
三系杂交稻稻瘟病抗性与产量的配合力分析
2022-04-29 12:45李智谋姜守全于江辉管锋姚仁祥方杰
热带农业科学 2022年3期
李智谋 姜守全 于江辉 管锋 姚仁祥 方杰


摘要 為了阐明三系杂交稻稻瘟病抗性和产量之间一般配合力( GCA)、特殊配合力(SCA )的遗传效应,揭示二者间的关联性。本研究以6个三系野败型不育系和13个恢复系配制的78个杂交稻组合为材料,进行田间自然抗性鉴定,于稻瘟病盛发期调查各组合叶瘟、穗瘟、稻穗损失率,成熟期测定单株产量。配合力方差分析结果表明:稻瘟病抗性和单株产量均受亲本基因的加性效应和非加性效应的共同影响,但非加性效应高于加性效应,而且恢复系 GCA 基因型方差的贡献率大于不育系;稻瘟病抗性和单株产量受遗传变异效应影响较大; GCA 效应分析表明,不育系 L5A 和恢复系 LR7、 LR10均为正效应,不育系的穗瘟和穗瘟损失率之间为差异极显著正相关,恢复系稻瘟病3指标间为差异极显著正相关,单株产量与稻瘟病抗性间未达到显著相关性; SCA 的效应分析表明, L2A/LR1、 L3A/LR13、 L4A/LR10、 L4A/LR13、 L5A/LR4、 L5A/LR11、 L6A/LR1稻瘟病抗性指数和单株产量均为正效应,杂交稻组合稻瘟病3指标间为差异极显著正相关,而单株产量与稻瘟病抗性间为差异显著或极显著负相关。因此,抗病高产三系杂交水稻应选育 GCA 较高的亲本,同时注意协调组合的 SCA。
关键词 杂交水稻;配合力;遗传力;稻瘟病;单株产量
中图分类号 S511 文献标识码 A DOI:10.12008/j.issn.1009-2196.2022.03.006
Analysis of Combining Ability of the Rice Blast Resistance and Yield in Three-line Indica Hybrid Rice
LI Zhimou1 JIANG Shouquan1 YU Jianghui2 GUAN Feng1 YAO Renxiang1 FANG Jie1
(1. Hunan Hejiashan Seed Stock Station, Changde, Hunan 415123, China;2. Key Laboratory of Agro-ecological Processes in theSubtropical Region, Institute of Subtropical Agriculture, Chinese Academy of Sciences, Changsha, Hunan 410125, China)
Abstract In order to elucidate the genetic effects of parental GCA and F1 SCA on the rice blast and yield per plant of the indica hybrid rice, 78 combinations of indica hybrid rice from 6 new wild-abortive male sterile lines in the three-line system and 13 restorer lines were selected to identify their rice blast resistance in the field. The leaf blast, panicle blast, panicle blast loss rate were observed during the peak period of the rice blast, and the rice yield per plant was measured at the mature stage. The variance analysis of combining ability showed that the inheritance of the rice blast indexes and yield per plant were all affected by both the additive and non-additive effects of the parental genes and that the non-additive effect of the parental genes was higher than the non-additive effect of the parental genes. The general combining ability (GCA) variance of the re- storer lines was higher than that of the male sterile lines. The resistance to rice blast and the yield per plant were more affected by genetic variance. The GCA effect analysis showed that the sterile line L5A and the restorer lines LR7 and LR10 had posi- tive effect. The sterile line was highly significantly positively correlated with panicle blast and rate of panicle blast loss; the restorer lines had a highly significant correlation among the three indices of the rice blast; and the rice yield per plant was not significantly correlated with resistance to the rice blast. Specific combining ability (SCA) analysis showed the hybrid rice combinations L2A/LR1, L3A/LR13, L4A/LR10, L4A/LR13, L5A/LR4, L5A/LR11 and L6A/LR1 had positive SCA effects on the rice blast resistance indexes and the yield per plant. The leaf blast, panicle blast and rate of panicle blast loss were highly significantly negatively correlated in these combinations, and the yield per plant had a highly significantly negative correlation with the leaf blast, and a significantly negative correlation with the panicle blast or panicle blast loss rate. Therefore, parents with high GCA should be selected to develop indica hybrid rice with high disease resistance and yield in the three-line system, and the SCA of the crosses should also be considered.
Keywords hybrid rice; combining ability; heritability; rice blast; yield per plant
水稻( Oryza sativa L.)是世界上近一半人口的主粮,但水稻的种植和生长除干旱和洪涝灾害等非生物胁迫外,还受到由子囊菌Magnaporthe grisea(hebert)引起的稻瘟病侵害。稻瘟病是一种毁灭性真菌病害,其中叶瘟的症状最为典型,而穗瘟造成的损失最为严重[1]。目前,我国稻瘟病年发生面积在380万 hm2以上,由此造成的产量损失达10亿kg[2-3]。选育并推广抗病品种是稻瘟病防治最为经济有效的途径。基于标记基因型鉴定的分子标记辅助选择( marker assisted selec- tion, MAS )能大大提高水稻的稻瘟病抗性、选择准确性及育种效率,近年来培育出了大量抗病水稻新品系或新种质[4-12]。但多年来,育种家根据田间材料农艺性状及抗病虫性,通过常规育种方法选育出的品种占审定品种的极大比重,因此常规育种在我国种业的发展中依然具有重要地位。如杂交稻抗稻瘟病组合内6优595、沪优716、常优粳8号、川优712、佳谷优金占、泰优粤禾丝苗等,均为近3年内利用常规育种方法选育通过国家级或省级审定的抗稻瘟病杂交稻高产组合[13-18]。因此,育种家通过培育抗稻瘟病及具高产潜力的不育系或恢复系成为选育三系杂交稻抗病高产组合的重要方法。杂交稻组合杂种优势的发挥受亲本配合力的影响,分析杂交稻亲本配合力及其遗传规律对抗病高产育种具有重要价值。研究表明,通过分析不同品系或品种的配合力情况,可筛选出最优的杂交稻组合[19];通过遗传分析,可以確定出父、母本性状对杂种 F1性状的贡献率等[20-21]。为了进一步阐明杂交稻稻瘟病抗性和产量间的遗传和互作关系,本研究通过不完全双列杂交对78个杂交稻组合的稻瘟病和单株产量的一般配合力、特殊配合力和遗传力等进行分析,评价不育系和恢复系对杂交稻组合稻瘟病抗性和产量的贡献,为高产抗病杂交稻组合的选育提供参考。
1 材料与方法
1.1 材料
试验选用的6个三系不育系为 L1A~L6A,13个籼型恢复系为 LR1~LR13,所有材料均由湖南省贺家山原种场选育(表1)。2019年将6个不育系与13个恢复系按照 NCII 设计在贺家山原种场育种基地配制78个杂交稻组合。所有材料均根据历年来田间观察选育稻瘟病抗性优良且农艺性状较好单株加代繁殖直至稳定。
1.2 方法
1.2.1 试验设计于 2020年在湖南益阳桃江县稻瘟病抗性鉴定圃对78个配组的杂交稻组合进行稻瘟病田间自然抗性鉴定。试验采用随机区组设计,5月25日播种,6月20日移栽,株行距为16.5 cm ×26.4 cm,每个材料设3次重复,每次重复30穴。以 CO39作为稻瘟病诱发/感病对照品种,全生育期不施用杀菌剂,肥水管理按一般高产栽培管理。于稻瘟病盛发期调查各材料叶瘟、穗瘟、稻穗发病率指数,每个重复调查30株。成熟期每个小区除边行外各选取10穴单株混合收种求取平均单株产量。
1.2.2 病情调查及数据分析病情调查和分级标准参照《水稻品种试验稻瘟病抗性鉴定与评价技术规程》(NY/T 2646—2014)、《水稻品种(系)抗稻瘟病鉴定方法与抗性评价技技术规程》(DB 32/T 1123—2007)。用 DPS v14.1软件对数据进行配合力参数计算,并进一步计算各亲本的一般配合力,组合的特殊配合力,配合力采用遗传方差、广义遗传力和狭义遗传力参数。
2 结果与分析
2.1 稻瘟病抗性指标和单株产量的配合力方差分析
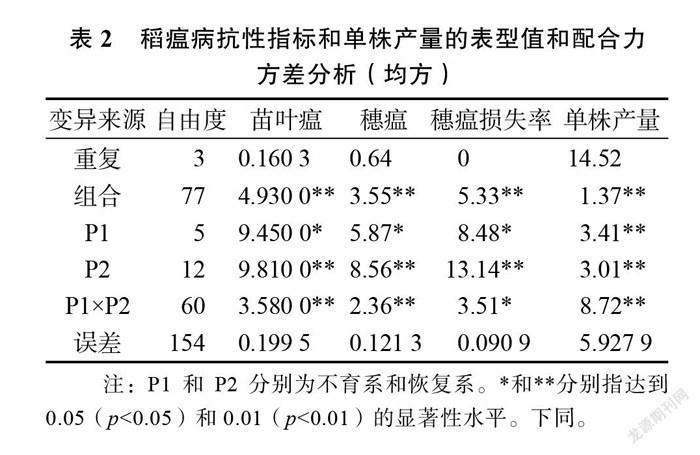
由配合力方差分析结果(表2)表明,供试组合的稻瘟病抗性3指标(叶瘟、穗瘟、稻穗损失率)和单株产量在重复间均未达到差异显著水平,说明这些表型比较稳定;而稻瘟病抗性指标和单株产量均方均达到了差异极显著水平,说明不同的杂交稻组合稻瘟病抗性和单株产量在基因型间存在明显的遗传差异;不育系( P1)的稻瘟病3项指标一般配合力( GCA)均达到了差异显著水平,恢复系( P2)的稻瘟病3 项指标 GCA 和杂交稻组合的特殊配合力( SCA )均达到了差异极显著水平,亲本单株产量的 GCA 和组合的 SCA 均为差异极显著水平,表明稻瘟病各项指标和单株产量性状的遗传均受基因的加性效应和非加性效应的影响。因此,可进一步分析亲本的 GCA 和组合的 SCA。
2.2 群体遗传参数分析
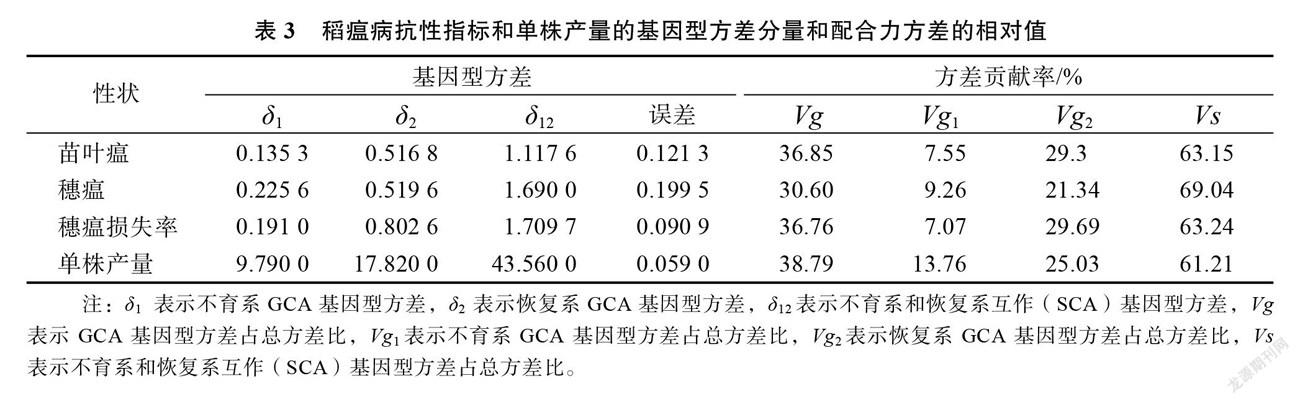
不育系和恢复系的 GCA 方差为加性方差,而组合的 SCA 方差为非加性方差。表3可知,检测的4项性状中亲本(δ1、δ2) GCA 基因型方差均小于亲本间互作(δ12)SCA 基因型方差,而且 SCA 基因方差 Vs 贡献率在稻瘟病和单株产量所占比重均大于60%,即亲本基因的非加性效应对后代性状贡献率超过60%,同时亲本的基因加性效应 GCA 贡献率超过30%,表明稻瘟病和单株产量遗传基因为非加性效应起主导作用,而基因的加性效应也起重要作用,因此育种上应特别注重组合 SCA 的筛选,亦要考虑亲本的 GCA。而且,穗瘟 SCA 基因方差最大,表明穗瘟受基因的非加性效应最高,苗叶瘟和穗瘟损失率 SCA 基因方差贡献率差异不大。
分析不育系、恢复系对杂交组合稻瘟病各指标和单株产量的方差贡献率可以看出,在 GCA 基因型方差( Vg)中不育系和恢复系所占的比重不同。研究结果表明,在测试的4项性状中,恢复系( Vg2)GCA基因型方差的贡献率比不育系( Vg1)都大,表明恢复系对杂种 F1稻瘟病抗性作用和单株产量的贡献均高于不育系,但苗叶瘟和穗瘟损失率不育系( Vg1) GCA 基因型方差的贡献率比较接近,也占到7%以上,单株产量不育系( Vg1) GCA 基因型方差的贡献率占到13.76%,为4项指标最高。因此,育种中亲本 GCA 的考虑主要注重恢复系的选择,同时兼顾不育系(表3)。
杂交稻组合稻瘟病和单株产量的遗传力分析结果(表4)表明,广义遗传率均在90%以上,表明遗传变异效应大,且单株产量的广义遗传率达到了99.92%;而狭义遗传率为单株产量>穗瘟损失率病级>苗叶瘟>穗瘟,变幅为28.28%~38.76%,说明穗瘟受环境影响相对较大,在后代遗传中的稳定性较差,由不育系、恢复系直接传递给杂种后代的能力较差,应考虑环境和组合的 SCA 影响。
2.3 亲本稻瘟病各指标和单株产量的一般配合力效应分析
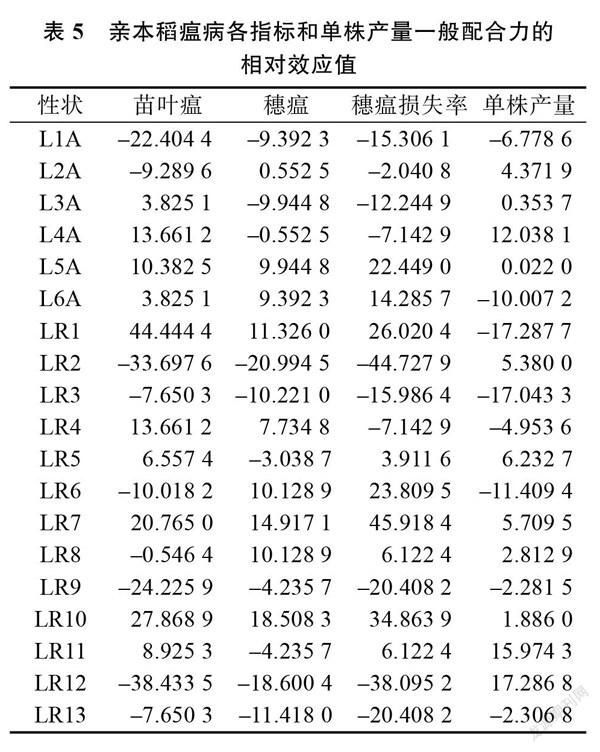
各亲本的 GCA 效应分析结果见表5。研究表明,不同不育系或恢复系的稻瘟病各指标间或单株产量 GCA 效应存在明显差异,且表现为不同的正负效应,说明亲本稻瘟病各指标间和单株产量的加性效应值存在明显差异。研究表明, 6个不育系中 L3A~L6A 苗叶瘟表现为正效应, L2A 、 L5A、 L6A 穗瘟表现为正效应, L5A、 L6A 穗瘟损失率表现为正效应, L2A~L5A 单株产量表现为正效应,因此, L5A 为各指标均为正效应的不育系;13个恢复系中 LR1、LR4、LR5、LR7、LR10、 LR11苗叶瘟表现为正效应, LR1、 LR2、 LR3、 LR4、LR6~LR8、LR10穗瘟表现为正效应, LR1、 LR5~LR8、 LR10、 LR11穗瘟损失率表现为正效应, LR2、 LR5、 LR7、 LR8、 LR10~LR12单株产量表现为正效应,因此, LR7、 LR10为各指标均为正效应的恢复系。综上所述,不育系 L5A 和恢复系 LR7、 LR10有较好的应用前景。
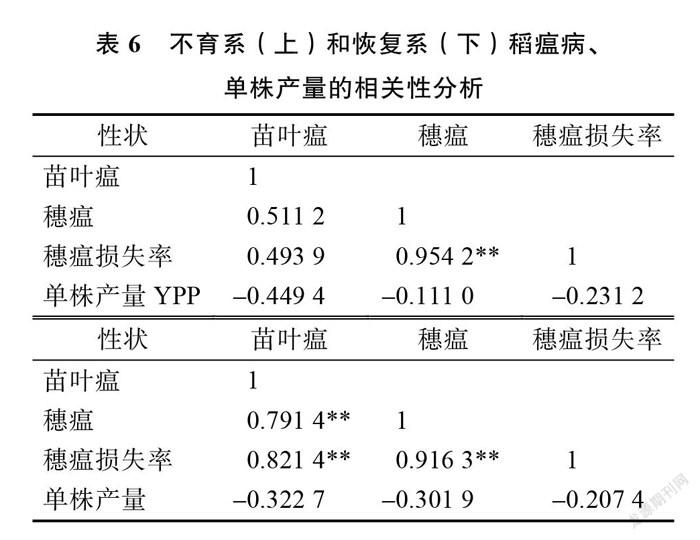
为了进一步明确稻瘟病3项指标与单株产量间亲本 GCA 的关联性,对不育系和恢复系4项测试指标的 GCA 进行相关性分析。结果(表6)表明,不育系的穗瘟和穗瘟损失率之间 GCA 为差异极显著正相关,恢复系的苗叶瘟、穗瘟、穗瘟损失率之间 GCA 为差异极显著正相关,不育系和恢复系的单株产量与稻瘟病3指标间无差异显著相关性。因此,在亲本抗稻瘟病的 GCA 选择时,不育系可以注重穗瘟或穗瘟损失率选择其一、恢复系可以选择苗叶瘟、穗瘟、穗瘟损失率任何1个指标为参考。
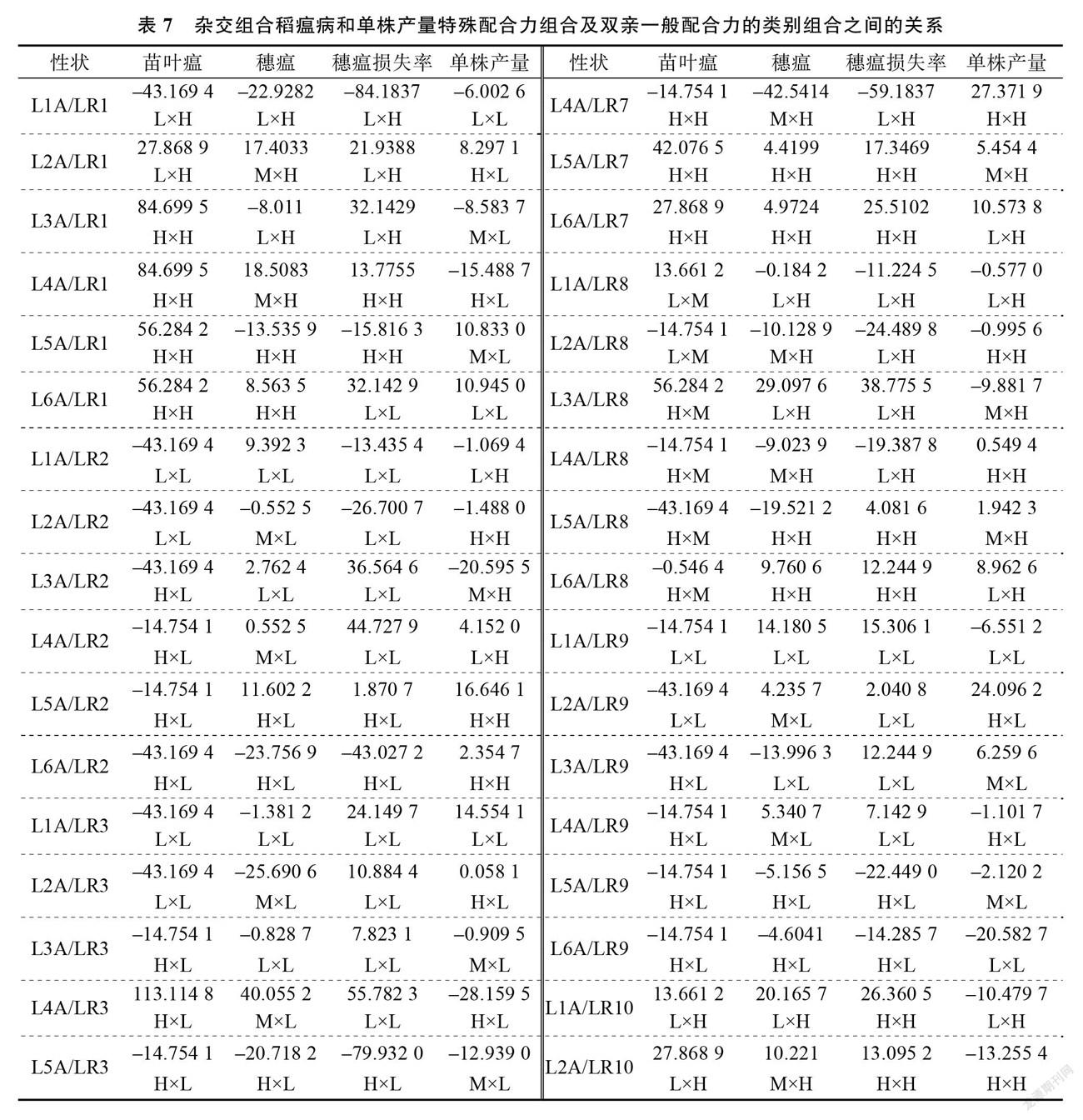
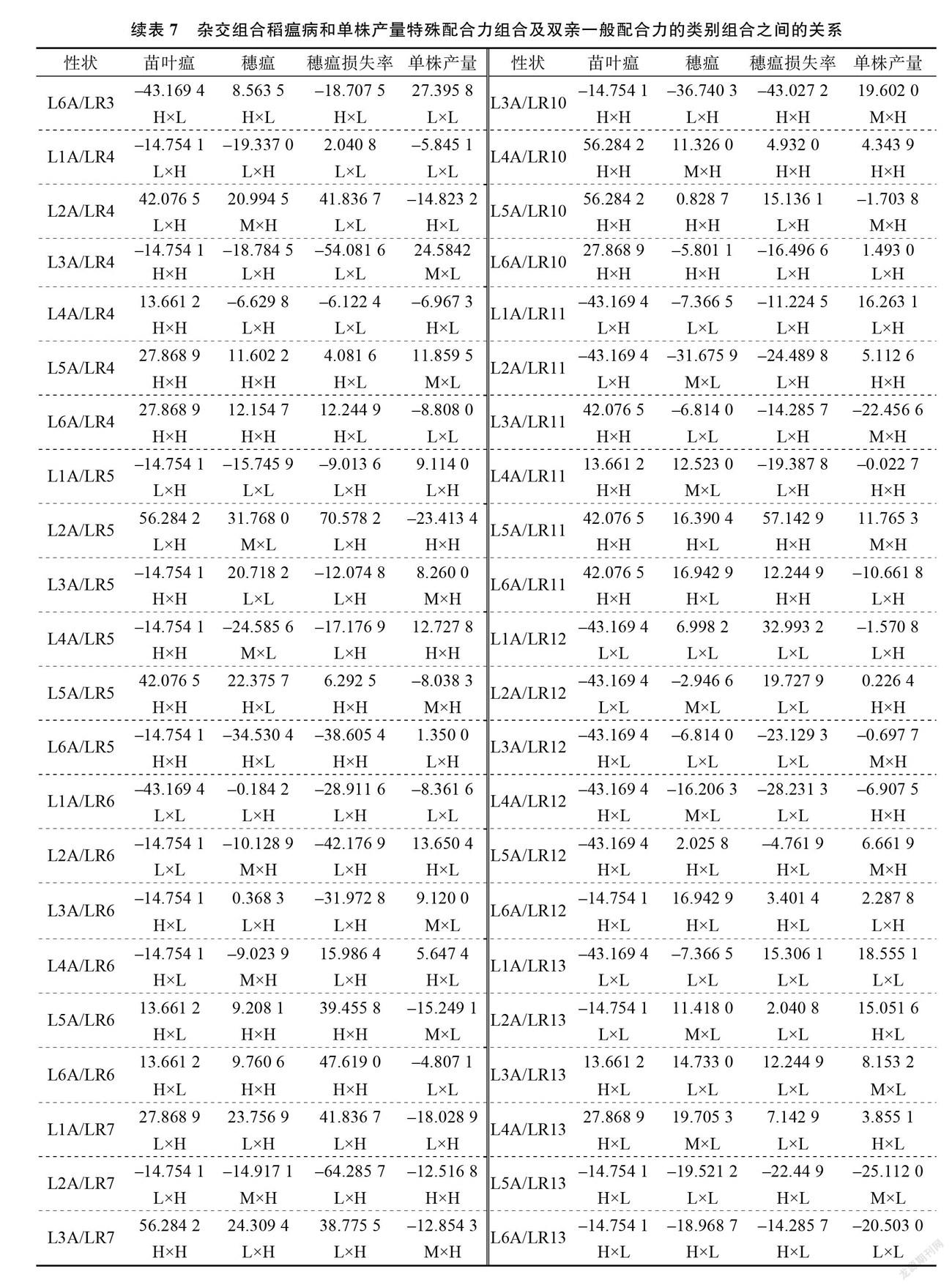
2.4 特殊配合力的效应分析及和亲本一般配合力的关系
SCA 效应反映杂交组合非加性效应的大小,可以为优良抗稻瘟病且高产组合的选育提供理论依据。由表7可知, SCA 的效应值在不同组合、稻瘟病不同指标间、单株产量上均存在显著差异,且同一组合的不同指标、同一亲本的不同组合所表现出的 SCA 也有所不同。在供试的78个杂交稻组合中, L4A/LR3苗叶瘟、穗瘟的 SCA 效应值最高,穗瘟损失率的 SCA 效应值也较高(第三)。表明组合 L4A/LR3稻瘟病抗性受亲本基因的非加性效应最具优势。 L6A/LR3、 L4A/LR7单株产量的 SCA 效应值位列前2。此外, L2A/LR1、L3A/LR13、 L4A/LR10、 L4A/LR13、 L5A/LR4、 L5A/LR11、 L6A/LR1稻瘟病各指标和单株产量 SCA 均为正效应。
为了进一步明确双亲 GCA 和组合 SCA 的关系,将不育系和恢复系 GCA 的效应值分为高( H)、中( M)、低( L)3种类型,以 GCA 效应值>1为高、在–1~1为中、<–1为低。从双亲的 GCA 来看,叶瘟的 SCA 和不育系、恢复系的关联较大,在78个杂交稻组合中,叶瘟的 SCA 效应值为正值的组合有31个,其中母本、父本 GCA 为 H×H 的组合有18个,占到58.1%,表明父母本的 GCA 的效应值为高( H)配制出 SCA 较高的组合概率大些。穗瘟的 SCA 效应值为正值的组合有39个,母本、父本 GCA 为 H ×H 的组合9个, M×L 的组合8个, H×L 的组合7个, M×H 的组合6个, L×L 的组合6个, L ×H 的组合3个,表明穗瘟受基因的加性效应无规律可循。穗瘟损失率的 SCA效应值为正值的组合有44个,母本、父本 GCA 为 L×L 的组合19个,占到43.2%,双亲 H×H 的组合13个,占到29.5%,可见父母本 GCA 为 L×L 型、 H ×H 型配制出 SCA 较高的组合概率大些。单株产量的 SCA 效应值为正值的组合有39个,其中恢复系为 H 型的组合的有22个,占到56.4%,但在 SCA 效应值为负值的39个组合中,恢复系为 H 型的组合的有20个,占到51.3%,表明单株产量的 SCA 的效应值与恢复系的 GCA 无关。综上所述,三系杂交稻组合稻瘟病各项指标的非加性效应受亲本 GCA 的影响比较复杂。
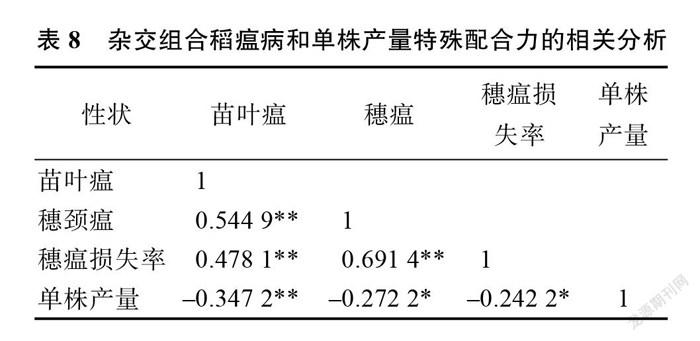
为了进一步研究杂交稻组合稻瘟病各指标和单株产量 SCA 之间关联性的大小,对稻瘟病3指标、单株产量的 SCA 进行相关分析。结果表明,叶瘟、穗瘟、穗瘟损失率之间 SCA 为差异极显著正相关,而单株产量与叶瘟间 SCA 为差异极显著负相关,与穗瘟、穗瘟损失率间为差异显著负相关(表8)。表明杂交稻组合稻瘟病抗性的 SCA 高,单株产量反而会降低。
3 讨论与结论
3.1 讨论
研究结果表明,三系杂交稻组合稻瘟病抗性和单株产量受杂种 F1 SCA 和亲本 GCA 的共同作用,其中杂种 F1 SCA 的遗传贡献超过60%,说明稻瘟病抗性和单株产量主要受亲本基因的非加性效应影响,而亲本 GCA 的遗传贡献也在30%以上;在基因的加性效应中,恢复系对杂种 F1稻瘟病的抗性作用和单株产量贡献高于不育系,印证了前人就杂交水稻单株产量方面的报道。刘金波等[22]通过不完全双列杂交( NCII )模式对28个杂交稻组合主要农艺性状进行了配合力分析,结果表明,单株产量的 GCA 和 SCA 均达到差异极显著水平,且单株产量主要受组合 SCA 的影响,父本 GCA 大于母本 GCA 的作用;王丽等[23]以引进的5个不育系和自育的6个恢复系进行不完全双列杂交,研究认为单株产量受基因非加性效应影响大,恢复系对后代的影响大于不育系。而沈村义等[2,4]、刘龙钦等[25]研究认为,单株产量性状的遗传主要受亲本加性效应控制,李双等[26]、潘清洁等[27]研究认为,单株产量受不育系影响较大。前人关于单株产量等农艺性状的遗传率进行了大量的分析,但研究结果不同或有差异。本研究结果表明,单株产量的广义遗传率达到了99.92%,即遗传变异效应和环境影响较大。據此推测,试验结果除与供试材料有较大关系外,还和试验的环境条件、气候因素等关系较大。本研究结果表明,单株产量 SCA 与叶瘟、穗瘟、穗瘟损失率的 SCA 间为差异显著或极显著负相关,不育系和恢复系单株产量的 GCA与稻瘟病3指标间无显著相关性,但恢复系对4项测试指标的遗传贡献均高于不育系,因此抗病高产杂交稻组合亲本的选育对恢复系的选择更为关键,组合的选育要注意稻瘟病抗性和产量间的协调,不能一味追求高产或高抗。
杂交水稻强优势组合的选育关键在于亲本选择,而亲本的正确组配关键在于亲本配合力分析。研究认为,杂交稻组合单株产量的 SCA 与亲本的 GCA关联不大[22-23,26];本研究结果与此结论一致,结果表明,杂种 F1的单株产量和穗瘟 SCA 与不育系、恢复系的GCA 效应值关系较复杂,无规律可循。而父母本叶瘟 GCA 为 H ×H 型的材料容易配制出 SCA 较高的杂交稻组合,父母本为 L ×L 及 H ×H 型组合的穗瘟损失率容易配制出 SCA 较广的杂交稻组合。而且本研究发现,不育系的穗瘟和穗瘟损失率的 GCA 为差异极显著正相关,恢复系稻瘟病3指标间为差异极显著正相关。因此,抗病组合的选育稻瘟病3指标均应选择 GCA较高的亲本即 H ×H 型组合,对应杂交稻组合稻瘟病 SCA 的效应高的概率大些。同时,尽管前人研究认为,杂交稻组合单株产量的 SCA 和亲本 GCA 之间没有明显的对应关系,但研究认为, GCA 较高的亲本更容易配出性状优良的杂种后代,组合间的 SCA 效应值高低也是决定后代杂种优势强弱的重要指标[28],为了提高育种效率,育种工作者倾向于在 GCA 高的亲本基础上组配 SCA 高的杂交组合[22, 28]。为此,三系杂交水稻单株产量和稻瘟病抗性应选亲本 GCA 较高的材料。
3.2 结论
本文对78个籼型杂交稻组合稻瘟病和单株产量的 GCA、SCA 和遗传率等进行研究,结果表明,稻瘟病各项指标和单株产量性状的遗传均受基因的加性效应和非加性效应的共同影响,不育系 L5A 和恢复系 LR7、LR10为 GCA 较好的材料,恢复系 GCA 基因型方差的贡献率大于不育系,不育系的穗瘟和穗瘟损失率之间 GCA 为差异极显著正相关,恢复系的叶瘟、穗瘟、穗瘟损失率之间 GCA 为差异极显著正相关,而单株产量与稻瘟病3指标间无差异显著相关性。杂交稻组合叶瘟、穗瘟、穗瘟损失率之间 SCA 为差异极显著正相关,而单株产量与稻瘟病3指标为差异显著或极显著负相关。因此,抗稻瘟病高产三系杂交水稻应选育 GCA 较高的亲本,同时注意协调组合的 SCA。
参考文献
[1] 沈浙南, 时焕斌, 邱结华, 等.江苏省212份水稻材料中抗稻瘟病基因 Pi2/Pi9/Piz-t/Pigm的情况分析[J/OL].分子植物育种:1-11[2022-03-03]. http://kns.cnki.net/kcms/detail/46.1068.S.20210413.1020.002.html.
[2] 邓云.2017-2019年福建省稻瘟病菌生理小种组成与致病性[J].福建农林大学学报(自然科学版), 2021, 50(4):451-459.
[3] 王伟舵,于俊杰, 聂亚锋, 等.2011—2011年江苏省稻瘟病菌种群动态及毒力变化[J].江苏农业学报, 2015, 31(2):285−289.
[4] 倪大虎, 易成新, 李莉, 等.利用分子标记辅助选择聚合水稻基因 Xa21和 Pi9(t)[J].分子植物育种 , 2005(3):329-334.
[5] 殷得所, 夏明元, 李进波, 等.抗稻瘟病基因 Pi9的 STS连锁标记开发及在分子标记辅助育种中的应用[J].中国水稻科学, 2011, 25(1):25-30.
[6] Jiang H C, Feng Y T, Bao L, et al. Improving blast resistanceof Jin 23B and its hybrid rice by marker-assisted gene pyra- miding[J]. Molecular Breeding, 2012, 30(4):1679-1688.
[7] 曹志, 曾盖, 郝明, 等.利用 MAS 技术改良水稻两用核不育系 C815S 的稻瘟病抗性[J].分子植物育种, 2015, 13(6):1193-1200.
[8] 杨丰宇 , 李永聪 , 刘雄伦 , 等. 分子标记辅助选择改良早籼稻1701的稻瘟病抗性[J].分子植物育种, 2017, 15(6):2212-2217.
[9] 张礼霞, 王林友, 范宏环, 等.利用Pigm基因改良粳稻保持系的稻瘟病抗性研究[J].核农学报, 2017, 31(3):424-431.
[10] Chen Q H, Zeng G, Hao M, et al. Improvement of rice blastand brown planthopper resistance of PTGMS line C815S in two-line hybrid rice through marker-assisted selection[J].Molecular Breeding, 2020, 40(2):1-13.
[11] 李永聪, 黄俊, 廖花, 等.分子标记辅助选择改良水稻恢复系 R389及其杂交种稻瘟病抗性[J].华北农学报, 2018, 33(2):112-118.
[12] 刘树芳, 董丽英, 赵国珍, 等.抗稻瘟病基因 Piz-t 和 Pi9连锁标记开发及在云南粳稻中的应用[J].西南农业学报, 2016, 29(4):721-725.
[13] 盧东柏, 刘维, 陆展华, 等.抗稻瘟病三系杂交水稻新组合泰优粤禾丝苗[J].杂交水稻, 2021, 36(4):121-123.
[14] 赵鹏, 郑灵, 沈久淑, 等.抗稻瘟病香稻恢复系内恢10595的选育与利用[J].杂交水稻, 2021, 36(4):31-32.
[15] 黄成志, 雷树凡, 吕直文, 等.抗稻瘟病优质三系杂交水稻新组合沪优716[J].杂交水稻, 2021, 3(5):1-3.
[16] 陈惠清, 王天生, 谢少和, 等.抗稻瘟病优质籼型三系不育系佳谷 A 的選育[J].杂交水稻, 2021, 36(2):30-32.
[17] 王小虎, 潘斌清, 兰国防, 等.优质抗稻瘟病杂交粳稻新组合常优粳8号的选育[J].中国种业, 2021(6):99-101.
[18] 袁小珍, 杨波, 刘洁, 等.优质抗稻瘟三系杂交水稻新组合川优712[J].杂交水稻, 2021, 36(5):1-3.
[19] 唐文帮, 肖应辉, 邓化冰, 等.水稻两用核不育系 C815S的配合力及利用价值[J].湖南农业大学学报(自然科学版), 2010, 36(4):367-372.
[20] 余守武, 尹建华, 刘宜柏, 等.三交水稻的育种研究Ⅲ.三交中晚稻主要农艺性状的配合力和遗传力分析[J].作物学报, 2005, 31(6):784-789.
[21] 宋宇, 邹小云, 贺浩华, 等.籼型三系杂交水稻产量及其相关性状的配合力分析[J].江西农业大学学报 , 2004, 26(5):719-725.
[22] 刘金波, 宋兆强, 王宝祥, 等.几个新选杂交水稻不育系和恢复系的主要农艺性状的配合力分析[J].西南农业学报, 2016, 29(2):209-213.
[23] 王丽, 赵德明, 林纲, 等.杂交稻新组合主要农艺性状配合力分析及利用评价[J].江西农业大学学报, 2012, 34(2):208-212+219.
[24] 沈村义, 蔡开炯, 蒋素, 等.杂交水稻主要经济性状配合力的研究[J].湖南农业科学, 2016(9):17-20.
[25] 刘龙钦, 江文清, 梁康迳, 等.若干籼型杂交稻亲本的农艺性状配合力分析[J].福建稻麦科技, 2018, 36(1):1-6.
[26] 李双, 唐显岩, 王春雷, 等.8个籼型恢复系相关农艺性状的配合力分析[J].植物遗传资源学报, 2019, 20(5):1 178-1185.
[27] 潘清洁, 赵福胜, 罗洪发, 等.5个新选水稻三系不育系主要农艺性状配合力分析[J].南方农业学报 , 2020, 51(1):36-41.
[28] 肖长春, 程祖锌, 黄昕颖, 等.功能型红米恢复系主要农艺性状的配合力和遗传力分析[J].热带作物学报, 2021, 42(7):1876-1884.
(责任编辑 龙娅丽)
猜你喜欢
农民致富之友(2020年15期)2020-05-25
江苏农业学报(2019年1期)2019-09-10
农民致富之友(2019年24期)2019-08-20
农民致富之友(2019年5期)2019-03-13
农民致富之友(2018年20期)2018-11-25
农家科技下旬刊(2017年1期)2017-04-08
南方农业·下旬(2016年8期)2017-02-23
湖南师范大学学报·自然科学版(2015年4期)2016-03-01
安徽农学通报(2015年20期)2015-11-07
安徽农学通报(2015年10期)2015-06-15
