
基于SSR分子标记的重瓣萱草品种遗传多样性分析
2022-06-28 01:17张贺蕾
江西农业学报 2022年4期
张贺蕾,陈 芬,马 丽,刘 博
(1.江苏省农业科学院 宿迁农科所,江苏 宿迁 223800;2.大连海洋大学 科技与环境学院,江苏 大连 116023)
0 引言
萱草(Hemerocallis fulva)又名金针菜、鹿箭、忘忧草等,是阿福花科(Asphodelaceae)萱草属(Hemerocallis L.)的多年生宿根草本花卉。其品种种类繁多,花色丰富多彩,花型千变万化,适应性强,喜阳光又耐半荫,喜干旱又耐湿润。萱草属植物有15~18种,主要分布于亚洲温热带至热带地区,原产于我国的约有11种[1]。重瓣萱草(Hemerocallis fulva var. kwanso Regel)多为雄蕊瓣化,花瓣数多于6,根肉质,叶片较宽,早上开花晚上凋谢,花型独特,可分为芍药型重瓣和套叠型重瓣[2];因其花瓣多重层次感强,近年来还出现了重瓣卷边型、重瓣花心环型、重瓣镶边型等[3]。由于重瓣萱草的自然结实率偏低,其繁殖方式以分株繁殖为主[4],多丛植、片植、行植于花带和花境中,也可作树林地被植物。此外,萱草是我国传统的母亲花,寓意着伟大的母爱,近年来,越来越多的优新品种流行于家庭园艺。同时,萱草作为金针菜的近亲,依托黄花菜产业不仅促进了休闲农业的打造,促进了金针菜产业发展的提档升级,而且是“美丽乡村”建设中不可多得的乡土植物[5]。
遗传多样性是一个群体或者种内个体之间的遗传变异总和。不同物种的遗传多样性都是独一无二的,都储存在其基因中,是长期进化的产物。目前,与国外相比,我国重瓣萱草育种工作进展较为缓慢,应用于我国的重瓣萱草大部分都是国外培育的品种。主要是由于不断的反复杂交,没有新的基因加入,从而使其种间差别不显著。因此,利用分子生物学手段,从分子水平研究重瓣萱草品种的亲缘关系和遗传背景是十分重要的[6]。
SSR荧光标记毛细电泳法是近年来发展起来的一种以DNA测序仪为平台检测PCR扩增产物的一种方法[7];与银染法相比,荧光测序技术每个位点多检测3个等位变异,检测效果更加准确、高效[8]。它不仅实现了扩增产物电泳的自动化,而且实现了电泳数据记录的自动化[9],该方法已在研究样本遗传多样性、基因鉴定中得到应用。本研究选用34个重瓣萱草品种作为实验材料,利用荧光标记毛细管电泳技术进行分子标记检测,从DNA水平上探讨了重瓣萱草的遗传多样性和多态性水平,并根据其遗传相似系数进行了聚类分析,以期加快发掘优异的基因资源,推进重瓣萱草种质资源的分子育种研究,为今后培育更多的重瓣萱草新品种提供参考和依据。
1 材料和方法
1.1 实验材料
实验材料为从国内外收集的32种重瓣萱草品种:残酷的命运、粉娃娃、摩西火、双子座侍者、四十二街、紫色糖、粉红、钢琴男子、午后、谎言和口红、太空海岸雪天使、强盗、边境松露、国家的骄傲、荷叶双摺边、天马行空富丽、夏娃、摩洛哥夏季、独领风骚、快乐的流氓、黄鹤楼、中国山、事理情歌、莱邦、命定、蕾丝方巾、惊艳、黄色公主、冰淇淋、粉红女孩、双面娇娃、双重美味。还有2种重瓣萱草品种是以‘酒红341’(单瓣型)作为母本,‘蕾丝方巾’作为父本,杂交后获得的F1代品种:15087-3、15087-5。供试材料(表1)均栽植于江苏省农业科学院宿迁农科所萱草种质资源圃内,采用分株繁殖栽培方式,每个材料分割成3株,株距为20 cm左右,行距25 cm左右,并将其标号。采用正常的水肥管理方法,定期除草,使其始终保持干净的生长环境。

表1 34个重瓣萱草品种信息表
1.2 用CTAB法提取植物基因组DNA
DNA的提取采用优化后的CTAB法:取34个重瓣萱草品种3~5 g新鲜材料的叶子或幼芽,剪碎,放到已加有钢珠的2 mL的离心管中,再将离心管放入盛有液氮的保温桶中,震荡,磨碎;在其解冻之前迅速加入65 ℃预热的CTAB提取液600~700 μL,迅速摇匀;将粗提取液装入1.5 mL的离心管中,在65 ℃水浴20 min,期间不时摇动;随后将离心管放进离心机,以8000 r/min离心10 min,离心后轻拿轻放,为避免吸入杂质,将上清转到新的1.5 mL的离心管中;加入Glass Milk 3~5 μL和600 μL DNA Binding Buffer(或6 mol/L NaCl),充分混匀,在65 ℃水浴15 min,中间需翻转;室温放置5 min,放入离心机,4000 r/min离心1 min,弃上清,留管底,加入500 μL无水乙醇,吹打悬浮,洗涤,8000 r/min离心1 min;弃上清,重复上一步,再次加入500 μL无水乙醇,吹打悬浮,洗涤,8000 r/min离心1 min,弃上清,室温干燥;最后加100 μL TE(pH 7.0),70 ℃水浴5 min,离心,吸取上清转移至新的离心管中,放冰箱-20 ℃保存。
1.3 荧光毛细管电泳SSR-PCR扩增体系
反应体系总体积为25 μL,其中包含模板DNA(20 ng/μL)2 μL、10×Buffer 2.5 μL、dNTP(25 mmol/L)2 μL、荧光标记(FAM)正向SSR引物(5 μmol/L)1 μL、方 向 引 物(5 μmol/L)1 μL、rTap酶(5 U/μL)0.2 μL、ddH2O 16.3 μL。扩增程序为:95 ℃预变性5 min;95 ℃变性1 min;55~59 ℃退火45 s,72 ℃延伸45 s,35个循环;最后60 ℃延伸30 min,4 ℃保存。
1.4 荧光毛细管电泳位点信息监测
将PCR产物移入分析区无菌通风橱上进行后续操作。取96孔板,在板的侧壁上写上上机板号,如SSR_20210501-YuMI-1E。取20 μL Rox500内标,加入980 μL去离子甲酰胺内,涡漩混匀,离心,以每孔8 μL分装到新的96孔板内,再取2 μL PCR产物对应地加入各孔中(加PCR产物时检查2个板是否对应一致,是否将96孔板放倒,确保每孔对应正确)。最后将PCR产物板盖上密封膜,存留;将测序跑样板盖上封口膜,短暂离心后,于95 ℃变性5 min,结束后立刻放到冰盒内保存5~10 min;然后将封口膜慢慢揭掉,更换96孔胶皮垫,离心。打开ABI3730xl仪器及电脑,待稳定后打开信号收集软件。
1.5 数据分析
待34个重瓣萱草电泳结束后,用GeneMapper 4.0软件对样品的原始数据进行分析,直接读取样品SSR片段长度,再利用Popgene和Power Marker V 3.25进行数据转换并分析,计算出主等位基因频率、基因型数、等位基因数、基因多样性、杂合度、多态性信息含量等遗传多样性相关指标。利用NTSYS-pc 2.10 e算出遗传相似系数,绘制聚类树状图。
2 结果与分析
2.1 SSR引物筛选及优化
本实验引物均由青岛科创质量检测有限公司合成。共合成302对引物,并根据SSR Hunte检测到的SSR位点进行筛选,初步筛选出PCR相对稳定、具有多态性、重复性好的12对高多态性引物(表2),并将其优化后,对34个重瓣萱草品种进一步进行位点检测。

表2 12对SSR引物信息表
2.2 多态性分析
34个重瓣萱草品种在12对高多态性SSR引物上进行扩增,扩增结果见表3。12对高多态性SSR引物共获得111个等位基因,平均9.2500个,34个重瓣萱草品种的遗传多样性变化(基因多样性)范围为0.5826~0.9277,多态性信息含量(PIC)变 化 范 围 为0.5684~0.9236,最 高 的 均 为P5,最低的均为P7。遗传多样性平均值为0.8043,多态性信息含量(PIC)平均值为0.7875,Shannon信息指数(I)平均值为1.8753,观测杂合度变化范围为0.1333~0.7333,期望杂合度变化范围为0.5057~0.9385(表4),说明34个重瓣萱草品种之间遗传基础广,遗传多样性处于较高水平。在位点P5的遗传多样性水平偏离平衡程度最低,在位点P7的遗传多样性水平偏离平衡程度最高。
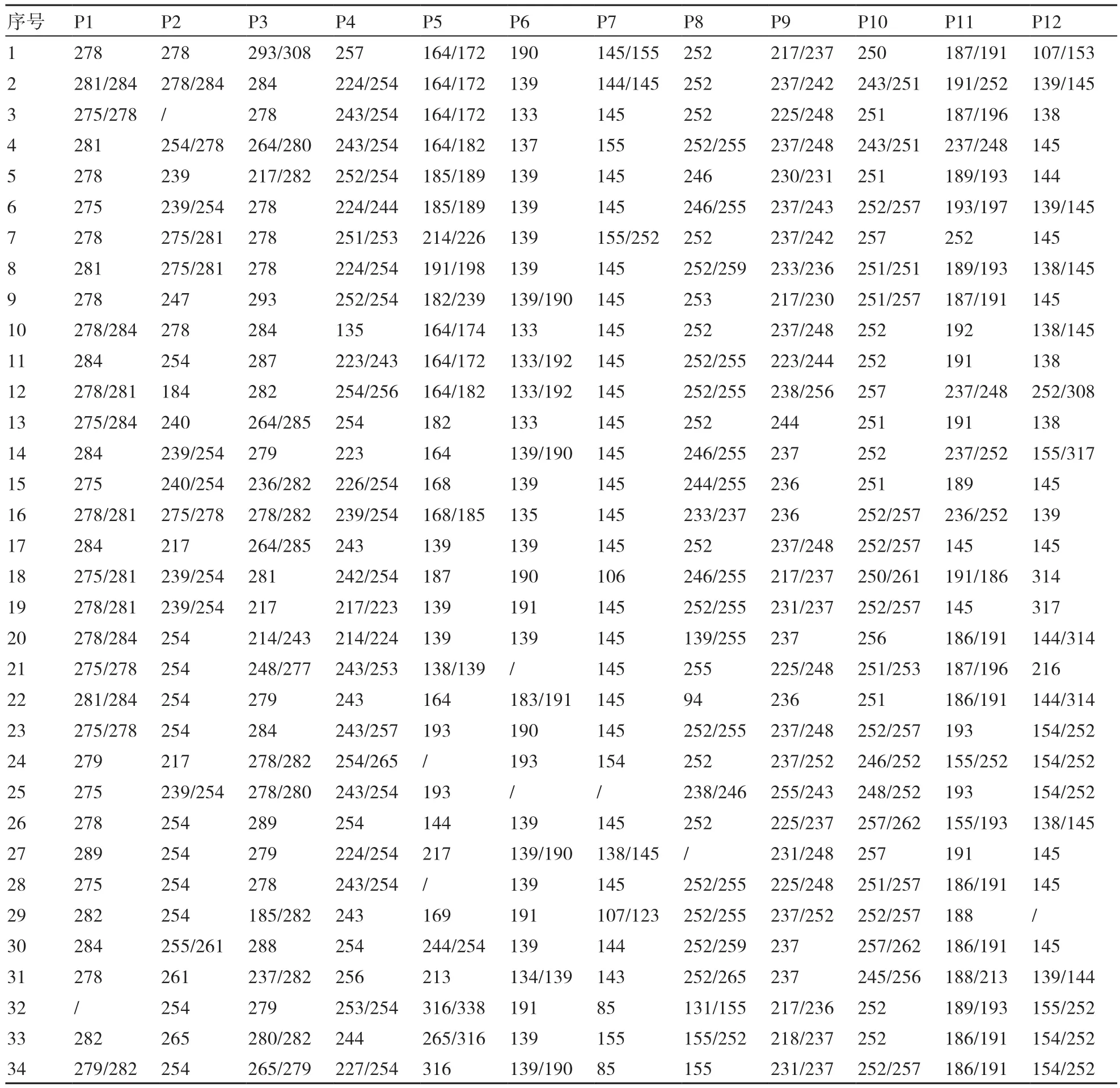
表3 12对SSR引物对34个重瓣萱草品种的扩增结果
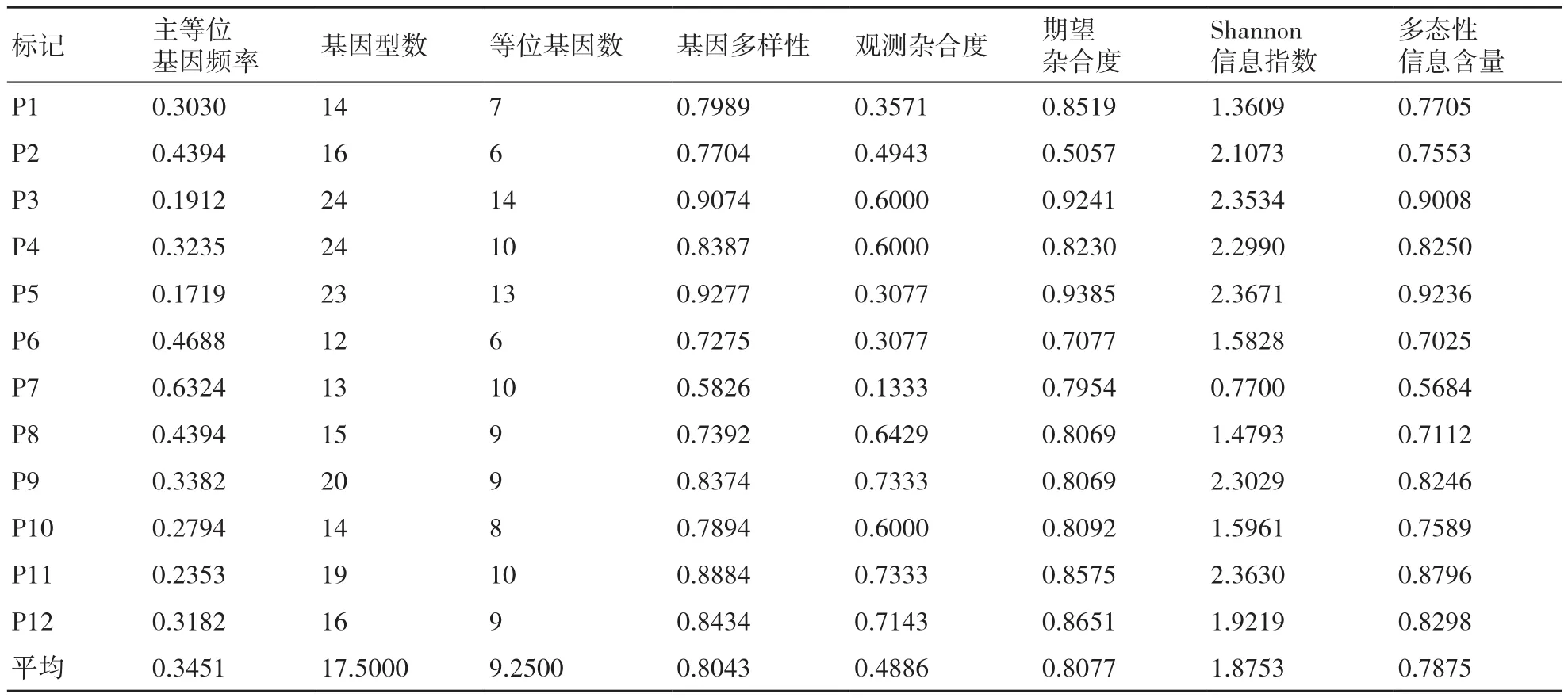
表4 12对引物对34个重瓣萱草品种的SSR遗传多样性
2.3 UPGMA聚类分析
34个重瓣萱草品种基于遗传相似系数矩阵,采用UPGMA方法进行聚类分析,根据聚类树状图(图1),在距离系数L=0.4128处,可将这34个种质划分为两大类群。‘中国山’‘双面娇娃’‘粉红女孩’‘双重美味’4个重瓣萱草品种聚成类群Ⅰ,‘残酷的命运’‘粉娃娃’‘摩西火’‘双子座侍者’‘四十二街’等30个重瓣萱草品种聚成类群Ⅱ,其中重瓣萱草‘摩洛哥夏季’在L=0.4108时与类群Ⅱ聚在一起,说明重瓣萱草‘摩洛哥夏季’与类群Ⅱ的相似性大于与类群Ⅰ的相似性,在L=0.4128时聚到一起。这表明两大类群之间亲缘关系较远,存在较大的遗传差异。
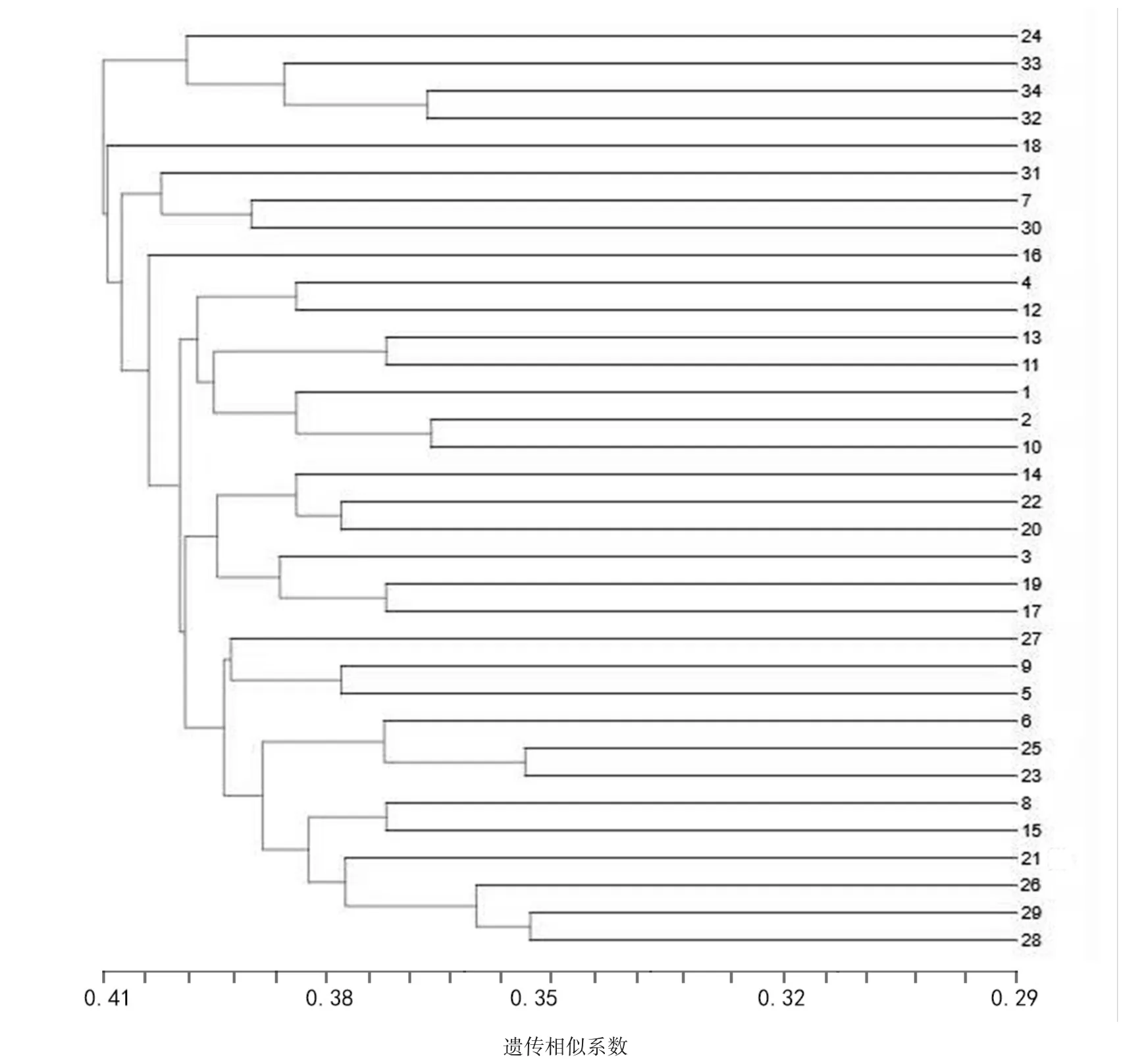
图1 UPGMA法构建的34种重瓣萱草品种的树状聚类图
类群I可划分为2个亚类:(I)亚类和(Ⅱ)亚类,其中(I)亚类为24号‘莱邦’,(Ⅱ)亚类又分A、B 2个小亚类。在B小亚类中,32号‘双面娇娃’、34号‘双重美味’聚类在一起,与A小亚类中的33号‘粉红女孩’聚成(Ⅱ)亚类。这一类群中4个重瓣萱草品种内外花被片颜色相同,且红色系为主要色系,花瓣宽度较大,叶片宽度均较宽,着花密度较大。
类群Ⅱ包括重瓣萱草‘残酷的命运’‘粉娃娃’‘摩西火’‘双子座侍者’等共计30个重瓣萱草品种,其中7号‘粉红’、30号‘黄色公主’组成小类,与31号‘冰淇淋’聚在一起,再与(Ⅲ)亚类聚成Ⅱ大类。在所有供试的重瓣萱草品种中,28号‘15087-3’、29号‘15087-5’最早聚在一起,说明这2种重瓣萱草亲缘关系极其接近,它们的相似性最高,遗传差异最小,聚类后又与26号‘蕾丝方巾’在0.2500时聚在一起,这与28号‘15087-3’和29号‘15087-5’是以‘酒红341’作为母本,‘蕾丝方巾’作为父本杂交后所得F1代品种相一致。这一类群中的30个重瓣萱草品种颜色多样,有红色、紫红、橙红、橙黄、黄色、橙色六大色系,且花序形态多样,叶丛姿态均为半直立,叶丛高度均为中等,直径较小,内外花被片边缘波皱程度较强。
3 讨论
随着我国大力推广生态型、节约型的园林绿化理念,宿根花卉在园林中发挥着越来越重要的作用[10],萱草以行植、丛植、群植的配置方式大量栽植于花境、花坛中。重瓣萱草因其独特花型具有较高的观赏性,深受人们的喜爱,但我国对萱草的
研究仍处于初级阶段,对重瓣萱草的研究更少,对其进行分类始终存在困难,进而无法充分掌握萱草的遗传信息,影响了育种工作的进一步开展[11]。同时,我国重瓣萱草杂交所依赖的种质基础薄弱,萱草种质资源的遗传基础较为狭窄,并且常出现杂交不亲和现象,使得难以选育出有突破性的新品种。我国学者通过分子标记的方法对萱草遗传多样性研究较少,对重瓣萱草的研究更是寥寥无几,并且前人对萱草的分子标记研究主要是利用EST-SSR[12]、ISSR[13]等标记。本研究利用SSR分子标记对34个重瓣萱草品种的遗传多样性进行研究,共筛选出PCR相对稳定、重复性好、可扩增出预期大小片段的12对引物,且它们都具有较高的多态性(PIC>0.5),这12对引物中共扩增出604条条带,多态性百分率达100%。对比任阳[14]使用ISSR分子标记对50个萱草属植物的研究中共扩增出196条条带,多态性百分率为96.43%;彭泽宇[15]使用SRAP分子标记对13个萱草品种的研究表明,多态性百分率为90.1%,本研究的多态性百分率均比其高。本实验采用荧光毛细管电泳法可以准确区分出34个重瓣萱草品种,克服了聚苯烯酰胺凝胶电泳法中人工对SSR片段辨别产生的误差[16],说明荧光毛细管电泳法的多态性和开发效率均较高,可有效地解释材料间的遗传差异,同时,也印证了该方法可以对不同重瓣萱草的遗传多样性水平进行鉴定和研究。
随着科技水平的不断提高,SSR分子标记被广泛应用于遗传多样性的研究中,研究的成功与否取决于选用的分子标记的基因型数和质量的准确性[17]。朱华芳等[18]利用5对SSR标记对6个萱草原种或变种和14个萱草园艺品种进行了PCR扩增,共扩增出23条条带,扩增片段大小在600~100 bp之间。本研究利用12个SSR标记对34个重瓣萱草品种进行检测,共获得了111个等位基因,平均9.2500个,遗传多样性平均值为0.8043,多态性信息含量(PIC)平均值为0.7875,Shannon信息指数(I)平均值为1.8753,表明各供试材料间遗传多样性水平较高;与此同时,平均观测杂合度明显低于期望杂合度(表3),说明这34个重瓣萱草品种纯合子合体较多。本实验数据为重瓣萱草品种的功能基因定位奠定了一定的理论基础,同时也为之后的研究提供了纯度鉴定依据。
罗军武等[19]研究认为,彼此间亲缘关系的远近可以通过遗传距离的大小反映出来。戴李川[20]研究表明,遗传距离越大,遗传多样性越高,遗传基础越广泛。本研究实验数据表明,34个重瓣萱草品种的遗传相似系数范围为0.225~0.413,在聚类分析图中,类群I中的4个重瓣萱草品种与类群Ⅱ中的30个重瓣萱草品种亲缘关系相对较近;28号‘15087-3’、29号‘15087-5’最早聚在一起,说明这2种重瓣萱草的亲缘关系极其接近。重瓣萱草在遗传进化与育种中基因库较为局限,遗传基础比较狭窄,有可能是因为在育种时选择的亲本较为接近。因此,在遗传相似系数较低的重瓣萱草品种中应加快发掘优异的基因资源,从而推进重瓣萱草品种的分子育种研究,以期为今后培育更多的重瓣萱草新品种提供参考和依据。
猜你喜欢
科教新报(2022年24期)2022-07-08
中国典型病例大全(2022年9期)2022-04-19
大众健康(2021年7期)2021-07-28
食品与生活(2020年8期)2020-09-12
科学导报(2020年39期)2020-06-21
绿色科技(2019年14期)2019-11-19
江苏农业科学(2019年5期)2019-09-02
江苏农业科学(2017年17期)2017-11-15
医学信息(2017年1期)2017-02-28
小说评论(2014年3期)2014-11-14
