
基于具缘纹孔膜特征的木质部栓塞机制研究进展*
2022-05-11 11:54李玉军万贤崇姜笑梅孙熙佑朱新京殷亚方
林业科学 2022年2期
李 姗 李玉军 张 亚 王 杰 万贤崇 姜笑梅 孙熙佑 朱新京 殷亚方
(1.中国林业科学研究院木材工业研究所 北京 100091; 2.中国林业科学研究院木材标本馆 北京 100091; 3.国家林业和草原局木材科学与技术重点实验室 北京 100091; 4.陕西科技大学环境科学与工程学院 西安 710021; 5.西北工业大学机电学院 西安 710072; 6.安徽师范大学生命科学学院 芜湖 241000; 7.中国林业科学研究院林业新技术研究所 北京 100091)
在全球陆地生态系统中,植物约贡献45%的碳汇资源,对保障水文和营养循环、维持生物多样性起着至关重要的作用(Panetal., 2011); 但随着气候变化、干旱、虫害和冻害胁迫导致的植物木质部栓塞频发(申卫军等, 1999; Clarketal., 2016; 李志民等, 2019),原本充水的木质部管道被空气占据,植物面临着输水功能障碍、分生组织活力降低、初级生产力减小、死亡率增加的风险(Chenetal., 2019; Choatetal., 2018; Lengetal., 2013; Lietal., 2017; 2021),且一些已经发生栓塞的植物复水之后面临着更高的栓塞形成风险(栓塞疲劳)(Hackeetal., 2001; Stilleretal., 2002),其木质部抗栓塞能力降低。因此,明确木质部栓塞成因,厘清木质部栓塞形成、扩散和疲劳机制,对提高植物抗寒、抗旱能力具有重要的科学价值。
大多数被子植物导管和裸子植物管胞中普遍存在的具缘纹孔,可精准调控植物水分传导效率,应对气穴栓塞(Tyreeetal., 2002; 万贤崇等, 2007),其中具缘纹孔膜是木质部栓塞形成、扩散和疲劳产生的关键部位(Jansenetal., 2018)。近年来,具缘纹孔膜特征在木质部栓塞机制研究中日益受到关注。为更全面和清晰理解木质部栓塞形成、扩散和疲劳过程中具缘纹孔膜的作用,本研究在整理分析国内外木质部栓塞相关研究的基础上,首先探讨被子植物、裸子植物木质部栓塞的形成机制和扩散规律,阐释栓塞形成和扩散的“气种假说”,明确具缘纹孔膜的核心作用; 然后分别针对具缘纹孔膜的构造、化学、物理和微力学性质与微液流行为等主要特征以及不同特征间的相互关系,归纳具缘纹孔膜在木质部栓塞形成、扩散和疲劳等不同阶段的重要作用,概述三维结构模型构建、化学成分原位检测、力学行为模拟和单个纹孔膜微液流量化分析等具缘纹孔膜特征关键表征方法的发展现状和问题; 最后提出木质部栓塞应优先开展的研究方向。
1 木质部栓塞的形成机制和扩散规律
关于木质部栓塞成因,迄今为止被广泛接受和认可的最主要机制为“气种假说”(air seeding hypothesis)(Tyreeetal., 2002),其中具缘纹孔的纹孔膜起核心作用(图1)。具缘纹孔主要存在于被子植物导管和裸子植物管胞中(IAWA Committee, 1989; 2004),是木质部输水管道侧壁次生加厚产生的穹窿形空洞,将输水管道连接成一个三维网络。与之不同的另一种纹孔类型是单纹孔,纹孔呈圆柱形,缺乏拱形环抱的纹孔缘,纹孔膜一般不具纹孔塞或纹孔托,常出现在韧性纤维、轴向薄壁细胞和射线薄壁细胞中(IAWA Committee, 1989; 姜笑梅等, 2010)。具缘纹孔膜由构成输水管道的相邻木质部细胞复合胞间层(含初生壁)发育而成,为水分在相邻管道间的流动提供了重要通道。
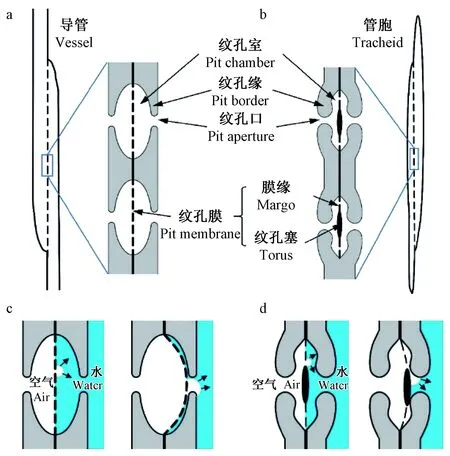
图1 被子植物导管与裸子植物管胞的具缘纹孔结构及“气种假说”
根据“气种假说”,当木质部汁液负压超出输水管道中水分的抗张强度时,连续的水柱断裂形成空腔,来自外界或植物组织自身的微小气泡在具缘纹孔膜两侧压力差作用下迅速进入原本充水的相邻管道,不断膨胀并填充空腔,最终导致充水管道内形成栓塞。木质部栓塞形成后,气泡经由已栓塞输水管道的具缘纹孔膜在相邻输水管道间传播,即发生栓塞扩散。此过程中,具缘纹孔膜位置由纹孔室中央向一侧逐渐偏移,甚至紧贴纹孔口导致纹孔闭塞(Sperryetal., 2004)。同时,具缘纹孔膜基于自身结构和力学性质对膜两侧压力差的响应发生变形甚至损伤(Hillabrandetal., 2016),微液流效率随之改变。在木质部栓塞后复水时,所引发的压力差变化使具缘纹孔膜回弹至纹孔室中央,当再次经历栓塞时又发生纹孔膜向纹孔口偏移,其反复偏移导致自身构造特征、力学性质和微液流效率发生显著变化,继而可能导致木质部在较低压力差阈值下形成栓塞,即栓塞疲劳(Umebayashietal., 2019)。持续栓塞会引起气泡经由具缘纹孔膜不断扩散、输水系统大面积栓塞,最终导致木质部水力失衡,植物即使复水也难以恢复活力,甚至发生死亡(Lietal., 2015)。
过去20年中,学者们多从组织水平研究不同树种木质部栓塞形成和扩散规律,并常采用木质部栓塞抗性作为评价栓塞形成、扩散和疲劳难易程度的关键指标(Lietal., 2016; Zhangetal., 2018)。以被子植物为例,葡萄(Vitisvinifera)树栓塞首先出现在髓心周围的初生木质部导管内,而后径向扩散至树皮(Brodersenetal., 2013); 小果黑核桃(Juglansmicrocarpa)栓塞则首先出现在散生导管,随后扩散至临近导管团(Knipferetal., 2015)。就裸子植物而言,北美红杉(Sequoiasempervirens)主干木质部栓塞在管胞间,同时以弦向带状扩散、散生管胞扩散以及与叶柄木质部相连通的管孔团扩散3种方式扩散(Choatetal., 2015); 而花旗松(Pseudotsugamenziesii)栓塞首先出现在髓心周围的初生木质部和靠近树皮的管胞中,并逐步向木质部中部扩散(Torres-Ruizetal., 2016)。木质部输水管道连通性以及具缘纹孔膜构造、性质和微液流行为的差异,可能是导致不同树种栓塞形成和扩散规律多样化的主要因素。近年来,基于木质部输水管道连通性开展木质部栓塞机制的研究相对较多,但受限于精准表征手段的局限性,从具缘纹孔膜角度探究栓塞形成和扩散机制的研究仅限于生物仿真模拟,如Park等(2019)采用纳米纤维素纸模拟栓塞经由被子植物具缘纹孔膜形成和扩散的方式,提出栓塞扩散呈现持续型和离散型2种方式,并进一步证实了“气种假说”。
此外,以往研究也提出栓塞形成的其他机制或假说,如气泡的同型成核机制(homogeneous nucleation)、异型成核机制(heterogeneous nucleation)以及纳米气泡假说(nanobubble hypothesis)等,这些机制或假说对栓塞形成和扩散可能同时起辅助作用(Pickard, 1981; Schenketal., 2015); 但是,目前绝大多数研究结果仍支持“气种假说”作为栓塞形成和扩散的主要机制,认为明确具缘纹孔膜构造特征、性质和微液流行为对理解木质部栓塞形成、扩散和疲劳机制至关重要(Torres-Ruizetal., 2016; Jansenetal., 2018)。
2 具缘纹孔膜主要特征及其在木质部栓塞中的作用
具缘纹孔膜特征主要包含其三维构造、化学、物理和微力学性质与微液流行为,这些特征相互关联、相互影响,在木质部栓塞形成、扩散和疲劳等阶段均起着重要作用,共同决定木质部栓塞抗性和疲劳性(图2,Lietal., 2020)。

图2 具缘纹孔膜主要特征及其在木质部栓塞中的作用示意
2.1 具缘纹孔膜构造特征
明确具缘纹孔膜构造特征,是深入理解木质部栓塞形成和扩散机制的重要基础。根据Young-Laplace方程,引起木质部栓塞扩散的相邻导管间压力差ΔP与纹孔膜上微孔半径R呈反比(Schenketal., 2015)。近年来提出的纳米气泡假说认为,当体积较大的气泡从已栓塞导管通过由无数个纹孔膜微孔组成的通道时,如果每个有效孔隙通道的最小孔隙半径越小,则大气泡被分解成纳米气泡(nanobubble)的概率越大。虽然这些纳米气泡可通过具缘纹孔膜进入相邻充水导管,但不会再次聚集形成栓塞(Schenketal., 2015)。同时,随着具缘纹孔膜厚度增加,纹孔膜具有小孔径通道的概率也提高,因此当具缘纹孔膜微孔直径越小且纹孔膜越厚时,纳米气泡聚集并导致栓塞发生的概率越小,木质部栓塞抗性越强(Kaacketal., 2019; Lietal., 2016)。由此可见,具缘纹孔膜厚度、孔隙结构在木质部栓塞形成和扩散中发挥着决定性作用。
实现具缘纹孔膜构造特征的精准表征,是探究木质部栓塞形成、扩散和疲劳机制的前提条件。关于具缘纹孔膜构造特征的研究始于20世纪初,当时的观测局限在光学显微镜水平(Bailey, 1913; Phillips, 1933)。20世纪中叶出现的碳复型法结合透射电子显微镜技术,呈现了具缘纹孔膜的亚显微构造(Frey-Wysslingetal., 1956; Ctéetal., 1964)。随后,各国学者结合透射电子显微镜、场发射扫描电子显微镜、冷冻扫描电子显微镜等多种手段研究了不同树种具缘纹孔膜的超微构造及其生物学功能(周崟等, 1990; 姜笑梅等, 1992; 1996; 殷亚方, 2002; Jansenetal., 2009; Gorbetal., 2010; Lietal., 2019; 2021; Wangetal., 2021)。样品保存状态对具缘纹孔膜观测结果影响较大,相较于离体干燥的木材样品,采自活立木新鲜样品更有利于具缘纹孔膜微观结构的精准表征(Lietal., 2016; Zhangetal., 2020)。研究表明,被子植物与裸子植物的具缘纹孔膜构造特征差异明显,被子植物的具缘纹孔膜大多具有均质性,由多层随机分布的纤维素微纤丝堆叠排列,形成大量形状不规则的微孔,孔隙直径在5~20 nm之间(Kaacketal., 2019),这些微孔既是相邻导管间的水分流动通道,也是栓塞在相邻导管间的扩散通道。此外,也有部分被子植物的具缘纹孔膜呈现3层膜、2层膜和有塞膜的特异型结构,其各层间纤维素微纤丝排布方式不一(Jansenetal., 2004; Sano, 2005)。而裸子植物的具缘纹孔膜具有异质性,通常由中部加厚的纹孔塞和高度孔隙化的膜缘组成,膜缘微纤丝束随机组合呈网状排列(周崟等, 1994; 佟永萍等, 2007),孔隙直径约为0.1 μm(Hackeetal., 2004),是其木质部水分流动的安全阀门(Roskillyetal., 2019)。受样品处理方式和表征手段的影响,目前具缘纹孔膜高分辨率成像仍存在技术瓶颈,不同手段获取的结果差异较大(Zhangetal., 2020),且相关研究多集中在二维尺度,纳米精度水平上具缘纹孔膜的三维空间结构特征尚不明晰。
近年来,随着成像技术的快速发展,针对木质部细胞壁及其具缘纹孔膜构造特征的研究不断深入(Guoetal., 2016; Zhangetal., 2017),越来越多学者逐渐聚焦具缘纹孔膜三维结构的可视化及模型构建(Mascheketal., 2013; Zhangetal., 2020);但由于具缘纹孔膜上微孔的空间结构紧密相连且迂曲度复杂(Kaacketal., 2019),现有手段如X射线CT叠层成像(PXCT)、4Pi联用激光共聚焦显微镜等依然普遍存在成像分辨率不足等问题,具缘纹孔膜三维结构模型构建的相关研究还处于起步阶段(Schulte, 2012; Schulteetal., 2015)。最近,有研究采用虚拟沉积技术(Lietal., 2018)对被子植物具缘纹孔膜进行了三维模型构建(Lietal., 2020),该模型可呈现纤维素微纤丝的随机堆叠,更接近木质部具缘纹孔膜的真实形貌。
2.2 具缘纹孔膜化学和物理性质
在木质部栓塞形成、扩散和疲劳过程中,具缘纹孔膜的三维结构发生明显变化(Hackeetal., 2001; Tixieretal., 2014),这些变化与具缘纹孔膜自身化学性质密切相关(Hillabrandetal., 2016; Zhangetal., 2017),具缘纹孔膜的化学成分直接影响木质部栓塞形成和扩散规律(Pereiraetal., 2018a)。研究表明,枝条纤维素水解显著降低木质部栓塞抗性(Dusotoit-Coucaudetal., 2014),木质素含量与木质部栓塞抗性呈正相关(Pereiraetal., 2018b)。总体而言,木质部具缘纹孔膜的主要化学成分为纤维素,其被包埋在由木质素、半纤维素和果胶组成的无定形基质中(Schmitzetal., 2012),但不同植物木质部具缘纹孔膜的化学成分和含量不尽相同。被子植物成熟木质部具缘纹孔膜的主要化学成分为纤维素,还含有木质素(Herbetteetal., 2014)以及少量的酚类化合物和蛋白质等物质(Pereiraetal., 2018a),果胶和半纤维素在其发育过程中水解消失,仅纹孔环具有微量果胶(Kaacketal., 2019; Plavcovetal., 2011)。裸子植物木质部具缘纹孔膜一般含有木质素(Kimetal., 2014),且其纹孔塞与膜缘之间的化学成分差异显著,其中纹孔塞主要成分为果胶(Danieletal., 1996),而膜缘则主要为纤维素。尽管紫外分光光度法、X射线光谱显微镜、免疫标记、荧光标记、近红外纳米光谱技术等多种手段被用于具缘纹孔膜化学成分检测(Boyceetal., 2004; Schmitzetal., 2008; Mascheketal., 2013; Kimetal., 2016; Pereiraetal., 2018a),但遗憾的是,受限于技术手段和分辨率,目前还难以实现新鲜饱水状态下具缘纹孔膜化学成分的原位精准表征。
在木质部栓塞不同阶段,具缘纹孔膜所处水分环境发生明显变化。由于纤维素微纤丝具有亲水性,水分子存在影响细胞壁中聚合物分子链之间的孔隙分布状态(Yinetal., 2015; 2017; Hanetal., 2020),从而改变其原有结构(Kulasinskietal., 2015),因此,水分环境变化直接影响具缘纹孔膜三维结构,导致其表现出干缩湿胀行为,进而引起其微液流效率发生显著变化。尽管研究发现离体干燥状态下具缘纹孔膜厚度收缩近一半,孔隙直径也显著减小(Zhangetal., 2020),但其纤维素微纤丝的微观结构对不同水分环境的响应机制仍需要进一步探究。
2.3 具缘纹孔膜力学性质
在木质部栓塞形成过程中,具缘纹孔膜两侧的压力差发生明显变化,导致其力学行为随之改变。研究发现,冻融交替引发的色木槭(Acermono)木质部栓塞疲劳性具有明显的季节性变化规律,具体表现为夏季栓塞疲劳性最大、冬季栓塞疲劳性最小,这可能与具缘纹孔膜力学性质的季节性差异有关(Zhangetal., 2017)。作为一种具各向异性的生物质材料,具缘纹孔膜在外力作用下表现出非线性的力学变形行为,随着外界压力增大,纹孔膜逐渐经历弹性、塑性变形阶段(Tyreeetal., 2002)。水分导致部分纤维素分子链间的氢键断裂,引发包括纤维素微纤丝弹性模量在内的力学性质降低(Kulasinskietal., 2015),由此可推断,具缘纹孔膜力学性质对栓塞不同阶段其所处的水分环境变化具有敏感性。现阶段,已构建的细胞壁微力学表征方法(Wimmeretal., 1997; Yinetal., 2011)开始逐渐应用于具缘纹孔膜力学性质研究,具缘纹孔膜弹性形变时的弹性模量可通过纳米压痕与纳米弯曲技术结合原子力显微镜进行估算(Capronetal., 2014),探针式微力学系统可测试裸子植物具缘纹孔膜在载荷持续增大条件下的塑性变形(Zelinkaetal., 2015)。但现有微力学测试分析方法仍存在样品制备困难和精度偏低等问题,开展具缘纹孔膜力学性质的理论分析和精确模拟,将有助于揭示木质部栓塞不同阶段纹孔膜的力学行为。
基于具缘纹孔膜的几何结构特征(厚度与直径差别明显),可将其简化为经典板壳结构,采用Kirchhoff-Love等薄板理论分析具缘纹孔膜在两侧压力差作用下的弹性变形规律(Tixieretal., 2014),不过这种唯象型力学分析难以表征具缘纹孔膜变形前后的微观结构差异。为此,可通过具缘纹孔膜的三维结构分析,建立其纤维素微纤丝分布的网络结构模型,如“轮辐”网络模型(Sperryetal., 2004)和随机纤维网络模型(Lietal., 2020),其中基于单根纤维素微纤丝建立的随机纤维网络模型,可用于预测被子植物具缘纹孔膜力学性质及其在拉伸作用力下的位移变化规律。这些研究方法和模拟技术的发展,有助于研究不同水分条件下具缘纹孔膜刚度和强度的变化规律,以及其在微纳尺度下的变形和断裂机制,从而为在纹孔膜微力学尺度理解木质部栓塞形成、扩散和疲劳机制奠定理论基础。
2.4 具缘纹孔膜微液流行为
由于植物细胞次生壁呈现高度木质化,阻碍水分流动(Donaldsonetal., 2018),而具缘纹孔膜的天然多孔隙结构使其成为植物输水管道中水分跨细胞流动的关键部位,因此了解具缘纹孔膜微液流行为有助于明晰木质部网络水分输导功能,明确木质部栓塞过程中水分传导功能的变化。水分传导效率是表征微液流行为的重要指标,采用传统植物生理学方法,难以直接获取具缘纹孔膜尺度的水分传导效率。前人通过低压液流计比较分析木质部在纤维素酶降解前后的水分传导效率,直接对具缘纹孔膜的水分传导效率进行估算,结果显示,纤维素酶对部分裸子植物具缘纹孔膜的降解无效(Schulteetal., 1988),这可能是由于其具缘纹孔膜纹孔塞结构含有丰富的果胶成分。微型毛细管技术(micro-capillary technique)结合低压液流计被用于直接测定被子植物具缘纹孔膜的水分传导效率,但该技术不适用于裸子植物(Choatetal., 2006)。目前,一般多采用低压液流计测定木质部水分传导效率,通过剔除输水管腔和纹孔口的水分传导效率,从而间接获得具缘纹孔膜的水分传导效率(Sperryetal., 2004);但上述方法受限于木材二维解剖特征,无法获得单个纹孔膜的水分传导效率。X射线微计算机断层扫描技术能够实现木质部三维网络结构中纹孔分布成像和量化分析(Brodersenetal., 2016),核磁共振流体成像技术可对木质部液流速率进行无损检测(Windtetal., 2009),在此基础上结合低压液流计与酶解法,将有望实现单个纹孔膜微液流行为的量化分析。
木质部运输水分过程中,具缘纹孔膜的微液流行为受木质部汁液中阳离子含量的影响,阳离子浓度增加可使其水分传导效率提高(离子效应)(van Ieperenetal., 2000)。最初前人认为主要是因为具缘纹孔膜中果胶的存在(Zwienieckietal., 2001),但随着越来越多研究证实具缘纹孔膜中果胶含量很低,该解释逐渐受到质疑(Herbetteetal., 2014; Klepschetal., 2016)。van Doorn等(2011)提出,木质部汁液中阳离子浓度增加引起的具缘纹孔膜部分或整体收缩是导致其孔隙直径增加和液流效率提高的主要原因。具缘纹孔膜化学成分与其微液流行为密切相关,开展具缘纹孔膜微区化学成分研究对理解其三维结构、微液流行为在栓塞过程中的变化规律具有重要意义。
具缘纹孔膜的孔隙特征因树种而异,决定其微液流行为的关键孔隙结构特征有所不同。裸子植物具缘纹孔膜的微液流效率与其孔隙度呈正相关,大尺寸孔隙承担主要的水分输导功能(Schulte, 2012; Schulteetal., 2015)。而决定被子植物具缘纹孔膜水分传导效率的关键因素不是直径最大的孔隙,而是其孔隙通道的阻塞度(Kaacketal., 2019; Lietal., 2020)。目前,尽管多数研究基于计算流体动力学模型所得到的数据与试验结果匹配度较高(Sperryetal., 2005),但一般是将纹孔膜简化为刚体,忽略了变形对水分传导的影响(Schulte, 2012; Schulteetal., 2015),或仅考虑了具缘纹孔膜与水分之间的流固耦合(陈琦, 2016),与具缘纹孔膜栓塞过程中的实际情况不符。事实上,木质部栓塞涉及水分、气体、固体3种介质的相互作用,同时,栓塞过程中具缘纹孔膜在压力差作用下发生变形或断裂,又会反过来影响水分传导效率和气泡扩散规律。但具缘纹孔膜在不同水分条件下的微液流效率如何响应、气泡在具缘纹孔膜三维孔隙结构内以何种方式形成和扩散目前尚不清楚。
植物生长发育过程中,不同生物学宏观性状与木质部微观构造均发挥着非常重要的作用,其间存在一定的权衡关系。例如,在宏观性状方面,油棕(Elaeisguineensis)的植株高度与其木质部栓塞抗性呈正比(Waiteetal., 2019),加州刺柏(Juniperuscalifornica)的枝条体积与其木质部栓塞抗性呈反比(Mahaffeyetal., 2020);在微观构造方面,木质部管道连接度、纹孔膜厚度、纹孔膜孔隙度等主要特征共同决定木质部栓塞抗性(Scholzetal., 2013; Zhaoetal., 2019)。此外,纹孔塞与纹孔口直径之间的比例等构造特征也与木质部栓塞抗性显著相关(Boucheetal., 2014)。这说明,在研究不同树种木质部栓塞形成和扩散规律时,应综合考虑宏观性状、微观特征及其相互之间权衡关系对理解木质部栓塞机制的重要性。
3 优先研究问题及展望
综上所述,围绕木质部具缘纹孔膜,本研究提出木质部栓塞领域应优先开展以下3方面研究: 1) 不同水分条件下具缘纹孔膜化学成分、物理和力学性质的变化; 2) 气泡在水分与具缘纹孔膜三维孔隙结构间水-固耦合界面的形成与扩散方式; 3) 不同水分条件下具缘纹孔膜结构变化及其对微液流效率的影响。具缘纹孔膜三维空间结构的原位精准表征与生物模型构建及优化,仍是亟待突破的关键技术瓶颈,构建具缘纹孔膜三维模型需充分考虑不同树种的纹孔膜类型。在厘清具缘纹孔膜构造特征的基础上,进一步研究不同水分条件下其化学、物理、力学性质和微液流行为的变化以及相互关系,揭示具缘纹孔膜特征对木质部栓塞的影响规律这一关键科学问题,将有助于深入理解木质部栓塞机制,为探究木质部抗旱机制、选育优良耐旱种质资源提供科学依据。
猜你喜欢
中华实用诊断与治疗杂志(2022年1期)2022-08-31
灌溉排水学报(2022年7期)2022-08-08
中国典型病例大全(2022年11期)2022-05-13
医学概论(2022年4期)2022-04-24
大庆石油地质与开发(2022年2期)2022-04-09
长江科学院院报(2022年2期)2022-03-02
天津农业科学(2015年11期)2015-12-03
安徽农业科学(2015年34期)2015-10-21
热带农业科学(2015年6期)2015-07-18
中国民族民间医药·下半月(2014年2期)2014-09-26
