
贵州火龙果的气孔特征及其日变化规律
2022-05-06 00:29聂克艳李莉婕王小红张贵进赵泽英
贵州农业科学 2022年4期
聂克艳, 李莉婕, 王 苓, 王小红, 张贵进, 赵泽英*
(1.贵州省农业科技信息研究所, 贵州 贵阳 550006; 2.安顺学院, 贵州 安顺 561000)
0 引言
【研究意义】贵州火龙果(Hylocereusundatus)主要在南北盘江流域和红水河流域的罗甸、关岭和望谟等区域种植,是当地农业产业发展的支柱产业之一[1]。紫红龙是贵州火龙果主栽品种,所处的山区生态环境复杂多样,土层浅薄保水难,多为季节性干旱的干热河谷和低海拔的石漠化地区[1]。气孔作为植物与外界环境进行水分和气体交换的重要通道,在某种程度上间接反应了植物对生长环境水分含量变化的适应。气孔在控制植物体内水分损失和生物量积累之间的平衡中起着关键作用,气孔开度和导度直接影响植物蒸腾。研究火龙果的气孔特征及其日变化规律,对探究贵州田间水分管理方法和促进火龙果高效生产具有重要意义。【前人研究进展】朱玉等[2]对3种北高丛蓝莓气孔特征研究表明,高温条件下蓝莓主要通过增加气孔的宽度和规则化气孔的空间分布提高光合效率。孙志蓉等[3]对环草石斛和铁皮石斛不同生长发育时期试管苗叶片的气孔进行扫描电镜观察发现,随着生长时间的延长,环草石斛和铁皮石斛叶片气孔的大小变化不大,而气孔的数量和密度均增加。美花石斛在水分充足和干旱处理后,其气孔开放率表现一致,均是夜间开放率远大于中午开放率,表现出专性植物特征[4]。张延龙等[5]对起源于我国的葡萄属(Vitis)20个种或变种叶片气孔特性进行观察发现,气孔纵径对葡萄属种的分类有较大的价值。PAZOUREK[6]研究发现,鸢尾属的叶片气孔密度随着光强度的减少而增加。通常情况下,同种植物单个叶片的气孔密度排序为叶基>叶中>叶尖[7]。董天英等[8]研究发现,向日葵等10种常见植物叶片不同部位的气孔密度无明显规律性。邹龙海[9]研究铁皮石斛的景天酸代谢途径与其关键基因的共表达网络结果表明,铁皮石斛成熟叶片的光合代谢为CAM途径。苏文华[10]等对不同天气条件下铁皮石斛叶片CO2吸收速率的一昼夜波动研究发现,晴天铁皮石斛夜晚和白天的CO2吸收速率均表现为CAM途径;而在阴雨天则表现为C3途径;多云天气下,白天和夜晚均吸收CO2,植株光合代谢途径在C3和CAM途径之间转换,说明铁皮石斛的光合代谢途径随着外界环境条件的变化在C3和CAM途径之间变换。【研究切入点】火龙果肉质茎作为蒸腾作用的主要器官,分布着大量气孔,直接控制气孔蒸腾强度。火龙果肉质茎气孔性状及其影响因素在很大程度上可有效促进其在作物生产上的调控,但到目前为止,少见火龙果肉质茎气孔的相关研究。【拟解决的关键问题】通过电镜观察紫红龙火龙果的肉质茎和果实表皮气孔,分析各部位气孔特征和气孔日变化规律,以期为火龙果气孔研究、树体水分调控和田间水分管理提供科学支撑。
1 材料与方法
1.1 材料
供试火龙果品种为紫红龙(贵州省果树科学研究所选育,黔审果2009005号),种植于贵州数字农业技术研究基地(惠水县)大棚,为5年生成年树。选择长势相对一致、育龄大体相当的1年生肉质茎和2年生肉质茎标记备用。
1.2 方法
1.2.1 肉质茎和果实表皮的显微结构观察 参照李莉婕[1]的扫描电镜观察方法,用刀片将肉质茎上部表皮切成长宽厚分别为4 mm×4 mm×1 mm的小方块,将其放在10 mL离心管中,用7 mL 2.5%戊二醛4℃固定12 h以上;用pH 7.0的磷酸缓冲液漂洗4次后,用浓度分别为30%、50%、60%、70%、80%、90%、100%乙醇梯度脱水各15~20 min,然后用乙酸异戊醋置换,最后用100%乙醇浸泡过夜。用CO2临界点干燥仪对样品进行干燥,用离子溅射仪镀金,在S-3400型扫描电子显微镜(日本)下进行扫描观察,加速电压模式,10.0 kV,分辨率4.0 nm,放大100~1 000倍。
1.2.2 肉质茎和果实表皮的气孔密度和大小测量 用300倍照片计算气孔密度,用5 000倍照片测量气孔大小和开张程度,每个处理选取10个视野,取平均值,用Photoshop进行测量。用形状指数S(通过计算单一气孔形状与相同面积的圆之间的偏离程度)测量气孔形状的复杂程度,即当气孔为圆形时,其形状指数为1;若气孔的形状越扁长,则气孔形状指数的值就越大。
式中,S为形状指数,P为气孔开口周长(mm),A为气孔开口面积(mm2)。
1.2.3 植物气孔特征测量 植物气孔特征主要表现在气孔器大小、气孔开口大小以及形状上。通过对火龙果1年生肉质茎、2年生肉质茎和果实表皮的气孔进行观察,选择气孔器的长度、宽度、面积、周长、气孔比和气孔内开口长度、宽度、面积、周长、形状指数等10个指标表述火龙果各部位气孔特征。通过电镜观察记录连续24 h肉质茎和果实的气孔开闭情况。气孔导度用气孔计测定。
1.3 数据处理
数据用Excel和SPSS统计软件进行整理。
2 结果与分析
2.1 肉质茎和果实表皮的显微结构
从图1看出,2年生肉质茎表皮的显微结构表面角质层纹路清晰,围绕气孔器呈拼图状镶嵌分布,沟壑分明,无显著纵横向发育特征,花纹较小,纹路较为平坦松散,与气孔器的形态差异明显。1年生肉质茎表皮的显微结构,表面角质层花纹的形态分布与2年生肉质茎大致相同,但1年生肉质茎纹路较为饱满立体,条纹之间孔隙较小,结构较为紧缩,紧密贴合于气孔器周围。果实表皮的显微结构,花纹与肉质茎有明显差异,其纹路较深,花纹较大,呈明显纵向发育,褶皱较多,具有乳突状突起,与向下凹陷的椭圆形气孔器差异较明显。
2.2 肉质茎和果实表皮的气孔密度和空间格局
气孔大小和气孔密度能够反映植物的生态学特性。火龙果肉质茎气孔器分布于茎的3个面,呈散生型分布,镶嵌于凹凸不平的表皮之上,气孔两侧的肾形保卫细胞环绕形成气孔口,2个副卫细胞环绕于肾形保卫细胞周围,与不规则的表皮细胞形状差异较大;茎表皮气孔开放时,保卫细胞内缘向内凹陷形成开口;茎气孔关闭时,气孔两侧保卫细胞内缘呈“一”字型紧密贴合。
从图2看出,2年生肉质茎的气孔器为近圆形,其保卫细胞与周围表皮细胞之间的沟壑较浅。1年生肉质茎的气孔器也为近圆形,其保卫细胞形状较为饱满,与周围表皮细胞的区别较明显。与肉质茎气孔器形状截然不同,火龙果果实表皮气孔器呈长椭圆形,不规则分布于果皮表面,多为向下凹陷,保卫细胞呈长条状,外缘较平滑;副卫细胞呈月牙形围绕在保卫细胞两侧,少数气孔器不见明显副卫细胞,与周围乳突状表皮细胞相比具有较大差异;果实表皮气孔开口较大,开放时,气孔由中间向椭圆形两端逐渐打开,关闭时,气孔保卫细胞呈平行状收缩合拢。火龙果2年生肉质茎的表皮气孔密度(30.07个/视野)略高于1年生肉质茎气孔密度(28.07个/视野),均极显著高于果实表皮的气孔密度(11.80个/视野)(图3)。
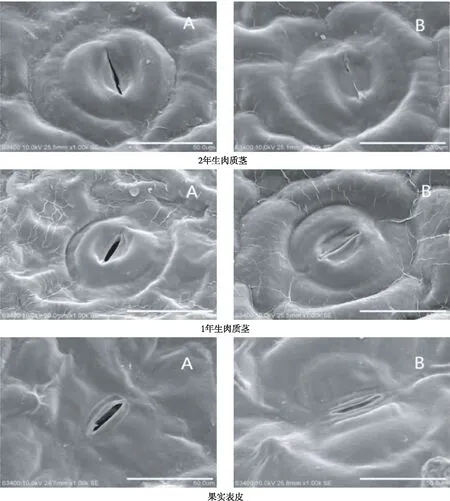
注:A为气孔开放状态,B为气孔关闭状态。
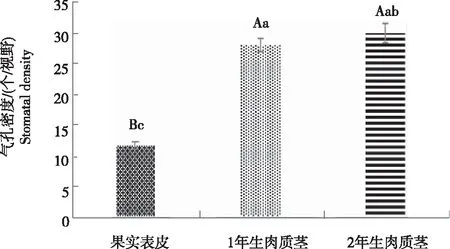
注:不同大、小写字母分别表示差异极显著(P<0.01)和显著(P<0.05)。
2.3 不同部位的气孔开放情况
从图4看出,24 h内,2年生肉质茎气孔开放率最高在4:00和18:00,分别达47.92%和50.00%;其次在8:00和10:00,分别为37.50%和33.04%;在0:00、2:00、6:00、12:00、14:00和20:00时,均未看到视野中有气孔开放。1年生肉质茎气孔开放率最高在4:00和16:00,分别达36.11%和24.29%,其次在18:00和20:00,分别为21.43%和23.81%;在2:00、10:00、12:00和14:00时,均未看到视野中有气孔开放。果实表皮气孔开放率最高在4:00和16:00,此2个时间点气孔开放率均达100%,其次在6:00和18:00,分别为92.31%和94.44%;气孔开放率最低在2:00和14:00,分别为75.91%和77.50%。

图4 连续24 h内火龙果肉质茎和果实气孔的平均开放率
2.4 肉质茎气孔导度的日变化规律
从图5看出,2年生肉质茎24 h内在0:00-5:00气孔导度逐渐增大至118.5 mmol/(m2·s),5:00-8:00逐渐降低至30.1 mmol/(m2·s),8:00-22:00呈上下波动变化;23:00时气孔导度最低,为24.3 mmol/(m2·s);23:30时增至最高,为129.7 mmol/m2·s)。1年生肉质茎在0:00-3:00气孔导度逐渐增大,3:00-6:00剧烈波动,在4:00达最高值140.4 mmol/(m2·s),6:00-23:30平缓波动,在10:30达最低值15.3 mmol/(m2·s)。1年生和2年生肉质茎变化趋势基本一致,2年生肉质茎气孔导度总体高于1年生肉质茎,且变化趋势更加明显。在7:00—17:00时气孔导度较低,在2:00—6:30以及18:00—至23:30时气孔导度较高。
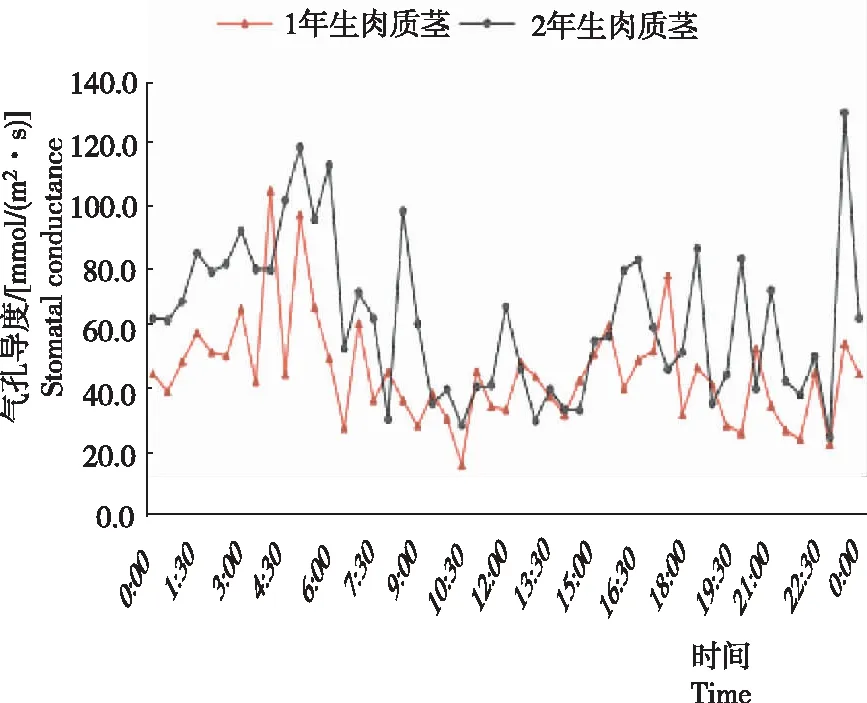
图5 火龙果肉质茎24 h内的的气孔导度
2.5 不同部位气孔特征及日变化规律
2.5.1 气孔器和气孔内开口特征 从表1看出,火龙果不同部位气孔大小以及形状均有所不同。1年生和2年生肉质茎气孔器的长、宽、面积和周长大小较相近,两者间差异不显著,但均显著高于果实表皮气孔器。1年生和2年生肉质茎气孔内开口的宽度显著低于果实表皮,而1年生肉质茎的气孔内开口宽度略大于2年生肉质茎,说明果实气孔内开口的张开程度比肉质茎大。火龙果各部位气孔内开口的面积和形状指数均表现出显著性差异,气孔内开口的长度和周长无显著性差异。在形状上,2年生肉质茎和果实的形状指数偏高,而1年生肉质茎的形状指数更接近于1,即1年生肉质茎的形状更近似于圆形。
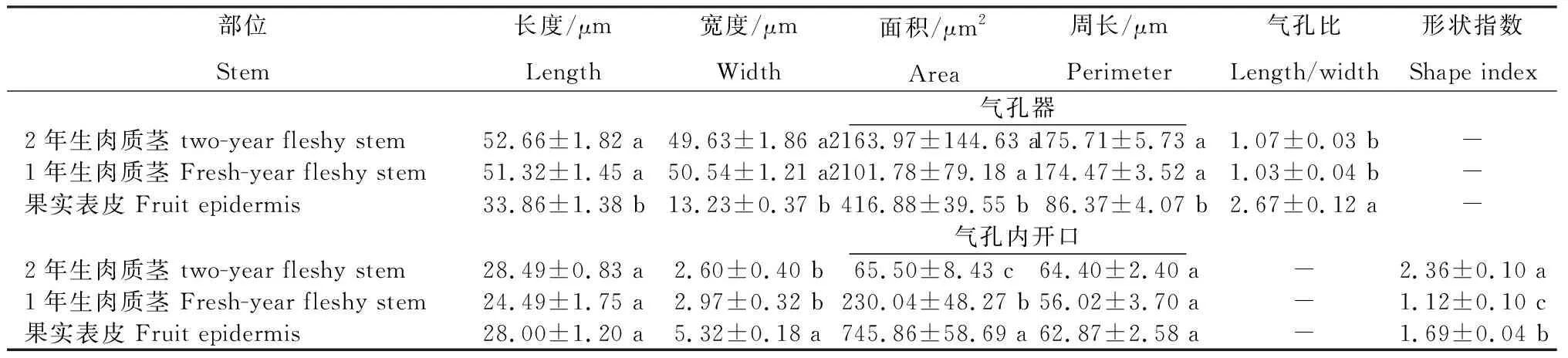
表1 火龙果不同部位的气孔器和气孔内开口特征
2.5.2 气孔开度的日变化规律 从图6看出,2年生肉质茎气孔开度(气孔内开口横径与纵径之积)在14:00-18:00、20:00-22:00、2:00-4:00、6:00-10:00呈升高趋势,10:00时达最高,为115.24 μm2;在12:00-14:00、0:00-2:00气孔处于关闭状态;在18:00-20:00、22:00-0:00、4:00~6:00呈下降趋势,并最终下降至气孔完全完闭。1年生肉质茎气孔开度在2:00-4:00、14:00-16:00、18:00-20:00呈升高趋势,在4:00时达最高,为124.59 μm2;在16:00-18:00、20:00-2:00、4:00-10:00呈不同幅度下降趋势;在2:00、10:00、12:00-14:00处于完全关闭状态。火龙果果实气孔在24 h中均呈不同程度的开放状态,在14:00-18:00、20:00-22:00,0:00-2:00、4:00-8:00气孔开度呈升高趋势,在18:00时达最高,为174.90 μm2,在12:00-14:00、18:00-20:00、22:00-0:00、2:00-4:00、8:00-10:00呈下降趋势,在0:00时最低,为87.37 μm2。

图6 火龙果各部位24 h内的气孔开度
2.6 气孔性状特征的相关性
从表2~4看出,火龙果各部位气孔不同性状特征之间存在不同程度的相关性。2年生肉质茎气孔开度与气孔内开口横径、面积、周长、形状指数及气孔开放率均呈极显著正相关,其中与气孔内开口横径的相关性最高;肉质茎形状指数与气孔内开口横径、面积、周长、气孔开度、气孔开放率均呈极显著正相关,其中与气孔内开口周长的相关性最高(表2)。1年生肉质茎气孔开度与气孔内开口横径、面积、周长及气孔开放率均呈极显著正相关,其中与气孔内开口横径相关性最高;肉质茎形状指数与气孔内开口横径、周长、气孔开放率均呈极显著正相关,其中与气孔内开口周长的相关性最高(表3)。果实气孔开度与气孔器面积、气孔内开口横径、面积及周长均呈极显著正相关,其中与气孔内开口横径的相关性最高;果实气孔的形状指数与气孔器横径、气孔内开口横径呈极显著负相关,与气孔比呈极显著正相关(表4)。表明,不同部位气孔特征指标的相关性具有差异,气孔内开口横径是影响气孔各特征指标变化的重要因素。
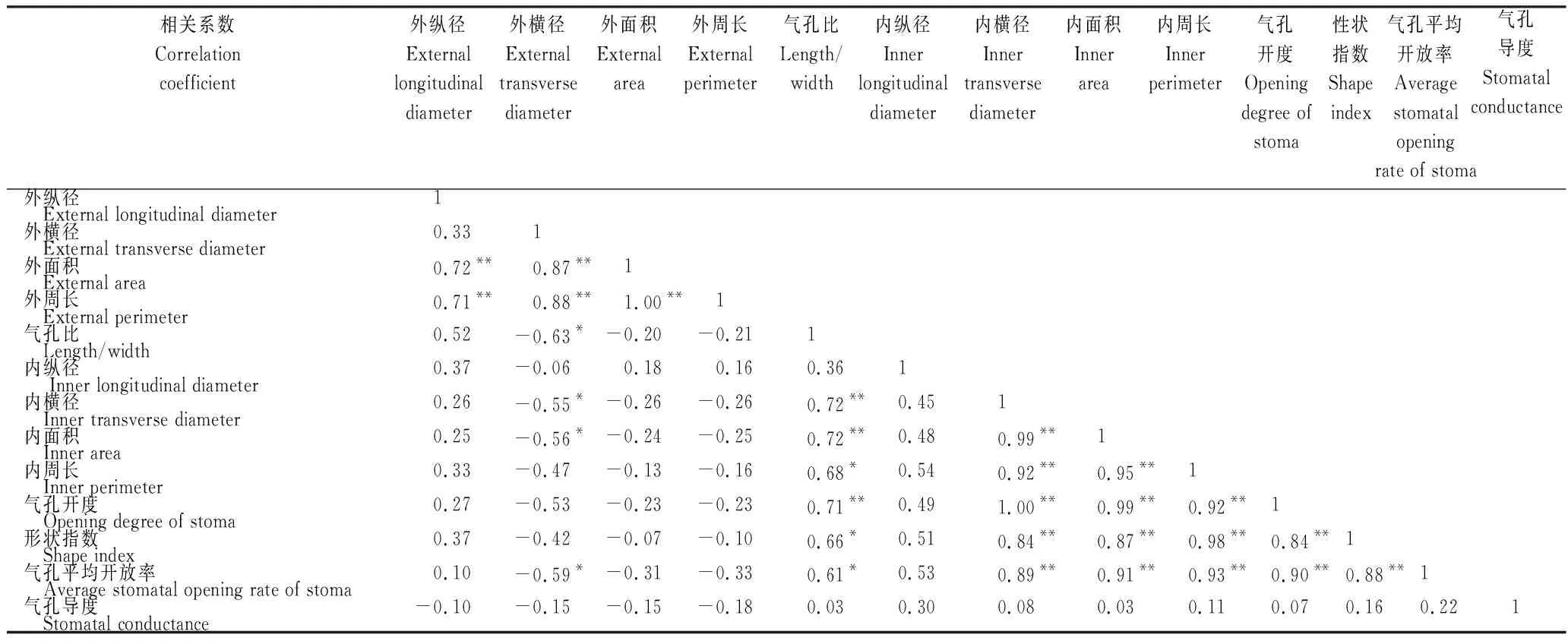
表2 2 年生肉质茎气孔特征指标的相关系数

表3 1年生肉质茎气孔特征指标的相关系数
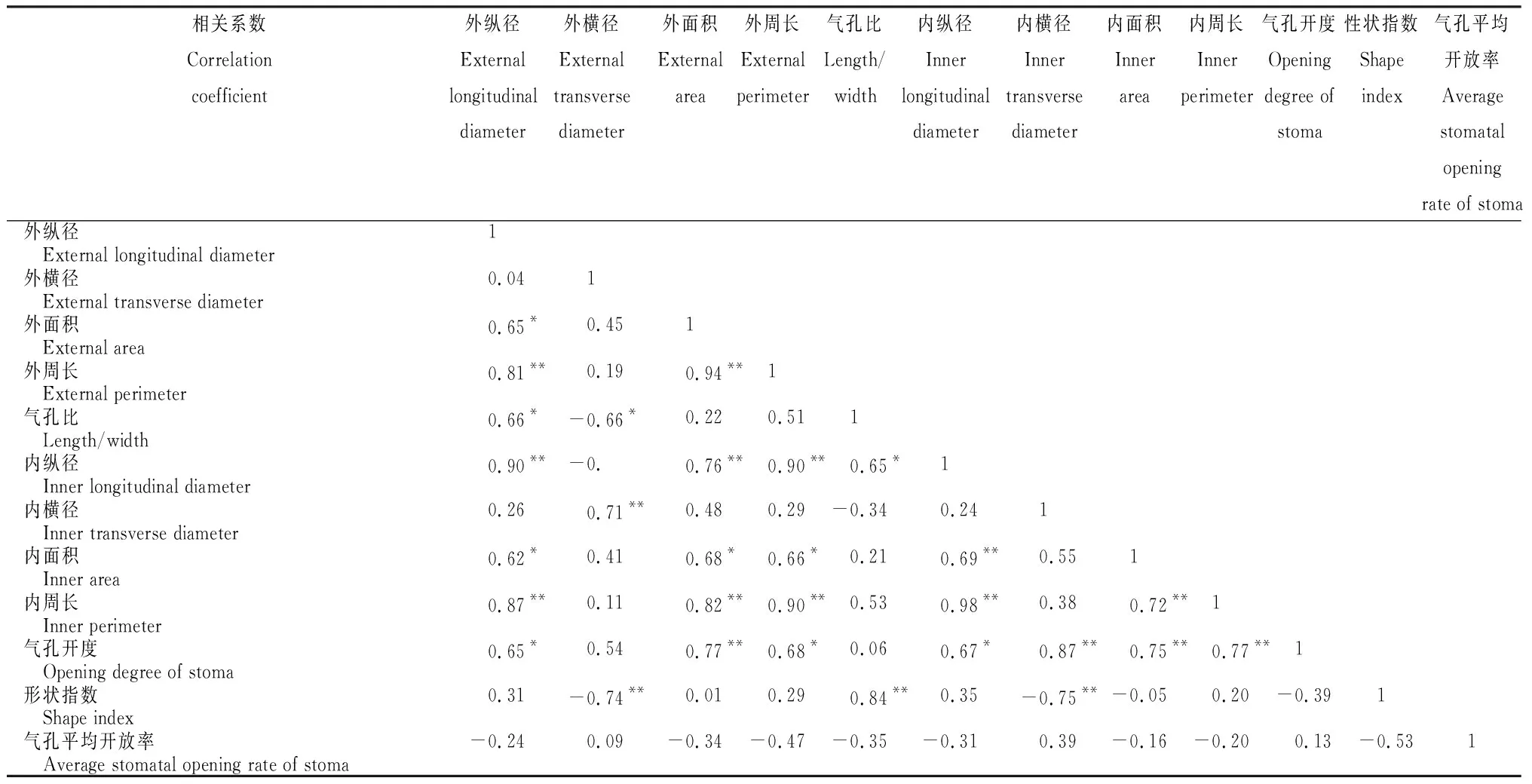
表4 果实气孔特征指标的相关系数
3 讨论
气孔是植物与外界环境进行气体交换的重要通道[11],其形态特征会因温湿度、土壤水含量和光照等外界因素发生变化[12-13],但气孔整体特征由遗传因素决定。研究发现,火龙果果实表皮与肉质茎表皮气孔形态特征差别较大;果实表皮褶皱较多,具有明显乳突状突起,与向下凹陷的肉质茎近圆形气孔器差异较明显。火龙果肉质茎气孔呈下陷状态,下陷的气孔在减少光线辐射和风沙损害方面有积极的作用[14],一方面受遗传因素控制,另一方则可能是对生长环境的一种适应机制[15-16]。此外,气孔形态特征还因肉质茎的生长发育时间不同而存在一定差异。与2年生肉质茎相比,1年生肉质茎纹路较为饱满立体,条纹之间孔隙较小,结构较为紧缩,紧密贴合于气孔器周围,这种差异可能是1年生肉质茎表皮细胞尚未发育成熟所致[17]。
火龙果肉质茎气孔器镶嵌于凹凸不平的表皮之上,肉质茎的气孔器呈近圆形,保卫细胞饱满膨胀,且不因肉质茎的生长发育时间而发生明显改变。火龙果果实表皮气孔器呈长椭圆形,保卫细胞呈长条状,与肉质茎近圆形的气孔器区别较为明显。相同放大倍数视野下,2年生肉质茎表皮气孔密度最大,约为果实表皮气孔密度的2.55倍,可见火龙果的气孔器主要分布在成熟的肉质茎中。气孔蒸腾是植物蒸腾作用的最主要方式,是植物吸收和运输水分的主要动力,火龙果的蒸腾部位主要在肉质茎。植物气孔特征表现主要与土壤含水量相关,气孔开度对抗旱性的影响最大[18],而对于抗旱系数直接作用影响最大的是气孔密度[12]。为使植物水分利用效率最大化,应采取有效措施来调控田间的生态因子以调控气孔密度,抑制蒸腾速率,避免产生植物“午休”现象,从而增强树体的光合作用。
CAM途径代谢的植物有专性和兼性之分,叶片年龄和外界条件变化会导致其光合碳同化途径在C3和CAM间转换[19]。遗传因素、个体发育及环境因素如光强[20]、相对湿度和水分都可以在一定程度上从生理生化方面影响CAM植物的表达。研究发现,火龙果在气孔的呼吸作用中表现出CAM植物特有的规律,火龙果各部位气孔主要在夜间开放,且在夜间气孔开度较大,但在白天也见果实气孔开放,而2年生肉质茎和1年生肉质茎在白天无烈日暴晒时可见气孔开放。火龙果肉质茎气孔导度在7:00-17:00时较低,在2:00-6:30及18:00-23:30时气孔导度较高。综上说明,火龙果肉质茎气孔的呼吸作用主要在夜晚进行,白天的呼吸作用较弱。与成熟叶片相比,自然条件下幼叶因其位于冠层顶部,更容易暴露在强光和高温下,而强光和高温能够直接或间接影响气孔开度和气孔导度[20]。火龙果新生肉质茎在一定程度上为成熟肉质茎遮挡了强光,从而也直接或间接地影响其气孔的开度和气孔导度。火龙果1年生肉质茎的气孔导度普遍低于2年生肉质茎,在气孔大小上,2年生肉质茎也表现出比1年生肉质茎更大的特征,显然气孔导度的高低主要受气孔发育的影响,与冬青卫矛和华北紫丁香的气孔上研究结果[21]一致。因此,叶片生长过程中气孔导度的提高主要取决于气孔大小,这一结果也表明,未发育成熟的1年生肉质茎气孔可能具有较弱的气体交换功能。
植物表面的微观形态结构是研究物种起源、进化和分类的重要指标。相关研究已在荔枝、龙眼、柑橘、苹果和胡杨等植物中广泛应用[22],气孔与各物种的分类学和种间亲缘有着密切联系,且有研究认为,气孔密度、大小的变异系数和重复力与遗传变异关系密切[23]。相关研究表明,蝴蝶兰叶片的气孔面积与气孔开度呈显著正相关,说明植物进行光合、呼吸和蒸腾作用等生理活动时,叶片上需要一定数量的气孔面积;气孔开度在一定程度上决定了气孔面积。同时,蝴蝶兰叶片一些气孔指标间具有较高的相关性[24]。研究表明,火龙果不同部位气孔性状指标之间的相关性具有较大差异。肉质茎的气孔特征指标中,气孔内开口横径和气孔开度及形状指数之间都具有较高的相关性,不仅如此,其他指标之间也表现出不同程度的相关性。因此,在气孔研究过程中,要考虑到性状之间的相互影响,在选择气孔指标时,应该关注这些性状彼此间的相关性并加以充分利用。
4 结论
火龙果气孔主要分布在成熟的肉质茎上,2年生肉质茎的气孔密度大于1年生肉质茎。火龙果肉质茎表皮气孔呈近圆形,保卫细胞饱满膨胀,显微结构图纹路较为平坦;果实表皮气孔呈椭圆形,保卫细胞呈长条状,具有明显乳突状突起;火龙果气孔开闭情况和常见CAM植物相似,气孔开放集中在清晨和傍晚时分。火龙果气孔导度整体呈波浪式变化规律,但在0:00和5:00火龙果气孔导度逐渐增大至峰值,日间降低,傍晚时分又有所升高。火龙果各部位气孔开度整体呈升-降-升-降的规律变化,且均在14:00和00:00出现增加趋势。火龙果气孔特征各指标间存在不同程度的相关性,肉质茎中气孔开度与气孔内开口横径、面积、周长、气孔开放率等均呈极显著正相关,气孔内开口横径是影响气孔各特征指标变化的重要因素。
猜你喜欢
今日农业(2022年2期)2022-11-16
航空材料学报(2022年4期)2022-08-04
美食(2022年4期)2022-04-16
干旱地区农业研究(2022年1期)2022-01-28
科学与财富(2021年33期)2021-05-10
美食(2020年5期)2020-06-01
山东工业技术(2019年9期)2019-05-29
科技创新与品牌(2019年12期)2019-02-06
戏剧之家(2018年21期)2018-10-19
魅力中国(2016年31期)2017-06-15
