
NDVI数据采用荷兰皇家气象研究所提供的GIMMS NDVI(Global Inventory Modelling and Mapping Studies NDVI),全球清單建模和制图研究NDVI数据(http://climexp.knmi.nl)。时间序列为1982—2018年,数据均已经过几何校正、辐射校正和大气校正等预处理。提取包含采样点(52°22′08″~55°22′18″ N,124°46′47″~124°46′57″ E)的NDVI最大值数据集,并且生成1982—2018年1—12月份逐月NDVI序列以及5—9 月份生长季NDVI序列。气象数据来源于塔河(52°21′ N,124°43′ E)气象站,包含月平均气温、月平均最高气温、月平均最低气温和月降水量数据。选取NDVI数据与气候数据重合年份2000—2018年进行皮尔逊(Person)相关分析,树轮数据选取与气候数据重合的1973—2018年进行相关分析。
2 研究方法
2.1 树木年轮样本采集及其年表特征
兴安落叶松是本研究区的优势树种之一。该树种为强阳性树种,适应能力较强,寿命长,耐寒、耐旱力强[22]。该树种对气候变化极其敏感,每年都表现出明显的年轮宽度变化[23]。取塔河地区落叶松的树木年轮样本,在该地区分别采取20株,采取樣本时使用内径为5.15 mm的生长锥于大约胸径高处钻取样芯,一株树钻取两根样芯,分别从东部和北部钻取,样芯经过晾晒、干燥、固定和打磨等后期处理。利用LINTAB树木年轮分析仪对树木年轮宽度进行测量,其次利用准确性检验(COFECHA)计算机程序对测量后的树轮样本序列进行质量检验和控制,消除定年和测量过程中的一些错误,剔除生长异常和相关性较差的样本序列[24]。利用去除趋势(ARSTAN)计算机程序根据趋势的形式,通过拟合负指数曲线或水平线来消除交叉日期树轮序列的趋势。如果年轮系列显示年龄趋势,则使用负指数函数,如果年轮系列不显示年龄趋势,则使用水平线[25]。得到标准(STD)年表、差值(RES)年表、回归(ARSTAN)年表。树轮宽度指数是年轮实际测量宽度值与期望值的比值,在建立年表的过程中,通常采用统计学方法对测量的年轮宽度序列进行曲线拟合,由遗传因子支配的、随树木年龄导致的树木径向生长减缓趋势以及其他非限制因子造成的树木生长波动可以消除,去除这些因素后,获取主要限制因子制约造成的年轮宽度变化序列。落叶松树轮宽度指数与样本量如图4所示。树轮宽度指数代替了原年轮宽度序列,做进一步年轮气候学分析。
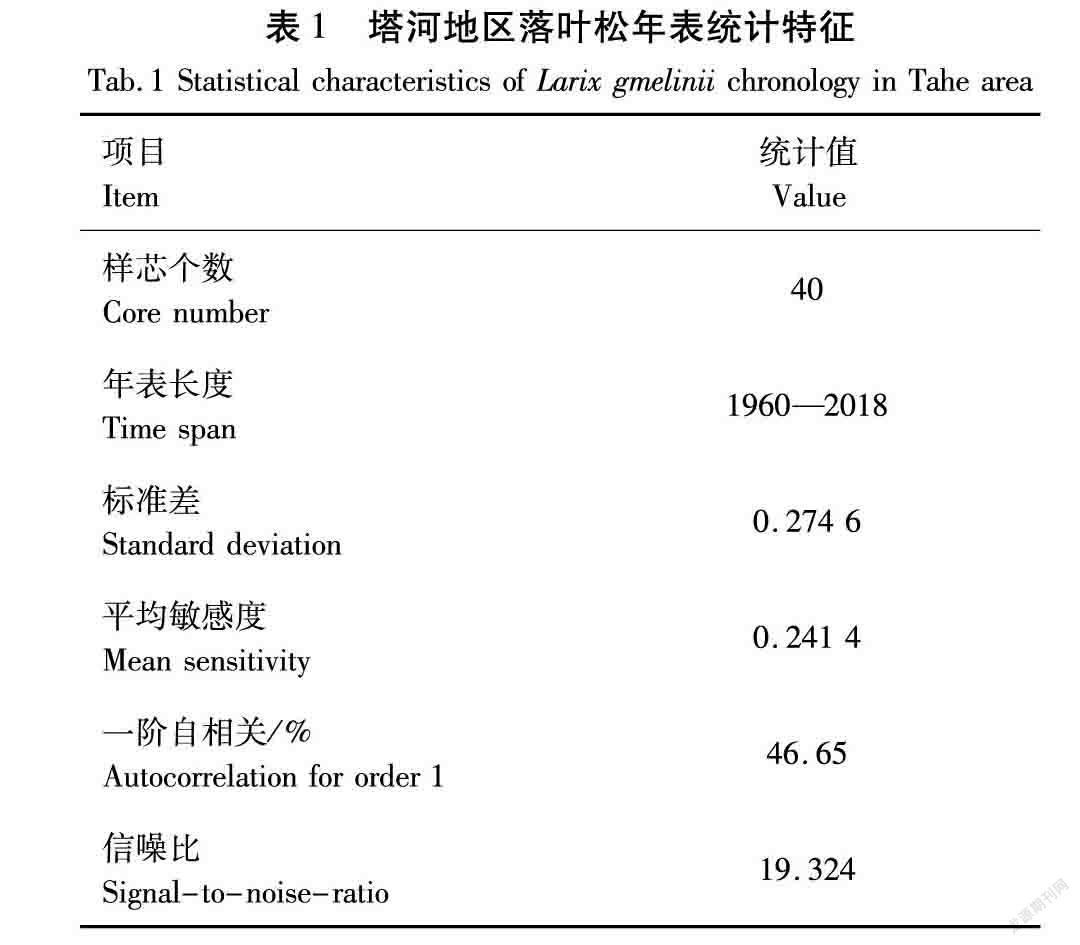
从塔河地区落叶松年表统计特征发现,年表统计特征见表1,塔河地区的落叶松的信噪比和平均敏感度均较高,敏感度值反映了所含的气候信息含量和气候因子对树木生长的限制作用,敏感度越大,气候信息含量越多,相应的非气候噪音越少[26-27]。信噪比越大,气候信息含量越大,树木基本不受人为活动的影响,很好地记录并反映了周围环境变化。
2.2 相关分析法
确定树木径向生长与气候要素之间的相关性。2个变量之间的Person相关系数定义为2个变量之间的协方差和标准差的商,公式为:
P=∑ni=1(xi-x)(yi-y)∑ni=1(xi-x)2∑ni=1(yi-y)2。
(1)
式中:P为相关系数;xi与 yi分别代表了x和y在第i年的值;x和y表示平均值;n为样本容量。
相关系数值为-1~1,正值代表变量间呈正相关关系,负值代表变量间呈负相关关系,相关系数的绝对值越接近于1,变量间相关性越强。
3 研究结果
3.1 树轮径向生长与气候因子的相关分析
因为考虑到树木年轮生长存在滞后效应,所以选取前一年6月至当年10月的气候因子进行分析,列出相关系数显著月份。利用大兴安岭地区塔河树轮宽度标准年表与研究区的气候因子进行相关分析(降水量、平均最低气温、平均气温和平均最高气温),分析结果如图5所示。由图5得出,树轮年表与降水量的相关性并不显著,表明塔河的落叶松生长受降水影响较弱。树轮年表与上一年7月份的月平均最低气温达到了显著负相关(r=-0.296,P<0.05),与上一年9月份(r=-0.329,P<0.05)、当年6月(r=-0.318,P<0.05)、7月(r=-0.317,P<0.05)的平均气温显著负相关,与6—7月平均气温显著负相关(r=-0.413,P<0.01),与上一年9月(r=-0.348,P<0.05)、当年2月(r=-0.315,P<0.05)、6月(r=-0.382,P<0.05)、7月(r=-0.351)的月平均最高气温显著负相关,与当年6—7月平均最高气温显著负相关(r=-0.476,P<0.01)。
3.2 NDVI与气候因子的相关分析
图6为该研究区内1982—2018年NDVI逐月变化,在7月出现最大值,12月和1月出现最小值。兴安落叶松展叶子期在5—10月,符合逐月变化曲线。该地区NDVI逐月变化符合温带大陆性气候特征,降水量主要集中在夏季,且夏季温度高,树木光合作用旺盛。
气候与NDVI是相互反映的关系,植物生长过程中会受不同气候因子的作用和影响,生长状况与树木生长环境中的各因子的综合作用有关[7]。利用生长季NDVI均值与研究区气候因子(月降水量、平均最低气温、平均气温和平均最高气温)进行相关分析,分析结果如图7所示,NDVI与降水量的相关性并不显著。与12月(r=-0.471,P<0.05)、2月(r=-0.399,P<0.05)平均最低气温显著负相关,与5月平均最低气温显著正相关(r=0.335,P<0.05);与12月平均气温显著负相关(r=-0.362,P<0.05),与5月(r=0.394,P<0.05)、7月(r=0.368,P<0.05)平均气温显著正相关,与8月(r=0.465,P<0.01)、7—8月(r=0.512,P<0.01)平均气温极显著正相关;与5月平均最高气温显著正相关(r=0.341,P<0.05)与7月(r=0.457,P<0.01)、8月(r=0.555,P<0.01)、7—8月(r=0.584,P<0.01)平均最高气温极显著正相关。
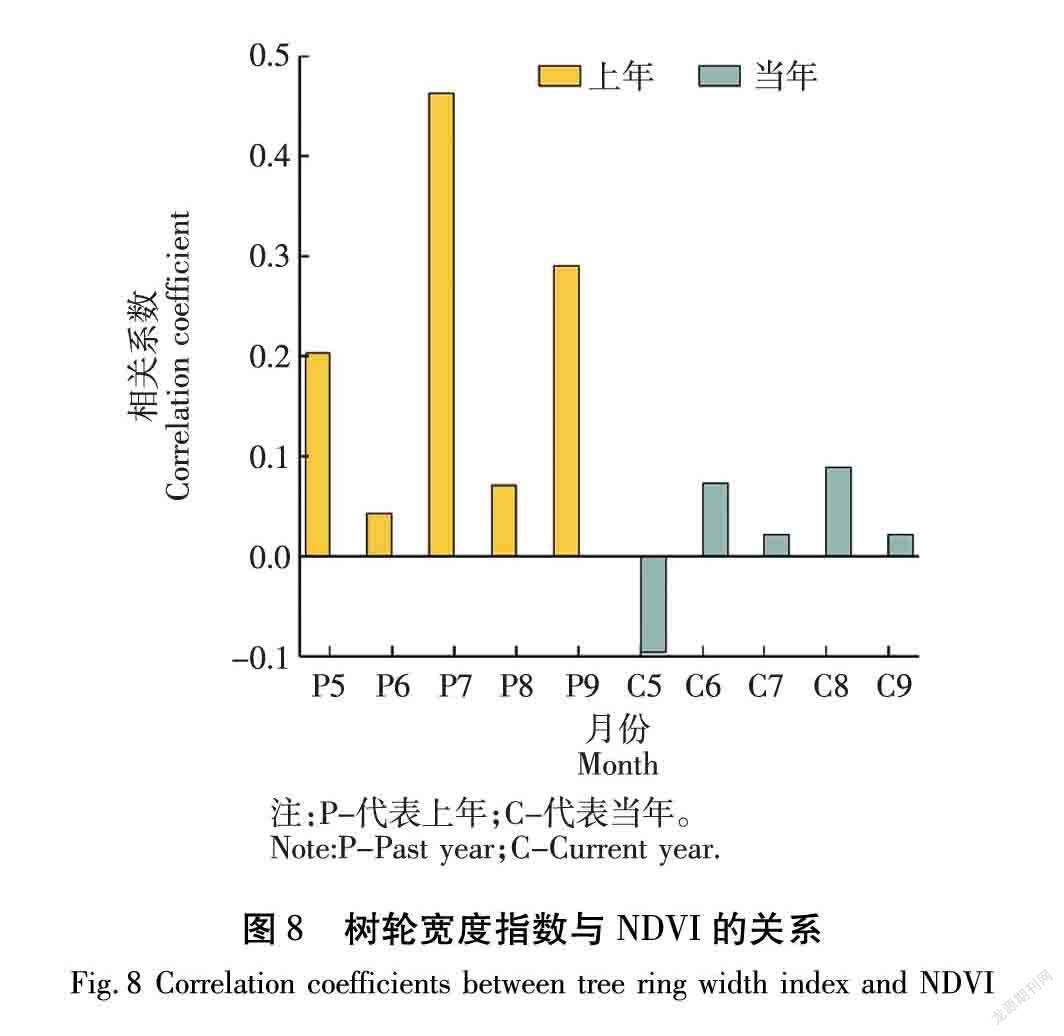
3.3 树木径向生长与NDVI的相关分析
兴安落叶松树高的生长期从5月下旬到9月中下旬。但快速生长在6月中旬至7月下旬[28]。而兴安落叶松展叶在5月中旬,落叶在10月[29]。本文研究结果与前人的研究结果不同,树轮宽度指数与当年7月的NDVI值相关性不高,与前一年7月的NDVI在95%置信度水平下正相关(r=0.463,P<0.05)。兴安落叶松的树木冠层生长与径向生长过程存在差异,但二者存在紧密的联系。植物叶片进行光合作用合成营养成分,然后通过筛管运送到植物的各个部位,造成树木的长高和加粗生长。在树的生长过程中,细胞逐渐形成层,在树叶以及枝干部分进行着细胞的有丝分裂,这样细胞也会逐渐增多,叶子茂盛,主干粗壮。由于从根部吸收的养分和从叶片吸收的养分运输速度不同,导致树轮宽度指数与当年7月NDVI相关不显著。
4 讨论
4.1 树木径向生长、NDVI与气候要素的响应
大兴安岭地区的主要降水集中在6—8月份,这3个月的降水量占全年降水量的70%。但这 3个月的降水量与树轮宽度指数、NDVI的相关性并不显著,与郭艳飞等[9]、 王丽丽等[16]研究结果相同。造成这一情况的主要原因可能是:7—8月細胞的分列与伸长已经基本完成,新叶片也发育成熟,进入了光合积累的阶段,树木的生长主要体现在晚材木质细胞的加厚上[16],晚材宽度在总轮宽度中占比较小。对于林冠层而言,6—8月生长所需要的水分主要由5月份供应,所以与5月降水量的相关系数较其他月份的相关系数高。6—8月的降水量对冠层生长影响较弱,但是对土壤水分的存储影响较大,影响来年的生长季前期阶段[14]。
兴安落叶松径向生长、冠层生长与气候因子的关系既存在一定的共性,又表现一定的差异性。6—7月平均最高气温是塔河落叶松径向生长的主要限制因子,7月是全年温度最高的月份,树木的各项生理活动进入盛期,如果此时温度过高,土壤中的水分蒸发加剧,导致可利用的水分减少,使树木根系得不到充足的水分;如果温度过低,树木的各项生理活动就会变缓慢,也会使树木的生长速度随之变慢[22]。所以树轮宽度指数与平均最低气温的相关性很弱,而与平均气温、平均最高气温的相关性强。NDVI与上年12月平均最低气温、平均气温、2月平均最低气温呈现显著负相关关系。12月份开始进入冬季时期,一直到2月份,由于温度过低,林冠层叶片不再进行光合作用,树叶基本全部凋落。5月平均气温是落叶松冠层生长的限制因子,5月份的平均气温促进树木冠层生长。5月温度的升高可促进落叶松叶片发育,并且对光合作用起着重要作用。这与Sano等[23]的研究结果相同。NDVI与7月、8月、7—8月平均最高气温极显著相关,7—8月是温度最高的月份,也是叶片绿度最高的时期,此时叶片细胞分裂最为旺盛,光合作用较为强烈。与这与Coulthard等 [30]研究区域的高海拔林区的NDVI呈现相同结果。
4.2 树木生长与NDVI的响应
NDVI 与树轮年表间的相关性有着明确的生理基础[2]。植物的生长发育受外界环境和内在因素的制约,具有一定的阶段性和季节性,塔河地区落叶松的树轮宽度与上年生长季的NDVI月平均值呈正相关,5—9月具有适合植被生长的环境条件,在此阶段生长速率增大,并且到最快,产生较多的营养物质,之后进入生长季的末期。植物没有循环系统,树木径向生长的养分来源于叶片进行光合作用后产生的有机质,以及土壤中的养分,共同作用产生的。树叶吸收阳光进行光合作用,在叶片中合成葡萄糖,经过生物反应转化为树木必需的营养物质。持续地向树木的器官运输所合成的营养物质,为树木的生长发育提供能量或者存储起来[7,31]。根茎将上一年多余的物质贮藏起来,用于下一年树木生长。因为落叶松径向生长与冠层生长受气候因子影响不同,导致塔河落叶松树木年轮的生长与上一年7月NDVI的月平均值显著正相关,此研究结果与柳凯茜等[7] 、商志远等[32]研究结果相同。
本文只研究了月平均气温、月平均最低气温、月平均最高气温和月降水量对树木生长的影响,没有考虑到其他气候要素对于树木生长的影响,比如日照、相对湿度等,并且NDVI值代表的是植被覆盖度,因为研究区内既有林地又有草地,植被类型也有一定的差异,所以对树轮宽度指数与NDVI关系进行研究时,也会受到一定的干扰。
5 结论
大兴安岭地区植被覆盖率较高,被列入我国国有重点林区名单之中。在全球气候变化的背景下,研究塔河县落叶松生长对气候因子的响应,不仅为树木生长动态提供参考依据,而且长时间序列的树轮宽度指数为研究塔河地区过去的长期生态环境变化具有重要意义,也有助于预测气候变化对生态系统和人类社会的影响。
NDVI与植被覆盖度有很强的相关性,植物冠层的背景影响可以由NDVI反映。所以本文利用1982—2018年归一化植被指数、1973—2018年树轮宽度指数与月平均气温、月平均最低气温、月平均最高气温、月降水量进行相关分析,探讨研究兴安落叶松树木径向生长与NDVI的关系,得到以下结论。
(1) 塔河树木年轮与6月、7月、6—7月份的月平均最高气温呈显著负相关,树木径向生长的受控因子是6—7月平均最高气温。
(2) NDVI与7—8月最高气温呈极显著正相关,树木冠层生长的受控因子是7—8月平均最高气温。
(3) 塔河地区落叶松的生长受到降水的影响较弱,但是不排除树木生长不受降水控制。
(4) 塔河地区的落叶松与当年7月NDVI相关性不显著,与上年7月NDVI显著相关。
【参 考 文 献】
[1]闫平,徐健楠,菊花,等.云南三江并流地区树轮宽度指数与NDVI关系研究[J].林业资源管理,2019,48(6):42-48.
YAN P, XU J N, JU H, et al. A study on the relation between tree ring width index and NDVI in the three parallel rivers region, Yunnan Province[J]. Forest Resources Management, 2019, 48(6): 42-48.
[2]路俊伟,王乃昂,侯迎,等.石羊河上游地区树轮年表与NDVI关系分析[J].干旱区研究,2012,29(4):667-673.
LU J W, WANG N A, HOU Y, et al. Analysis on the correlation between NDVI and tree-ring width chronologies in the upper reaches of the Shiyang river[J]. Arid Zone Research, 2012, 29(4): 667-673.
[3]李明明,李刚.植被类型对树轮重建贺兰山NDVI的影响[J].甘肃农业大学学报,2020,55(5):152-161.
LI M M, LI G. Effects of vegetation type on tree ring based NDVI reconstruction for Helan Mountains[J]. Journal of Gansu Agricultural University, 2020, 55(5): 152-161.
[4]GAMON J A, FIELD C B, GOULDEN M L, et al. Relationships between NDVI, canopy structure, and photosynthesis in three Californian vegetation types[J]. Ecological Applications, 1995, 5(1): 28-41.
[5]惠刚盈,胡艳波,赵中华.结构化森林经营研究进展[J].林业科学研究,2018,31(1):85-93.
HUI G Y, HU Y B, ZHAO Z H. Research progress of structure-based forest management[J]. Forest Research, 2018, 31(1): 85-93.
[6]胡艳波,惠刚盈.优化林分空间结构的森林经营方法探讨[J].林业科学研究,2006,19(1):1-8.
HU Y B, HUI G Y. A discussion on forest management method optimizing forest spatial structure[J]. Forest Research, 2006, 19(1): 1-8.
[7]柳凯茜,曾彪,杨太保,等.重庆青华林场树轮宽度指数分析及其与NDVI的关系[J].水土保持研究,2015,22(3):170-175.
LIU K X, ZENG B, YANG T B, et al. Analysis of tree ring width index and its relationship with NDVI in Qinghua forest of Chongqing[J]. Research of Soil and Water Conservation, 2015, 22(3): 170-175.
[8]張旭,宋文琦,赵慧颖,等.小兴安岭南部臭冷杉红松林树轮指数记录的7月NDVI变化[J].北京林业大学学报,2018,40(12):9-17.
ZHANG X, SONG W Q, ZHAO H Y, et al. Variation of July NDVI recorded by tree-ring index of Pinus koraiensis and Abies nephrolepis forests in the southern Xiaoxing’an Mountains of northeastern China[J]. Journal of Beijing Forestry University, 2018, 40(12): 9-17.
[9]郭艳飞,甘淼,杨美琳,等.天山东部树木径向生长与NDVI的关系[J].干旱区资源与环境,2018,32(3):176-180.
GUO Y F, GAN M, YANG M L, et al. Relationship between tree radial growth and normalized difference vegetation index in eastern Tianshan Mountains[J]. Journal of Arid Land Resources and Environment, 2018, 32(3): 176-180.
[10]汪青春,时兴合,刘义花,等.青海湖布哈河流域树轮宽度指数与NDVI植被指数的关系[J].冰川冻土,2012,34(6):1424-1432.
WANG Q C, SHI X H, LIU Y H, et al. Relationship between tree ring indices and vegetation index(NDVI) in Buha river basin, Qinghai[J]. Journal of Glaciology and Geocryology, 2012, 34(6): 1424-1432.
[11]贾飞飞,鲁瑞洁,高尚玉.腾格里沙漠南缘哈思山树轮记录的NDVI变化[J].第四纪研究,2018,38(2):327-335.
JIA F F, LU R J, GAO S Y. Variations of NDVI recorded by tree-ring in the Hasi mountain, the southern margin of the Tengger desert[J]. Quaternary Sciences, 2018, 38(2):327-335.
[12]VICENTE-SERRANO S M, MARTIN-HERNANDEZ N, CAMARERO J J, et al. Linking tree-ring growth and satellite-derived gross primary growth in multiple forest biomes. Temporal-scale matters[J]. Ecological Indicators, 2020, 108: 105753.
[13]CARLN ALLENDE T, VILLANUEVA DIAZ J, SOTO CASTRO G, et al. Tree rings as indicators of climatic variation in the Trans-Mexican Volcanic Belt, central Mexico[J]. Ecological Indicators, 2021, 120: 106920.
[14]HE J C, SHAO X M. Relationships between tree-ring width index and NDVI of grassland in Delingha[J]. Chinese Science Bulletin, 2006, 51(9): 1106-1114.
[15]杨婧雯,张秋良,宋文琦,等.大兴安岭兴安落叶松和樟子松径向生长对气候变化的响应差异[J].应用生态学报,2021,32(10):3415-3427.
YANG J W, ZHANG Q L, SONG W Q, et al. Response differences of radial growth of Larix gmelinii and Pinus sylvestris var. mongolica to climate change in Daxing’an Mountains, Northeast China[J]. Chinese Journal of Applied Ecology, 2021, 32(10): 3415-3427.
[16]王丽丽,邵雪梅,黄磊,等.黑龙江漠河兴安落叶松与樟子松树轮生长特性及其对气候的响应[J].植物生态学报,2005,29(3):280-285.
WANG L L, SHAO X M, HUANG L, et al. Tree-ring characteristics of Larix gmelinii and Pinus sylvestris var. mongolica and their response to climate in Mohe, China[J]. Acta Phytoecologica Sinica, 2005, 29(3): 280-285.
[17]梁媛.大兴安岭兴安落叶松径向生长对气候变化响应[D].哈尔滨:东北林业大学,2014.
LIANG Y. The response of the growth of Larix gmelinii to the climate change in Great Khingan[D]. Harbin: Northeast Forestry University, 2014.
[18]常晓丽,金会军,何瑞霞,等.中国东北大兴安岭多年冻土与寒区环境考察和研究进展[J].冰川冻土,2008,30(1):176-182.
CHANG X L, JIN H J, HE R X, et al. Advances in permafrost and cold regions environments studies in the Daxing’anling (Dahing’an) mountains, northeastern China[J]. Journal of Glaciology and Geocryology, 2008, 30(1): 176-182.
[19]王飞,张秋良,马秀枝,等.渐伐对草类-兴安落叶松林下植被多样性的影响[J].生态环境学报,2012,21(5):813-817.
WANG F, ZHANG Q L, MA X Z, et al. Effect of successive cutting on the diversity of understory vegetation in herbage-Larix gmelinii forest[J]. Ecology and Environmental Sciences, 2012, 21(5): 813-817.
[20]王敏,周润惠,余飞燕,等.不同林龄桉树人工林林下物种多样性和生物量的动态变化[J].植物研究,2021,41(4):496-505.、
WANG M, ZHOU R H, YU F Y, et al. Dynamic changes of undergrowth species diversity and biomass of Eucalyptus robusta plantations at different ages[J]. Bulletin of Botanical Research, 2021, 41(4): 496-505.
[21]周梅.大興安岭森林生态系统水文规律研究[M].北京:中国科学技术出版社,2003.
ZHOU M. China science and technology press[M]. Beijing: China Science and Technology Press, 2003.
[22]张先亮,崔明星,马艳军,等.大兴安岭库都尔地区兴安落叶松年轮宽度年表及其与气候变化的关系[J].应用生态学报,2010,21(10):2501-2507.
ZHANG X L, CUI M X, MA Y J, et al. Larix gmelinii tree-ring width chronology and its responses to climate change in Kuduer, Great Xing’an Mountains[J]. Chinese Journal of Applied Ecology, 2010, 21(10): 2501-2507.
[23]SANO M, FURUTA F, SWEDA T. Tree-ring-width chronology of Larix gmelinii as an indicator of changes in early summer temperature in east-central Kamchatka[J]. Journal of Forest Research, 2009, 14(3): 147-154.
[24]杨柳,李静茹,彭剑峰,等.1801年以来河南尧山地区油松高温变化及影响机制[J].生态学报,2021,41(1):79-91.
YANG L, LI J R, PENG J F, et al. Temperature variation and influence mechanism of Pinus tabulaeformis ring width recorded since 1801 at Yao Mountain, He’nan Province[J]. Acta Ecologica Sinica, 2021, 41(1): 79-91.
[25]WANG Z, LYU L X, LIU W X, et al. Topographic patterns of forest decline as detected from tree rings and NDVI[J]. CATENA, 2021, 198: 105011.
[26]陈峰,袁玉江,魏文寿,等.呼图壁河流域树轮年表的建立及其特征分析[J].沙漠与绿洲气象,2008,2(2):7-10.
CHEN F, YUAN Y J, WEI W S, et al. Development of the tree-ring chronology and its characteristic analysis in Hutubi river basin[J]. Desert and Oasis Meteorology, 2008, 2(2): 7-10.
[27]李陶,李明阳,钱春花,等. 结合冠层密度的森林净初级生产力遥感估测[J]. 南京林业大学学报(自然科学版), 2021, 45(5): 153-160.
LI Tao, LI Mingyang, QIAN Chunhua. Combining crown density to estimate forest net primary productivity by using remote sensing data[J].Journal of Nanjing Forestry University (Natural Science Edition), 2021, 45(5): 153-160.
[28]高景文,敖文军,刘顶锁,等.大兴安岭兴安落叶松的起源及生物特性[J].内蒙古科技与经济,2003,7(10):99-100.
GAO J W, AO W J, LIU D S, et al. Origin and biological characteristics of Larix gmelinii in Daxing’anling[J]. Inner Mongolia Science Technology and Economy, 2003, 7(10): 99-100.
[29]王秀伟,毛子军.兴安落叶松人工林冠层气体交换的时空特性[J].林业科学,2007,43(11):43-49.
WANG X W, MAO Z J. Temporal and spatial variation in gas exchange in canopy of Larix gmelinii plantation[J]. Scientia Silvae Sinicae, 2007, 43(11): 43-49.
[30]COULTHARD B L, TOUCHAN R, ANCHUKAITIS K J, et al. Tree growth and vegetation activity at the ecosystem-scale in the eastern Mediterranean[J]. Environmental Research Letters, 2017, 12(8): 084008.
[31]張扬,张秋良,李小梅,等.兴安落叶松林生长季碳交换对气候变化的响应[J].西部林业科学,2021,50(5):73-80,89.
ZHANG Y, ZHANG Q L, LI X M, et al. Response of carbon exchange to climate change in Larix gmelinii forest during growing season[J]. Journal of West China Forestry Science, 2021, 50(5): 73-80, 89.
[32]商志远,王建,崔明星,等.樟子松树轮δ13C的年内变化特征及其对气候要素的响应[J].植物生态学报,2012,36(12):1256-1267.
SHANG Z Y, WANG J, CUI M X, et al. Intra-annual variation in δ13C from tree rings of Pinus sylvestris var. mongolica and its response to climatic factors[J]. Chinese Journal of Plant Ecology, 2012, 36(12): 1256-1267.
