
2,4-表油菜素内酯对干旱胁迫下垂丝海棠生理特性的影响
2022-05-24 07:20缐旭林张仲兴王双成高彦龙王延秀
干旱地区农业研究 2022年3期
缐旭林,张 德,张仲兴,王双成,高彦龙,王延秀
(甘肃农业大学园艺学院,甘肃 兰州 730070)
土壤水分对植物生长发育至关重要。干旱胁迫会引起植物根系呼吸速率下降、呼吸代谢途径改变、呼吸代谢相关酶活性及产物种类变化,造成根系吸收、运输水分和养分等功能紊乱[1]。干旱胁迫下植物主要通过形态变化和渗透调节提高其抗旱性,其中形态变化主要表现为根系向土壤深处伸长、叶片表皮有发达角质层和表皮毛、加快叶片生长速度使老叶脱落和关闭气孔等;渗透调节主要表现为增加脱落酸含量、加快脯氨酸积累、提高与甜菜碱有关的酶类活性、水孔蛋白调控水分通透性、提高抗氧化酶活性清除活性氧等[2]。我国西北黄土高原属于半干旱大陆性季风气候区,干旱已成为限制该地区苹果生产的主要非生物胁迫之一[3]。研究发现,逆境胁迫下在番茄[4]、茶树[5]、黄瓜[6]中喷施外源物质均取得较好的效果。因此,选用适宜的外源物质对植株在逆境胁迫下生长具有重要的意义。
油菜素内酯(BRs)是从油菜素花粉中分离出来的一种具有极强生理活性的物质,已被公认为第六大植物激素,具有增强植物抗逆性的生理功能,广泛应用于缓解植物逆境胁迫[7-8]。2,4-表油菜素内酯(EBR)是目前在农作物上引用较多的BR化合物,其可以维持植物较高的叶绿素含量,减缓膜脂过氧化作用,增强植物的抗逆性[9-10]。盐胁迫下外源EBR能够明显缓解刺槐幼苗的氧化损伤,保护光合细胞器-叶绿体的结构和功能稳定,提升抗氧化能力,维持刺槐叶片光合系统的运转,提高其耐受性[11]。高温胁迫下外源施用油菜素内酯可明显增大叶榕气孔导度,降低蒸腾速率[12]。油菜素内酯也可调节杨树在盐、铜胁迫下叶片PSII最大光化学速率,进而提高其抗逆性[13]。干旱胁迫下适宜浓度的EBR可增强葡萄抗氧化酶活性,减少丙二醛(MDA)的产生,保证其正常生长[14];EBR处理后的羊草幼苗株高、叶面积和脯氨酸(Pro)含量均显著增加,抗旱能力显著提高[15]。外源EBR在不同逆境下均有相关的应用报道,且多见于叶榕、水稻[16]、辣椒[17]等植物,而在木本植物经济林果类中的应用及报道鲜见,亟需研究其在苹果砧木中是否具有相同作用机理。
垂丝海棠(MalushallianaKoehne)原产于甘肃河西走廊地区,耐旱、耐盐碱[18]。本试验以7~8叶龄的垂丝海棠实生苗为试验材料,探究外源喷施不同浓度的EBR对其生理特性的影响,并利用主成分综合分析法筛选出最适浓度,以期为生产实践提供一定的理论指导。
1 材料与方法
1.1 材料与处理
选择长势一致且生长健壮的7~8叶龄的垂丝海棠(MalushallianaKoehne)实生苗移载到大小相同、盛装2 kg基质(20%蛭石,20%珍珠岩,60%泥炭)的塑料花盆内(内径11.2 cm,深16.8 cm),每盆1株,并置于甘肃农业大学避雨棚进行统一管理。移栽14 d后,采用称重法来控制土壤含水量进行干旱胁迫处理,始终保持盆中田间持水量在50%±2%,对照处理始终保持田间持水量的80%。试验共设7个处理,每个处理6个重复,每个重复3株。分别以正常水分(CK)、干旱胁迫(T1)、干旱胁迫+0.20 mg·L-1油菜素内酯(T2)、干旱胁迫+0.40 mg·L-1油菜素内酯(T3)、干旱胁迫+0.60 mg·L-1油菜素内酯(T4)、干旱胁迫+0.80 mg·L-1油菜素内酯(T5)、干旱胁迫+1 mg·L-1油菜素内酯(T6)代表各处理。其中,油菜素内酯溶液以叶面喷施的方式于每日18∶00喷施,每盆均匀喷施50 mL,每隔2 d喷施1次,共喷施3次。干旱胁迫处理次日开始计算胁迫时间,分别于胁迫0、3、6、9 d进行各项指标的测定和分析。
1.2 生理指标的测定
1.2.1 光合及叶绿素含量的测定 光合参数测定:采用LI-6400便携式光合仪(LI-COR,Linco ln,NE,USA)于胁迫0、3、6、9 d上午9∶00—11∶00,选取同一节位的功能叶进行净光合速率(Pn)、蒸腾速率(Tr)、胞间CO2浓度(Ci)和气孔导度(Gs)的测定,每个处理4次重复。
叶绿素含量测定:参照贾婷婷等[19]方法进行测定。
1.2.2 抗氧化酶活性及渗透调节物质含量的测定 于胁迫0、3、6、9 d后,取植株中上部成熟叶片,去除叶脉剪碎并洗净磨碎,用于相关指标的测定。相对电导率(REC)测定采用电导法[20],丙二醛(MDA)含量采用硫代巴比妥酸(TBA)法测定[21]。脯氨酸(Pro)含量采用酸性茚三酮法测定[22]。过氧化物酶(POD)活性的测定采用愈创木酚法[23],超氧化物歧化酶(SOD)活性的测定采用氮蓝四唑光化还原法[24],过氧化氢酶(CAT)活性的测定采用紫外吸收法[25]。每处理4次重复。
1.3 数据处理与分析
用Excel 2010和Origin 2018进行数据处理及作图,用SPSS 22.0进行相关性和主成分分析。统计分析采用单因素ANOVA的LSD比较差异的显著水平(α=0.05)。
2 结果与分析
2.1 外源EBR对干旱胁迫下垂丝海棠叶片光合色素含量的影响
由图1A可知,随干旱胁迫时间的延长,各处理叶绿素a含量均呈持续下降的趋势,但降幅不同。胁迫至9 d时,各处理叶绿素a含量达到最低值,T2处理叶绿素a含量显著低于CK。但显著高于其它处理。与CK处理相比,T1、T2、T3、T4、T5和T6处理下叶绿素a含量降幅分别为40.20%、8.64%、24.10%、29.29%、34.15%和38.00%,说明0.20 mg·L-1EBR处理可降低干旱胁迫下垂丝海棠叶片叶绿素含量a生物合成的抑制作用。
由图1B可知,随干旱胁迫的持续,叶绿素b含量呈不断下降的趋势。胁迫至9 d时,各处理叶绿素b含量达到最低值。其中,叶绿素b含量在T1处理下显著低于其它处理,在T2处理下显著高于其它处理,低于CK。与CK处理相比,T1、T2、T3、T4、T5和T6处理下叶绿素b含量降幅分别为20.86%、2.33%、15.08%、16.72%、17.61%和20.28%,说明0.2 mg·L-1EBR处理可有效降低干旱胁迫下垂丝海棠叶片叶绿素b生物合成的抑制作用。
由图1C可知,叶绿素a+b和叶绿素b含量变化趋势相一致。胁迫至9 d时,叶绿素a+b在各处理下降至最低,均显著低于CK,与CK处理相比,T1、T2、T3、T4、T5和T6处理下叶绿素a+b含量降幅分别为32.24%、6.01%、20.39%、24.12%、27.35%和30.71%。
由图1D可知,各处理下的Chl a/b值呈逐渐下降的趋势。胁迫至9d时,各处理下的Chl a/b值达到最低,其中T2处理显著低于CK,但显著高于其它处理。与CK处理相比,T1、T2、T3、T4、T5和T6处理下叶绿素a/b降幅分别为37.63%、9.68%、16.13%、22.58%、31.18%和35.48%。

图1 外源油菜素内酯对干旱胁迫下垂丝海棠叶片光合色素含量的影响
2.2 外源EBR对干旱胁迫下垂丝海棠叶片光合特性的影响
由图2A可知,随干旱胁迫时间的延长,垂丝海棠叶片的Pn呈逐渐下降的趋势,但不同处理下Pn的降幅不同。胁迫至9 d时,各处理的Pn降至最低,均显著低于CK;与CK相比,降幅分别为25.34%、13.08%、19.86%、21.91%、23.74%、22.87%,说明0.20 mg·L-1EBR处理可有效抑制干旱胁迫下垂丝海棠叶片Pn的下降。
由图2B可知,随干旱胁迫时间的延长,垂丝海棠叶片的Gs与Pn变化趋势相同,但不同处理下Gs的降幅不同。胁迫6 d后,Gs呈现出逐渐下降的趋势,胁迫至9 d时,各EBR处理下的Gs降至最低,显著低于CK,与CK相比,T1、T2、T3、T4、T5和T6处理下Pn降幅分别为67.53%、33.77%、38.96%、45.45%、62.34%和64.94%,说明0.20 mg·L-1EBR处理可有效抑制干旱胁迫下垂丝海棠叶片Gs的下降。
由图2C可知,随干旱胁迫时间的延长,垂丝海棠叶片的Ci呈逐渐上升的趋势,但不同处理下Ci的升幅不同。胁迫至9 d时,不同EBR处理下垂丝海棠叶片的Ci达到峰值,分别为CK的2.53倍、1.27倍、1.79倍、2.03倍、2.24倍和2.35倍。由此可见,适宜浓度EBR能显著抑制干旱胁迫下Ci的升高。
由图2D可知,随着干旱胁迫时间的延长,垂丝海棠叶片的Tr与Gs变化趋势相同,呈逐渐下降的趋势。但不同处理对Tr的降幅不同。胁迫至9 d时,垂丝海棠叶片的Tr出现明显的下降,且各处理下垂丝海棠叶片的Tr达到最低,均显著低于CK,与CK相比,T1、T2、T3、T4、T5和T6处理下Tr降幅分别为45.82%、14.24%、18.58%、26.93%、31.588%和38.70%,说明0.20 mg·L-1EBR处理可有效抑制干旱胁迫下垂丝海棠叶片Tr的下降。
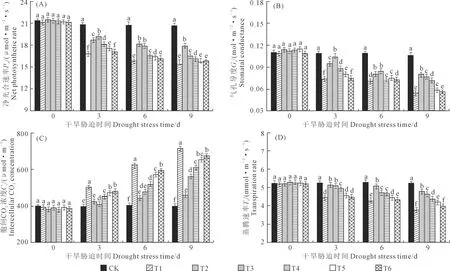
图2 外源油菜素内酯处理对干旱胁迫下垂丝海棠叶片Pn, Gs, Ci, Tr含量的影响
2.3 外源EBR对干旱胁迫下垂丝海棠叶片REC和Pro含量的影响
由图3A可知,随干旱胁迫时间的延长,垂丝海棠叶片REC呈先升后降的趋势。胁迫至3 d时,各EBR处理下垂丝海棠叶片REC出现显著升高,胁迫至6 d时,REC达到峰值,与CK相比, T1、T2、T3、T4、T5和T6处理下REC升幅分别为52.73%、16.98%、27.05%、25.41%、31.22%和38.49%,说明外源EBR对干旱胁迫引起的细胞膜透性变化有缓解作用,从而降低垂丝海棠叶片REC含量。
由图3B可知,随干旱胁迫时间的延长,垂丝海棠叶片Pro含量变化趋势与REC相同。胁迫至3 d时,各处理的Pro含量出现明显的增加,且随着EBR浓度的增加,Pro含量呈逐渐下降的趋势;胁迫至6 d时,各处理下的Pro含量达到峰值,在T1处理积累量最小,为27.33 mg·g-1;T2处理积累量最大,为40.52 mg·g-1。与CK相比,T1、T2、T3、T4、T5和T6处理下Pro升幅为35.13%、95.99%、67.54%、63.62%、44.21%和42.19%,说明外源EBR能进一步促进干旱胁迫下垂丝海棠叶片Pro的积累,且存在明显的浓度效应。
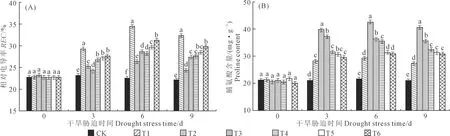
图3 外源油菜素内酯处理对干旱胁迫下垂丝海棠叶片REC和Pro含量的影响
2.4 外源EBR对干旱胁迫下垂丝海棠叶片MDA含量的影响
由图4可知,随胁迫时间的增加,垂丝海棠幼苗叶片的MDA含量呈上升的趋势,且不同处理下MDA含量的升幅不同。胁迫至9d时,各处理下的MDA含量达到峰值,其中T1处理下MDA含量最高,为8.25 μmol·g-1;T2处理下含量最低,为6.52 μmol·g-1,且显著高于CK(5.92 μmol·g-1),与CK相比,T1、T2、T3、T4、T5和T6处理下MDA升幅为39.36%、10.14%、14.36%、15.88%、23.99%和26.01%,说明0.2 mg·L-1EBR处理可有效降低干旱胁迫下垂丝海棠叶片MDA含量,缓解了干旱胁迫对垂丝海棠的伤害。
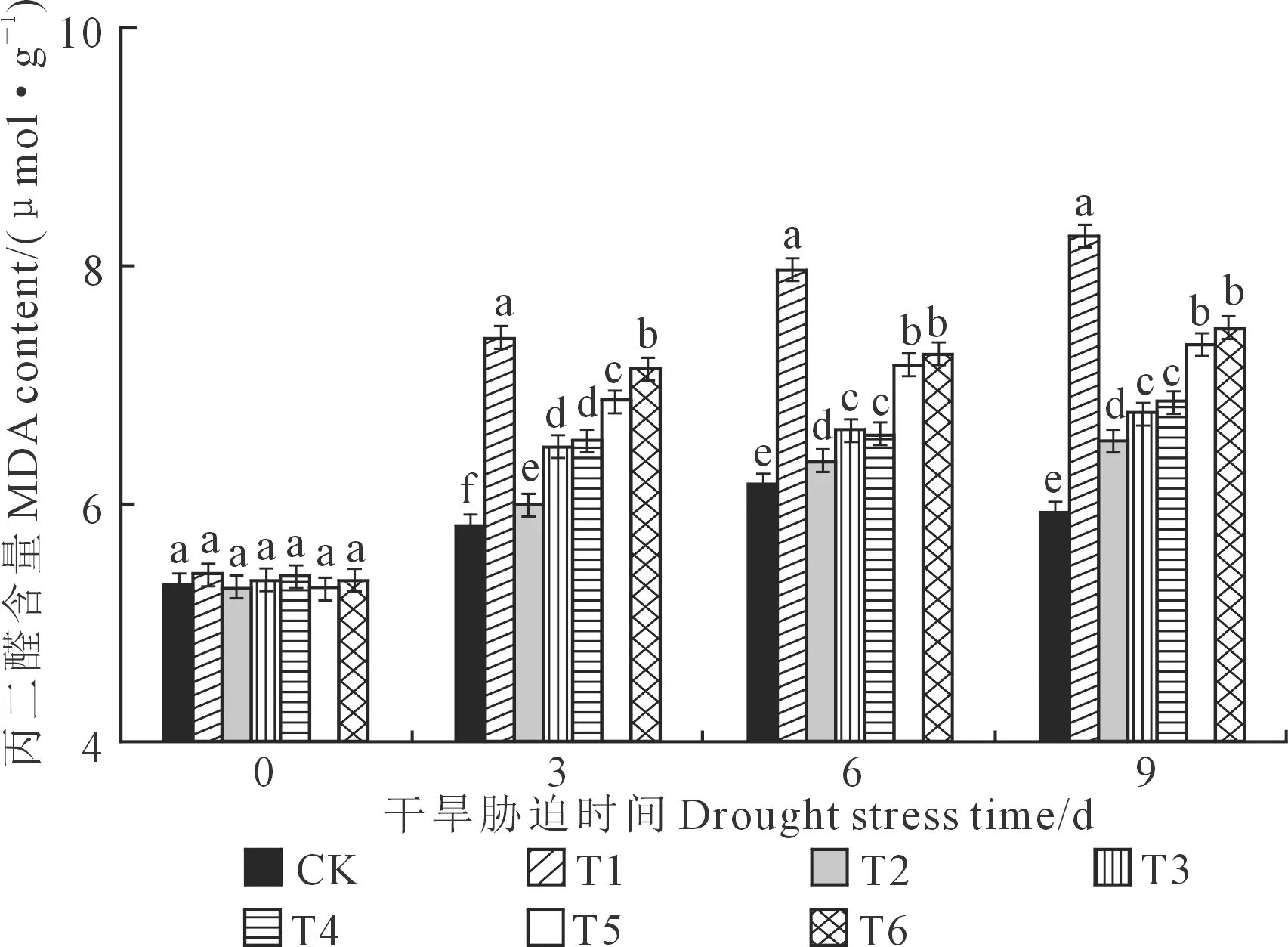
图4 外源油菜素内酯处理对干旱胁迫下垂丝海棠叶片MDA含量的影响
2.5 外源EBR对垂丝海棠叶片抗氧化酶活性的影响
由图5A可知,随干旱胁迫时间的延长,各处理的CAT活性呈先升后降的趋势。胁迫至3 d时,T3的CAT活性达到峰值,为130.41 U·g-1,胁迫至6 d时,T1、T2、T4、T5、T6的CAT活性达到峰值,分别为139.76、145.64、131.71、129.55、127.82 U·g-1,且随EBR浓度的增加,CAT活性呈降-升-降趋势。胁迫至9 d时各处理CAT活性下降,相比CK(125.83 U·g-1),T2(135.17 U·g-1)降幅最大,为7.42%,说明外源EBR增强了植物抗氧化系统的防御能力,维持了细胞内ROS系统的代谢平衡,从而在一定程度上缓解了干旱胁迫引起的氧化损伤。
由图5B可得,随胁迫时间的延长,垂丝海棠幼苗叶片POD活性呈先升后降的趋势。胁迫至6 d时T2的POD活性达到峰值,为198.38 U·g-1,显著高于CK,较CK提高46.73%。胁迫9 d时各处理下的POD活性下降,但各处理的POD活性均显著高于CK,说明外源EBR处理使干旱胁迫下垂丝海棠叶片POD活性升高,有效增强了植株抗氧化能力,从而提高了垂丝海棠抗旱性,其中T2处理效果最佳。
由图5C可知,随着胁迫时间的延长,垂丝海棠叶片的SOD与POD活性的变化趋势相同。各处理分别在胁迫3 d(T1、T6)、6d(T2、T3、T4、T5)达到峰值后不断降低,但不同处理对SOD活性的降幅不同。胁迫至9 d时,SOD活性下降,其中T2处理SOD活性最高(126.76 U·g-1),显著高于CK(88.23 U·g-1),是CK的1.44倍,说明外源EBR能缓解干旱胁迫对垂丝海棠叶片抗氧化酶活性的抑制作用,但浓度过高或过低时,缓解效果都会大大减弱。
2.6 干旱胁迫下不同浓度EBR对垂丝海棠生理效应的综合评价
2.6.1 相关性分析 将处理后垂丝海棠叶片的14个生理指标进行相关性分析(表1)。结果表明,垂丝海棠叶片的Pn与Chl a、Chl b、Chl a+b、Chl a/b、Gs呈极显著正相关(P<0.01),与Tr呈显著正相关(P<0.05),与REC、Ci呈极显著负相关(P<0.01)。

表1 不同处理下垂丝海棠叶片各项指标的相关性分析
2.6.2 主成分分析 为综合评价干旱胁迫下不同浓度EBR对垂丝海棠的生理响应特性,将胁迫处理后的14个指标进行主成分分析,提取特征值大于1的2个主成分,其特征值分别为9.765、3.747(表2)。第一、二主成分方差贡献率为69.747%、26.766%,累积方差贡献率达到96.513%,符合分析要求。如表2所示,第一主成分(PC1)综合了Pn、Chl a、Chl b、Chl a+b、Chl a/b、REC、MDA、Gs、Tr、Ci等指标,第二主成分(PC2)综合了Pro、SOD、POD和CAT等指标。

表2 主成分分析及方差解释
综合得分(F)是每个主成分得分与相对应方差贡献率的乘积之和,即:F=F1×69.747%+F2×26.766%。由表3可知,垂丝海棠在不同处理下的综合得分分别为:-1.20624(T1)、0.880667(T2)、0.318411(T3)、0.068989(T4)、-0.276(T5)和-0.56878(T6)。因此,干旱胁迫下不同浓度EBR对垂丝海棠生理特性的影响排名依次为:T2>T3>T4>T5>T6>T1。

表3 不同处理下垂丝海棠的综合得分及排名
3 讨 论
叶绿素能够吸收和转换光能,是植物进行光合作用的物质基础[26]。干旱胁迫会影响植物体内叶绿素生成,甚至会促进原有的叶绿素分解[27]。马野等[28]在玉米幼苗的研究中发现,喷施EBR可使叶绿素的含量显著增加。本试验中,随胁迫时间的延长,垂丝海棠叶片的Chl a、Chl b、Chl a+b、Chl a/b含量均呈下降趋势,通过不同浓度的外源EBR处理后, Chl a、Chl b、Chl a+b、Chl a/b含量相比T1均有不同程度的上升,这可能是外源EBR处理缓解了干旱胁迫对垂丝海棠叶绿体的损坏,从而保持其结构的完整性,与吴晓丽等[29]在花椰菜中的研究结论相似,进一步证实外源EBR可以有效缓解这种伤害。本试验相关性分析表明,Pn与Chl a、Chl b、Chl a+b、Chl a/b均呈极显著正相关,说明叶绿素含量的降低可能是光合作用减弱的重要因素。有研究表明,气孔限制和非气孔限制均会引起Pn下降[30-31]。若Ci和Gs同时下降,说明气孔因素是引起光合速率下降的主要原因,如果Gs下降,而Ci维持不变甚至上升,则表明叶肉细胞同化能力降低等非气孔因素是引起光合速率下降的主要原因[32]。本试验中,干旱胁迫下垂丝海棠叶片的Gs下降的同时Ci上升,说明在干旱胁迫下垂丝海棠Pn下降是由非气孔限制因素造成,即由垂丝海棠叶肉细胞的光合活性降低引起,这可能是干旱胁迫下细胞过氧化使光合电子传递受阻,影响类囊体蛋白质合成,导致叶片气孔导度降低、CO2同化缓慢、叶片蒸腾速率和光合速率下降[33]。通过不同浓度外源施EBR处理,垂丝海棠Ci升幅显著小于未施EBR处理,Pn、Gs显著升高,说明EBR抑制叶肉细胞光合活性的过度降低,从而提高垂丝海棠的净光合效率[32]。这与丁丹阳等[34]在烟草上的研究结果相一致。
细胞膜作为半透性膜,在细胞内外之间的物质交换和利用方面起着重要作用,相对电导率可评估植物受害程度[35]。干旱胁迫会破坏细胞膜的稳定性,导致细胞内电解质外渗,相对电导率升高[36]。本试验表明,随胁迫时间的延长,REC呈升高趋势,喷施外源EBR后,REC显著下降,说明外源EBR对干旱胁迫引起的细胞膜透性变化有缓解作用,从而降低REC,这与安汶铠等[37]在棉花抗旱上的研究结果相似。Pro含量通常被视作衡量植物抗逆性的指标[23]。本试验研究表明,随胁迫时间的延长,Pro含量急剧增加,可能是干旱胁迫引起蛋白质分解,脯氨酸首先被游离出来[38]。通过不同浓度外源EBR处理,垂丝海棠叶片中Pro含量显著高于对照,这可能是外源EBR直接参与植物生理生长过程而促进了Pro的合成。这与王雨婷等[14]在葡萄上的研究结论相同,说明外源EBR能进一步促进干旱胁迫下Pro的积累,且存在明显的浓度效应。
逆境胁迫下会打破植物体内ROS产生与清除的动态平衡,MDA含量受到影响,引发和加剧生物膜脂过氧化作用,从而对植物体造成伤害[39]。王丽君等[26]研究发现,喷施EBR可使干旱胁迫下烤烟叶片MDA含量下降52.55%。本研究表明,随胁迫时间的延长,垂丝海棠叶片MDA含量呈上升趋势,通过喷施外源EBR处理后,T2(8.2%)升幅显著低于T1(22.9%),表明通过外源EBR处理能有效缓解MDA含量升高,这与王雨婷等[14]在葡萄上的研究结果相同,可能是外源EBR作为抗氧化物质清除植物体内生成的ROS,抑制膜脂过氧化,减少MDA的大量积累,增强膜的稳定性,从而缓解干旱胁迫对垂丝海棠幼苗的氧化伤害[26]。研究发现,超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)三种抗氧化酶在预防ROS毒害方面具有重要作用[40-41]。本试验研究表明,随干旱胁迫时间的延长,SOD、POD、CAT活性呈先升后降的趋势,可能是由于植物在初期自发调节生理活动来增强SOD、POD、CAT活性,及时清除活性氧,但若胁迫持续或加剧,植物不能进行自发调节,导致酶活性下降,植物氧化伤害加剧[14]。通过不同浓度外源EBR处理后,其叶片SOD、POD、CAT活性显著高于未施EBR处理,且具有明显的浓度效应,这可能是外源EBR增强了植物抗氧化系统的防御能力,维持了细胞内ROS系统的代谢平衡,从而在一定程度上缓解了干旱胁迫引起的氧化损伤,这与李一萍等[42]在木薯中的研究结果一致。
4 结 论
干旱胁迫下,外源2,4-表油菜素内酯通过缓解叶绿素的降解,改善叶片的光合能力,通过稳定抗氧化酶活性降低对膜透性损害、增加其稳定性来缓解干旱胁迫,且具有浓度效应,0.2 mg·L-1处理效果最显著。
猜你喜欢
作物学报(2022年9期)2022-07-18
舰船科学技术(2022年11期)2022-07-15
中国农业科学(2022年10期)2022-06-28
中国药学药品知识仓库(2022年5期)2022-04-11
草业科学(2022年3期)2022-03-26
科学导报(2021年33期)2021-06-07
少儿科学周刊·少年版(2021年17期)2021-01-17
少儿科学周刊·儿童版(2021年17期)2021-01-17
阅读(科学探秘)(2020年8期)2020-11-06
中国医药导报(2018年28期)2018-01-18
