
KASP技术及其在牛SNP分型中的应用研究进展
2022-06-24 14:30马士龙谢书琼刘益丽唐娇江明锋
江苏农业科学 2022年11期
马士龙 谢书琼 刘益丽 唐娇 江明锋


摘要:竞争性等位基因特异性聚合酶链式反应(kompetitive allele specific PCR,简称KASP)是一种基于单核苷酸多态性(single nucleotide polymorphism,简称SNP)的新型基因分型技术,可从基因组水平对SNP和插入缺失多态性(insertion and deletion,简称Indel)进行精准分型,被广泛应用于生命科学研究以及医学领域。KASP与其他分型技术相比,支持低、中、高通量SNP检测,具备成本低、特异性好、准确性高等优势,可通过分子检测手段对家畜功能基因进行准确分型,从而达到优良性状的定向改良育种目标。目前,KASP技术已在牛功能基因相关SNP检测、疾病分型等方面应用起来,本文对近年来KASP技术及其在牛SNP基因分型中的应用研究进展进行概述,以期为该技术在牛的产乳、产肉等性状的改良应用方面提供借鉴。
关键词:KASP;SNP基因分型技术;功能基因;SNP检测;疾病分型;改良育种
中图分类号:S823.2 文献标志码:A
文章编号:1002-1302(2022)11-0031-06
收稿日期:2021-10-21
基金项目:四川省国际科技创新合作/港澳台科技创新合作项目(编号:2019YFH0035);四川省重点研发项目(编号:21ZDYF2193);四川省科技计划(编号:21ZDYF2706);西南民族大学研究生创新型科研项目(编号:CX2020SZ40)。
作者简介:马士龙(1996—),男,河南驻马店人,硕士,主要从动物遗传育种与繁殖研究。E-mail: 1471879112@ qq.com。
通信作者:江明鋒,博士,教授,主要从生理基因组学及基因工程研究。E-mail: mingfengjiang@vip.sina.com。
单核苷酸多态性(single nucleotide polymorphism,简称SNP)是动植物基因组中最常见的遗传变异类型之一,可用于构建高密度遗传图谱和标记辅助选择,有利于全基因组选择研究,对目的性状相关的候选基因SNP进行精细定位,可实现对性状的选择。目前,越来越多简单、快速、高效、高通量的SNP检测方法不断被开发出来,竞争性等位基因特异性聚合酶链式反应(kompetitive allele specific PCR,简称KASP)检测技术在基因组学的快速发展下应运而生,它是一种基于已知SNP的终点荧光基因分型技术,可对基因组SNP位点以及插入缺失多态性(insertion and deletion,简称Indel)进行双等位基因分型。国外首次报道出KASP适用于大样本(N>10 000)的少量SNP分型,并且鉴定出与牛结核病相关的多个SNP;我国已将其应用到奶牛隐形遗传缺陷病的检测。
21世纪以来,KASP作为国际上动植物遗传育种主流的SNP分型工具之一,在植物分子标记辅助选择领域应用最多,国内外已经构建出小麦、水稻等多种植物基因组KASP标记库;近几年,国内外不少学者将KASP技术应用在牛、羊等家畜的产乳、生长繁殖等生产性状基因SNP位点挖掘及遗传疾病鉴定方面,且取得了不错的进展。随着KASP技术逐渐成熟,该技术对国内奶牛、肉牛品种的选育及改良具有巨大应用潜力,本文对近年来KASP技术在牛育种领域的研究进展以及应用前景进行总结,以期为将来的育种研究提供理论依据。
1 KASP技术
KASP技术是继第2代测序技术(next generation sequencing,简称NGS)和基因芯片等SNP分析的第3代分子标记技术。2011年,KBiosciences公司开发出基于等位基因特异性寡核苷酸延伸和荧光共振能量转移(fluorescence resonance energy transfer,简称FRET)产生信号的KASP系统,该系统相对其他基因分型系统更容易、更便宜且更灵活,目前KBiosciences公司已被英国政府化学家实验室(LGC)收购。2013年,Lister等首次将KASP技术用于古DNA(aDNA)分析中,由于所需的DNA模板长度通常比Sanger测序(第1代测序)法短得多,并且对于样本产量低、降解水平高的情况,KASP有较高的成功率。另外KASP分型可在短时间内用生成遗传多样性的SNP数据,有助于快速进行重要表型性状调控基因的选择。Semagn等将KASP平台与传统SNP检测平台GoldenGate对比,发现KASP更精确、费用更低且效率更高;与此同时,低密度的KASP标记足以达到质量控制所需的鉴定目的,比高密度简化基因组测序(genotyping by sequencing,简称GBS)方法更有效。Yu等开发了Fluidigm/KASP标记系统,发现仅需11个KASP标记就可以完成品种鉴定。Zhang等利用牛纯合致死基因HH5 的KASP检测技术,与已报道的高分辨率溶解曲线(HRM)检测方法相比,大幅提高了检测效率。Kaur等认为KASP标记正成为利用SNP进行育种的首选标记。
当前,KASP系统在SNP分型方面比其他技术具有一定的优势,如KASP和TaqMan的SNP分型检出率(call rate)虽然都可达到99.9%,但KASP的引物设计和优化更具SNP位点适用性;另外KASP比HRM、Snapshot、质谱法的成本低,相较于Snapshot、质谱法,KASP引物设计简单、试验操作简易。而基于测序的简化基因组测序(GBS)虽说可从大样本量中快速挖掘和鉴定出高密度的SNP变异,但建库费时、数据量大、成本较高,对于筛选到的SNP后续还需验证;面对国内研发的奶牛液相芯片(液相杂交捕获测序),虽然可应用于A2等功能基因和遗传缺陷基因检测方面,但液相芯片仍不能满足低成本单标记检测的要求,针对主基因性状仅需几个SNP标记时,还需要KASP技术在大群体范围内进行检测。因具有设计简单、分型准确度高、成本较低、高效便捷等优势,KASP备受科研工作者的喜爱,而且KASP标记不含凝胶,同时还避免了高密度SNP阵列灵活性差的缺点,可通过荧光检测仪实现基因的快速准确检测,非常适用于高通量的育种研究。
1.1 KASP技术的原理
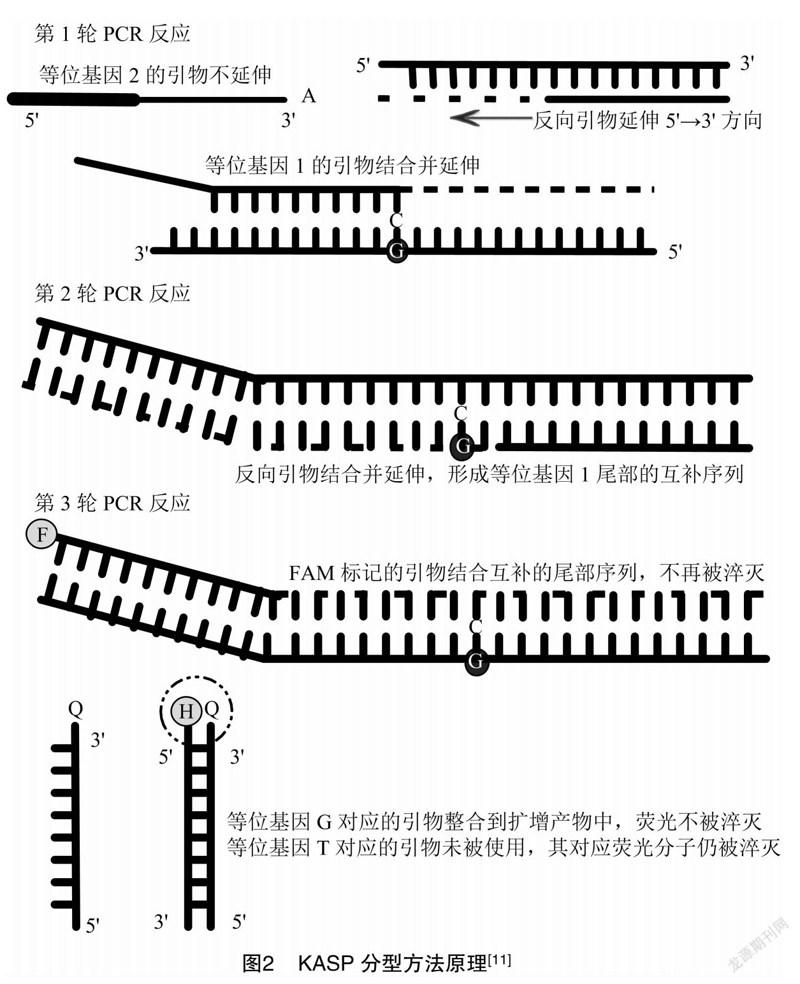
KASP 技术是通过终端荧光读取判断的高通量基因分型方法,其原理是依据引物末端碱基的特异性匹配,利用Touch-down PCR与荧光淬灭探针结合,发出荧光信号来对SNP分型以及检测InDels。该反应体系成分由3个部分构成(图1),第1部分为模板DNA;第2部分为KASP主混合物,包括英国LGC公司的FRET试剂盒(内含2个通用的淬灭荧光探针FAM 和 HEX)、ROX阴性参比染料以及PCR所需的Taq酶等其他成分;第3部分为KASP分析混合物,包含根据SNP位点前后序列特异性而设计的2条竞争性等位基因特异性正向引物和1条反向引物,2条正向引物的5′ 端连接了已知序列的等位基因1或等位基因2,它们分别与FAM 和 HEX对应结合,发出荧光信号。
整个过程主要分为3轮反应(图2)。第1轮PCR:KASP分析混合物中连有等位基因1的正向引物3′端对应其中1条DNA模板链上的SNP,引物与SNP上游结合并退火延伸,完成SNP的识别;反向引物也结合另一条DNA模板链并退火延伸。第2轮PCR:反向引物结合正向引物合成的序列并延伸,合成等位基因1末端序列的互补链,这一步把等位基因1引入与SNP对应的PCR产物。第3轮PCR:等位基因1对应的检测产物随PCR扩增呈指数性增长,淬灭荧光探针FAM退火到新合成的互补链上,发出荧光信号;如果样本SNP的基因型是纯合的,则只会产生1种荧光信号,杂合子的结果是混合荧光信号,从而确定SNP分型。
1.2 KASP试验数据分析
目前,KASP技术的2个核心组分:主混合物和分析混合物,都只能从LGC公司购买,由于主混合物是通用的,可适用于所有KASP试验,须要注意的是,不同荧光定量PCR仪对ROX阴性参比染料的要求不一样,这就要在LGC(https://www.biosearchtech.com)选购适合PCR仪的高、标准、低ROX配方的KASP主混合物。KASP分析混合物是不需要针对每个SNP合成特异的荧光引物,而是根据用户提交的序列,由Kraken软件设计。引物有2种格式,即计算机验证(KBD)和实验室完全验证(KOD),KOD引物是经LGC充分验证和优化后的,相较于KBD更有效,但价格会高一点。
另外,KASP扩增结束后,利用微孔板荧光分析仪、荧光定量PCR仪或带荧光检测功能的酶标仪便可检测FAM和HEX的不同激发波长来区分基因型;值得注意的是:检测机器读取温度都要小于等于40℃,同样是荧光定量PCR仪分析,均不使用实时功能来生成结束点曲线,而是在PCR运行完成后进行读取。如果要进行聚类分析,就要用到KlusterCaller或SNPviewer软件,SNPviewer只是个可视化工具,而KlusterCaller是Kraken的基因分型数据分析和报告模块,当数据导入KlusterCaller软件时,内置的算法会自动辨别结果是等位基因1纯合子、等位基因2纯合子还是杂合子,检测的传统聚类图都会自动生成(图3)。
2 KASP分型技术在牛育种中的应用
2.1 定位生长性状基因SNP位点
牛的体长、产肉量等生长性状是复杂的数量性状,受多基因控制。在牛的选种选育工作上,对生长、生产等参数进行遗传分析,能够对肉牛产业的优选配种工作进行指导。早期,连锁分析法和候选基因法在全基因组范围内鉴定和筛选生长性状的主效基因效果差,由此出现了SNPscan、质谱法、SNP芯片等分型技术,但是这些存在阳性率低、设备要求高、成本偏高等问题,而KASP可胜任牛基因组中高通量DNA变异鉴定,针对家畜生长发育功能基因的定位研究,被科研工作者广泛应用。
Kusza等对山羊生长发育基因GHRHR的 g.62894878A>G 位点进行KASP分型,成功率达94.81%;薛蕾采用KASP对内蒙古绒山羊群体的体质量、体高、体长等生长性状相关的特有候选Indels位点(均小于5 bp)进行鉴定,分型结果清晰明确。这2个研究结果都表明KASP可作为有效的基因分型工具对生长发育基因的SNP位点进行定位。目前KASP在牛生长性状基因上的研究还较少。Pareek等通过KASP分型平台验证了荷斯坦牛的BTA4_77555734、BTA19_27086284、BTA25_ 33248237、BTA16_ 81545856等位点,确定了生长和繁殖候选基因KCNIP4、CCSER1、DPP6、MAP3K5和GHR;这些SNP位点作为新的生长发育标记,有助于研究荷斯坦牛肉质和繁殖性状关键调控机制。颜泽等通过整合KASP技術和微流控芯片建立了针对3个牛无角基因Celtic P位点、Mongolian P位点、Friesian P位点的快速诊断方法,首次在三河牛中检测到Mongolian基因 P位点的存在。证明KASP技术可以有效对牛肉质、体质量、无角等生长性状准确分型,为肉牛业开展优良品种的分子标记辅助选择育种提供了技术手段。
2.2 产乳性状基因SNP定位分型
产乳性状是奶牛的重要经济性状,数量遗传学认为产乳性状(包括产乳量、乳脂量、蛋白含量、乳脂率、蛋白率等) 是由微效多基因决定的数量性状,也是分子标记辅助选择的理想性状之一。近些年,科研工作者利用SNP分型技术如质谱法、PCR-SSCP等技术对奶牛产乳性状的候选基因及相关变异位点进行检测,但都有操作繁琐、样本质量要求高、试验设备昂贵等缺点,而KASP技术具备灵活性高、样本需求低、设备要求低、分型准确率高等特点,非常适宜定位奶牛乳性状的重要候选基因。
如今,国内外已经用KASP技术分型到了多个SNP位点与PRLR、CSN3等产乳性状候选基因相关。Fontanesi等利用KASP技术鉴定出荷斯坦牛的催乳素受体编码基因PRLR的20: g.39071965C>T位点与产乳量、乳脂量等性状显著相关(P<0.05);酪蛋白编码基因CSN2、CSN3的SNP位点分别与乳蛋白率、乳蛋白量相关;这进一步锁定了荷斯坦牛高日产奶量的基因位点,加快了奶牛的定向育种。Cosenza等首次发现意大利水牛催乳素受体编码基因PRLR的20: g.11188 A>G位点(AA型)并进行KASP分型,发现AA型与牛乳脂肪酸等乳脂性状有显著影响(P<0.05),尤其与奇数和支链脂肪酸(OBCFA)紧密相关;此多态性位点可用于标记辅助选择提高牛乳品质。赵凤等报道了一项基于KASP技术检测奶牛κ型-酪蛋白编码基因CSN3A、B、E这3种常见基因型的方法,依据此技术,奶牛中心成功筛选出120头高产乳量BB基因型北京地区奶牛。武泽文等为了筛选β型-酪蛋白A2纯合基因型荷斯坦奶牛,在奶牛场进行 KASP基因分型,结果成功检测到A2纯合基因型。说明 KASP方法可以高效快速地对奶牛基因型进行筛选,并且该方法比PCR-SSCP(单链构象多态性)、基因芯片等方法更适合大样本检测高产奶BB基因型奶牛个体。Abousoliman等用KASP对泌乳性状基因的SNP进行分型,发现瘦素LEP(rs420693815)、信号传导及转录激活因子STAT5A(rs161082816)与生长发育和泌乳性状显著相关,而LEP、STAT5A也在牛的泌乳性状中发挥着重要作用,相关基因调控机制可进一步在牛上验证。
综上所述,产乳量、乳脂等性状SNP位点的KASP分型结果为我国奶牛优良性状的QTL定位和全基因组关联研究提供了重要信息,定位到的产乳候选基因PRLR、CSN3、LEP、STAT5A等SNP位点可用于我国荷斯坦奶牛的分子标记辅助选择工作中。
2.3 遗传缺陷疾病分型及检测
21世纪以来,我国为了提高奶牛的生产性能,在进口大量胚胎、种牛和冷冻精液的同时也导致了遗传缺陷在国内奶牛产业传播,造成了巨大的经济损失。近几年,全基因组测序技术在荷斯坦牛中鉴定出了牛白细胞黏附缺陷病(BLAD)、脊椎畸形综合征(CVM)、荷斯坦单倍型HH1、HH3、HH4、HH5以及胆固醇缺乏单倍型(HCD)和短脊柱综合征(BS)等隐性遗传缺陷病,这迫切需要分子检测方法对奶牛个体的缺陷基因进行研究。近几年,SNP芯片、PCR-RFLP(限制性片段长度多态性)、TaqMan探针法、高分辨率溶解曲线法、KASP等检测方法出现,其中属KASP最高效、经济,可以对样本进行高通量的检测。
2013年,le Roex等以868头非洲水牛为研究对象,通过KASP分型确定了与水牛结核病(BTB)显著相关的3个SNP:SNP41、SNP137、SNP144,它们分别位于SLC7A13、DMBT1、IL1a基因上,这为后续的牛结核病检测方法提供了理论依据。热应激几乎是所有奶牛产业都无法摆脱的难题,Ortega等使用KASP对热休克蛋白HSPA1L进行基因分型,发现HSPA1L的缺失突变可使牛胚胎抵抗热休克和高氧2种环境;如果选育出耐热品种,使牛获得抵抗热休克的能力,可大大降低奶牛产业的经济损失。孙东晓等通过设计BLAD的ITGB2基因突变位点1: g.145114963A>G的KASP引物,为大规模筛查携带BLAD荷斯坦奶牛的工作节省资源成本。2018年,孙东晓等再次发明了基于KASP技术的脊椎畸形综合征(CVM)检测方法,设计1组针对CVM的SLC35A3基因突变位点3: g.43412427G>T的引物,可将CVM携带牛准确分型,分型结果与梁若冰等的检测结果一致,且此方法工作效率较高,适合大批量地检测筛查BLAD和CVM荷斯坦奶牛。Lehner等采用KASP分型对荷斯坦奶牛的皱胃左方移位(LDA)症的SNP进行了验证分析,筛选出SLC4A3、EPHA4、CDH18、PAGE、KIAA1468等候选基因与LDA显著相关(P<0.05)。针对LDA的分子检测可提前筛查奶牛健康状态,避免因皱胃移位导致消化机能紊乱,产奶量骤降造成经济损失。2019年,吕小青等对荷斯坦单倍型HH1的APAF1基因上5: g.63150400C>T位点以及HH3的SMC2基因的g.95410507T>C位点进行KASP分型,发现荷斯坦奶牛群体HH1携带率达12.66%。而2020年,Zhang等研究8种荷斯坦牛常见遗传缺陷基因型频率,用相同方法确定了中国荷斯坦牛群最高的CVM的携带率10.5%,HH1次之(10.0%),其次是HH3(2.6%)、BS(2.1%)、HCD(1.3%)、HH5(0.8%)、BLAD (0.5%),与前者相符;说明荷斯坦奶牛群体的遗传缺陷病在国内已经普遍存在,有必要在国内排查携带隐性遗传缺陷基因牛。與此同时,Sieck等还发现荷斯坦犊牛下颌面部骨质疏松症(MD)的原因,通过KASP分型验证出牛胚胎时期CYP26C1错义突变(Chr26g.14404993T>C)导致维甲酸(RA)的失调,从而影响了内皮素-1信号通路,造成了MD的发生。以上KASP基因分型方法可简单、准确、快速鉴定出多种牛遗传缺陷基因,降低奶牛场有害突变疾病频率,并合理安排选种选配工作,为选育更健康的荷斯坦牛提供策略。
3 结语与展望
目前,SNP基因分型技术主要有HRM、Snapshot、质谱法、TaqMan、KASP,以及基于测序法鉴定SNP的GBS和液相芯片等;相较于其他技术,KASP更适合低、中、高通量SNP分型研究,但KASP仍有不足之处,例如荧光信号的定量处理会产生分型错误;同时也不能像直接测序法发现未知SNP位点,当突变等位基因数量少于野生型等位基因的5%时,很难区分突变或野生型的个体。如今,KASP技术与基因精准分型的研究需求有一定差距,但分子标记技术正在快速发展,引物的优化改良工作也将不断被突破,而KASP技术结合微流控芯片的运用正在打开新的大门,相信未来KASP技术进一步成熟。
总而言之,KASP作为一项高效、灵活、经济、准确的SNP分型检测方法,适合在大规模的群体中完成高通量的SNP检测与候选基因的精细定位;在全基因组关联分析(GWAS)分析得到与性状关联的SNP位点之后,进行大群体小范围SNP的KASP基因分型,可对关联信号的可靠性进行验证;还可通过开发目标性状的KASP标记结合QTL遗传连锁分析来进一步缩小定位区段。在动物产肉、产奶等重要性状的研究中发挥着越来越重要的作用;该项技术仍在动植物育种研究领域具有应用潜力,将来KASP技术会更加深入和广泛地被应用于牛的各方面研究,得到进一步的推广和应用,这将加快牛的育种进程,未来在新品种育种工作中发挥更大价值。
参考文献:
[1]曲娟娟,于 敏,董 蕾,等. SNP在动植物遗传育种中的应用[J].东北农业大学学报,2011,42(12):9-13.
[2]He C,Holme J,Anthony J. SNP genotyping:the KASP assay[J].Methods in Molecular Biology,2014,1145:75-86.
[3]Siva P,Buyyarapu R,Ibrokhim Y,et al.Genomics-assisted plant breeding in the 21st century:technological advances and progress[M].London:InTech,2012.
[4]le Roex N,Koets A P,van Helden P D,et al.Gene polymorphisms in African buffalo associated with susceptibility to bovine tuberculosis infection[J].PLoS One,2013,8(5):e64494.
[5]孙东晓,梁伟俊,梁若冰,等. 一种筛查荷斯坦奶牛白细胞黏附缺陷病携带者的分子检测方法:CN105177136A[P].2019-06-07.
[6]王志伟,乔祥梅,王志龙,等. 云南小麦品种(系)抗逆性相关基因的KASP标记检测[J].西南农业学报,2020,33(8):1601-1607.
[7]韦 宇,李孝琼,何新柳,等. 基于KASP技术的稻瘟病抗性基因Pi9分子标记的开发与评价[J].西南农业学报,2019,32(6):1216-1222.
[8]赵 凤,刘 林,杨宇泽,等. 基于KASP技术检测κ-酪蛋白基因型的方法:CN109321639A[P].2019-02-12.
[9]Fontanesi L,Calò D G,Galimberti G,et al. A candidate gene association study for nine economically important traits in Italian Holstein cattle[J].Animal Genetics,2014,45(4):576-580.
[10]武澤文,顾亚玲,张 娟,等. A2纯合基因型荷斯坦奶牛的筛选[J].黑龙江畜牧兽医,2019(8):44-47.
[11]薛 蕾. 山羊Indels筛选鉴定及其与生产性状关联分析[D].重庆:西南大学,2018.
[12]赵睿骁. 金川多肋牦牛的分子标记筛选及分型[D].成都:西南民族大学,2020.
[13]颜 泽,肖 炜,张胜利,等.基于KASP技术的牛3种无角基因联合检测方法研究[J].农业生物技术学报,2021,29(3):610-618.
[14]孙东晓,梁伟俊,梁若冰,等.一种筛查荷斯坦奶牛脊椎畸形综合征携带者的分子检测方法:CN105087571A[P].2015-11-25.
[15]梁若冰,谢 岩,范学华,等.中国荷斯坦公牛脊椎畸形综合征和尿苷酸合酶缺乏症遗传缺陷检测[J].中国奶牛,2014(7):22-26.
[16]吕小青,劳兰兰,赵 凤,等.北京地区荷斯坦母牛群体HH1遗传缺陷基因抽样调研[J].中国奶牛,2019(1):12-14.
[17]吕小青,刘 林,劳兰兰,等.遗传缺陷单倍型HH3检测方法的建立与应用[J].黑龙江动物繁殖,2019,27(5):16-18.
[18]Robinson P, Holme J. KASP version 4.0 SNP genotyping manual[EB/OL]. (2014-3-12)[2021-06-20]. http://www.cerealsdb.uk.net/cerealgenomics/CerealsDBNEW/PDFs/KASP_SNP_Genotyping_Manual pdf.
[19]Lister D L,Jones H,Jones M K,et al. Analysis of DNA polymorphism in ancient barley herbarium material:validation of the KASP SNP genotyping platform[J].Taxon,2013,62(4):779-789.
[20]Cavanagh C R,Chao S,Wang S,et al. Genome-wide comparative diversity uncovers multiple targets of selection for improvement in hexaploid wheat landraces and cultivars[J].Proceedings of the National Academy of Sciences of the United States of America,2013,110(20):8057-8062.
[21]Semagn K,Babu R,Hearne S,et al. Single nucleotide polymorphism genotyping using kompetitive allele specific PCR (KASP):overview of the technology and its application in crop improvement[J].Molecular Breeding,2014,33(1):1-14.
[22]Ertiro B T,Ogugo V,Worku M,et al. Comparison of kompetitive allele specific PCR (KASP) and genotyping by sequencing (GBS) for quality control analysis in maize[J].BMC Genomics,2015,16:908.
[23]Yu H J,Jeong Y M,Lee Y J,et al. Marker integration and development of Fluidigm/KASP assays for high-throughput genotyping of radish[J].Horticulture,Environment,and Biotechnology,2020,61(4):767-777.
[24]Zhang Y,Liang D,Huang H T,et al. Technical note:development and application of KASP assays for rapid screening of 8 genetic defects in Holstein cattle[J].Journal of Dairy Science,2020,103(1):619-624.
[25]Kaur B,Mavi G S,Gill M S,et al. Utilization of KASP technology for wheat improvement[J].Cereal Research Communications,2020,48(4):409-421.
[26]Li B W,Liu Y R,Hao X D,et al. Universal probe-based intermediate primer-triggered qPCR (UPIP-qPCR) for SNP genotyping[J].BMC Genomics,2021,22(1):850.
[27]吕世杰,陈付英,金 磊,等.利用簡化基因组测序筛选安格斯牛生长相关的受选择基因[J].畜牧兽医学报,2020,51(4):713-721.
[28]徐云碧,杨泉女,郑洪建,等.靶向测序基因型检测(GBTS)技术及其应用[J].中国农业科学,2020,53(15):2983-3004.
[29]Rasheed A,Wen W E,Gao F M,et al. Development and validation of KASP assays for genes underpinning key economic traits in bread wheat[J].Theoretical and Applied Genetics,2016,129(10):1843-1860.
[30]Burow G,Chopra R,Hughes H,et al. Marker assisted selection in Sorghum using KASP assay for the detection of single nucleotide polymorphism/insertion deletion[J].Methods in Molecular Biology,2019,1931:75-84.
[31]国 鹏,封 元,顾亚玲,等. 黑安格斯牛UCP基因多态性位点与生长性状的关联分析[J].黑龙江畜牧兽医,2019(15):6-9.
[32]张静静. 西门塔尔牛部分生长性状全基因组低密度芯片筛选[D].长春:吉林农业大学,2015.
[33]Kusza S,Cziszter L T,Ilie D E,et al. Kompetitive allele specific PCR (KASP) genotyping of 48 polymorphisms at different caprine loci in French Alpine and Saanen goat breeds and their association with milk composition[J].PeerJ,2018,6:e4416.
[34]Pareek C S,Smoczyński R,Kadarmideen H N,et al.Single nucleotidepolymorphismdiscovery inbovine pituitary gland using RNA-seq technology[J].PLoS One,2016,11(9):e0161370.
[35]饒文波. 引物设计及其在EGFR基因分型中的应用[D].西安:西北大学,2010.
[36]王洪亮,李志雄,刘小林,等. 牛PGLYRP3、PGLYRP4基因多态性及其与SCS、产奶性状关联分析[C]//2015中国遗传学会大会论文摘要汇编. 昆明:中国遗传学会, 2015.
[37]Cosenza G,Iannaccone M,Auzino B,et al. Remarkable genetic diversity detected at river buffalo prolactin receptor (PRLR) gene and association studies with milk fatty acid composition[J].Animal Genetics,2018,49(3):159-168.
[38]Abousoliman I,Reyer H,Oster M,et al. Analysis of candidate genes for growth and milk performance traits in the Egyptian barki sheep[J].Animals,2020,10(2):197.
[39]Ortega M S,Rocha-Frigoni N A S,Mingoti G Z,et al. Modification of embryonic resistance to heat shock in cattle by melatonin and genetic variation in HSPA1L[J].Journal of Dairy Science,2016,99(11):9152-9164.
[40]Lehner S,Zerbin I,Doll K,et al. A genome-wide association study for left-sided displacement of the abomasum using a high-density single nucleotide polymorphism array[J].Journal of Dairy Science,2018,101(2):1258-1266.
[41]Sieck R L,Fuller A M,Bedwell P S,et al. Mandibulofacial dysostosis attributed to a recessive mutation of CYP26C1 in Hereford cattle[J].Genes,2020,11(11):1246.
