
上海水源地毗邻湖库浮游植物群落结构的季节变化及其影响因子*
2022-06-30 07:06王三秀黄清辉
湖泊科学 2022年4期
王三秀,魏 莱,王 爽,陈 玲,黄清辉
(同济大学环境科学与工程学院长江水环境教育部重点实验室,上海 200092)
湖泊和水库通常充当着周边居民的生活饮用水水源地.然而,受自然环境变化及人类活动等因素的影响,部分湖库水生态系统受损严重[1].富营养化和水华频发是许多湖库面临的严峻挑战,特别是在长江中下游地区浅水湖泊,由人类活动影响及全球气候变暖的叠加效应所致[2-5].水华事件可引起水体缺氧或释放藻毒素,导致水生动物死亡,并威胁人类饮水安全与身体健康[6].浮游植物作为水生态系统中的初级生产者[7],在物质循环和能量传递中发挥重要作用,对水环境有不同的敏感响应和特异适应性,常用作水生生态系统健康状况的监测生物与评估指标[8-9].因此,研究水体浮游植物的群落结构变化规律及主要环境影响因子对水华的防治具有重要意义.
浮游植物的群落结构变化规律已有大量研究报道.研究表明,长三角地区太湖[10]、巢湖[11]、滆湖[12]、太平湖[13]、淀山湖[14]等湖泊浮游植物群落结构通常呈现出季节性演替的规律,且大多数表现出“夏、秋季相似,冬、春季相似”的特征.水温是影响浮游植物群落生物量、丰度或多样性指数的季节分布的关键因素之一;此外,也受透明度、总磷、总氮、溶解性总磷、溶解性总氮或氨氮浓度等水质参数的影响,但在不同湖泊和季节存在差异.例如,太湖夏、秋季优势功能群分布主要受到透明度的影响,春季优势功能群分布主要受到总氮浓度的影响[10];巢湖全年较优势的浮游植物功能群中以长孢藻为主,但在夏秋季时优势功能群为M(以微囊藻为主),其分布格局主要受水温、溶解性总磷、溶解性总氮、高锰酸盐指数和水深等影响[11].淀山湖冬季浮游藻类群落(以硅藻和绿藻为优势种群)生物量与总磷浓度显著相关,夏季(蓝绿藻种群占优势)则与氨氮浓度显著相关,但在春(以绿藻为主)、秋季(以蓝藻为主)与营养盐浓度的相关性不显著[14].
青草沙水库与金泽水库是上海市两大主要饮用水水源地,向全市供水量约80%,受长江和东太湖来水中氮磷的影响[15-16],库区将面临富营养化、藻华及异味等问题,进而可能危及供水安全[17].中央沙水域紧邻青草沙水库作为备用水源,筑堤围堰蓄水后,水闸常年关闭,与长江换水较弱,处于较封闭的状态.南白荡属于开放型湖荡,东部与金泽水库设有一个应急连通涵闸备用,北部与南部分别与葑漾荡和火泽荡连通,周围有居民分布,土地利用类型主要为耕地,受人类活动影响较大.张家路[18]曾报道了中央沙水域成库停止运行初期的浮游植物群落主要以绿藻和硅藻为主,细胞密度的季节变化范围为6.85×105~1.24×107cells/L,目前,尚未见有关南白荡的相关研究报道,中央沙水域建成多年后环境条件逐渐趋于稳定,浮游植物群落结构变化特征及其与水文水质的关系如何至今尚不明确.
本文旨在通过比较研究,分析中央沙与南白荡水域在不同的开放程度及周围土地利用差异影响条件下的浮游植物群落结构组成和变化,探讨驱动2个潜在水源地浮游植物群落分布的主要环境因子,以期为青草沙水库和金泽水库的水质管理提供参照样本及长三角地区湖库分类管理提供基础数据和科学依据.
1 研究与方法
1.1 研究区域概况
由河口滩涂湿地圈围成库的中央沙水域(31°26′34″~31°29′35″N, 121°30′31″~121°34′40″E),位于长江口南港和北港分流处,沙洲呈西北-东南走向,北邻青草沙水库,东邻长兴岛,库区面积为14.28 km2,库容约为1.03×108m3,2007-2014年曾为长兴岛集约化地表水厂供水(约4×104m3/d),2015年后水厂一直从青草沙水库取水[19].中央沙水域滩涂有水草和芦苇等植被分布,水域内包括众多的串沟和港汊,其中宽度200 m以上的大型港汊有2条,与外界水体交换能力较弱.
南白荡(31°1′40″~31°2′27″N, 120°56′0″~121°56′28″E)是太浦河北岸紧邻金泽水库[16]的一个开放型湖荡,位于金泽水源地二级保护区,西至西白荡,南至火泽荡,东至金泽水库,北至葑漾荡,湖泊面积约0.38 km2,平均水深约3 m,水源主要来北部的自葑漾荡,最终经火泽荡汇入太浦河[20],与外界水体交换能力强.南白荡周围有村落分布,土地利用类型主要为耕地,南部水域大型水生植物茂盛.
1.2 样品采集与处理
在中央沙水域汊道及沿岸总共布设6个采样点(编号为S1~S6),南白荡沿岸出入湖口以及周边相邻河道布设8个采样点(编号为P1~P8),中央水域S2、S5、P2、P3和P8样点采用无人船采样(图1).于2019年11月-2020年11月,每月一次进行了浮游植物采集和水质调查(新冠疫情暴发初期2020年2-4月除外),涵盖春(2020年5月)、夏(2020年6-8月)、秋(2019年11月、2020年9-11月)、冬(2019年12月、2020年1月)季节.

图1 中央沙水域与南白荡采样点分布
浮游植物定量采集:取离水面约0.5 m深处水样,用容量为1 L的聚乙烯瓶盛装,立即加入15 mL鲁哥试剂进行固定;定性采集:采用孔径为0.064 mm的25号浮游植物网在表层水面以“∞”字形拖动约3 min,再用蒸馏水冲洗附着在网上的浮游植物,收集10~20 mL样品装入聚乙烯瓶,用4%福尔马林溶液进行现场固定.水样在冷藏避光条件运回实验室,将1 L水样静置沉淀48 h,采用虹吸法吸去上层清液,样品浓缩定容至50 mL保存[21-22].浮游植物的计数和鉴定在显微镜下完成,主要根据王全喜[23-24]和胡鸿钧等[25]的文献进行所有浮游植物的镜检工作.
水温、pH、浊度(TUR)、溶解氧(DO)、电导率(EC)及总溶解盐(TDS)等指标利用多参数水质监测仪YSI EXO3现场测定,水体透明度用赛氏盘测定;采集部分水样带回实验室分析,参考《水和废水监测分析方法(第四版)》[26]进行样品指标浓度检测:总磷(TP)浓度采用钼酸铵分光光度法测定,总氮(TN)浓度采用碱性过硫酸钾消减紫外分光光度法测定,氨氮(NH3-N)浓度采用纳氏试剂分光光度法测定,高锰酸盐指数(CODMn)采用高锰酸钾消解滴定法测定;溶解性有机碳(DOC)浓度采用总有机碳分析仪(岛津TOC-VCPN,日本)分析;叶绿素a(Chl.a)浓度用丙酮萃取分光光度法测定[27].
1.3 数据处理与分析
以叶绿素a为基准参数,选取TP、TN、SD和CODMn参数进行相关加权综合,采用加权综合营养状态指数(TLI(∑))评价法对水体营养状态等级进行评估[28].根据Mcnaughton[29]优势度指数(Y)≥ 0.02确定浮游植物的优势属.筛选出生物量贡献大于1%或在各采样点出现频率大于60%的优势浮游植物,运用Canoco5.0软件对物种与环境因子进行排序分析[30-32],软件分析时将物种和环境因子均进行lg(x+1)转换(pH除外)[33],对物种进行去趋势分析(DCA)得到排序轴最大长度分别为1.4、3.3,均可选用冗余分析.
本研究所有数据统计分析和绘图用Excel和Origin 2018完成,采样点位分布图及浮游植物细胞密度的空间分布采用ArcGIS绘制,在IBM SPSS Statistics 22软件中进行数据相关性分析.
2 结果与分析
2.1 水体理化因子特征
中央沙水域和南白荡的水体理化指标用平均值±标准偏差表示(表1).中央沙水域与南白荡的水温季节变化一致,变化范围为9.50~29.5℃,水体均呈弱碱性;中央沙水域的透明度、TDS和EC低于南白荡,浊度与透明度呈相反变化的趋势,TDS与EC变化趋势基本一致;2个水体的DO浓度均表现为春冬季较高,夏秋季较低;中央沙水域的NH3-N浓度在0.029~0.191 mg/L之间,夏季显著高于其他季节,TN浓度在0.683~0.866 mg/L之间,根据《地表水环境质量标准》(GB 3838-2002),86%以上的样品满足Ⅲ类水质,南白荡的NH3-N浓度介于0.030~0.235 mg/L之间,夏季显著高于其他季节,TN浓度介于0.460~2.020 mg/L之间,冬季显著高于其他季节,62%以上的样品满足Ⅲ类水质,南白荡的NH3-N和TN浓度略高于中央沙水域;中央沙水域、南白荡TP浓度的变化范围分别为0.079~0.163、0.068~0.123 mg/L,大部分样品处于Ⅳ~Ⅴ类水质之间;中央沙水域的CODMn、DOC和Chl.a浓度均比南白荡高.中央沙水域的氨氮、总氮绝大多数满足Ⅲ类及更优水质要求,高锰酸盐指数和总磷较难满足Ⅲ类水质要求,南白荡的氨氮、高锰酸盐指数绝大多数满足Ⅲ类水质要求,总氮和总磷较难满足Ⅲ类水质要求.
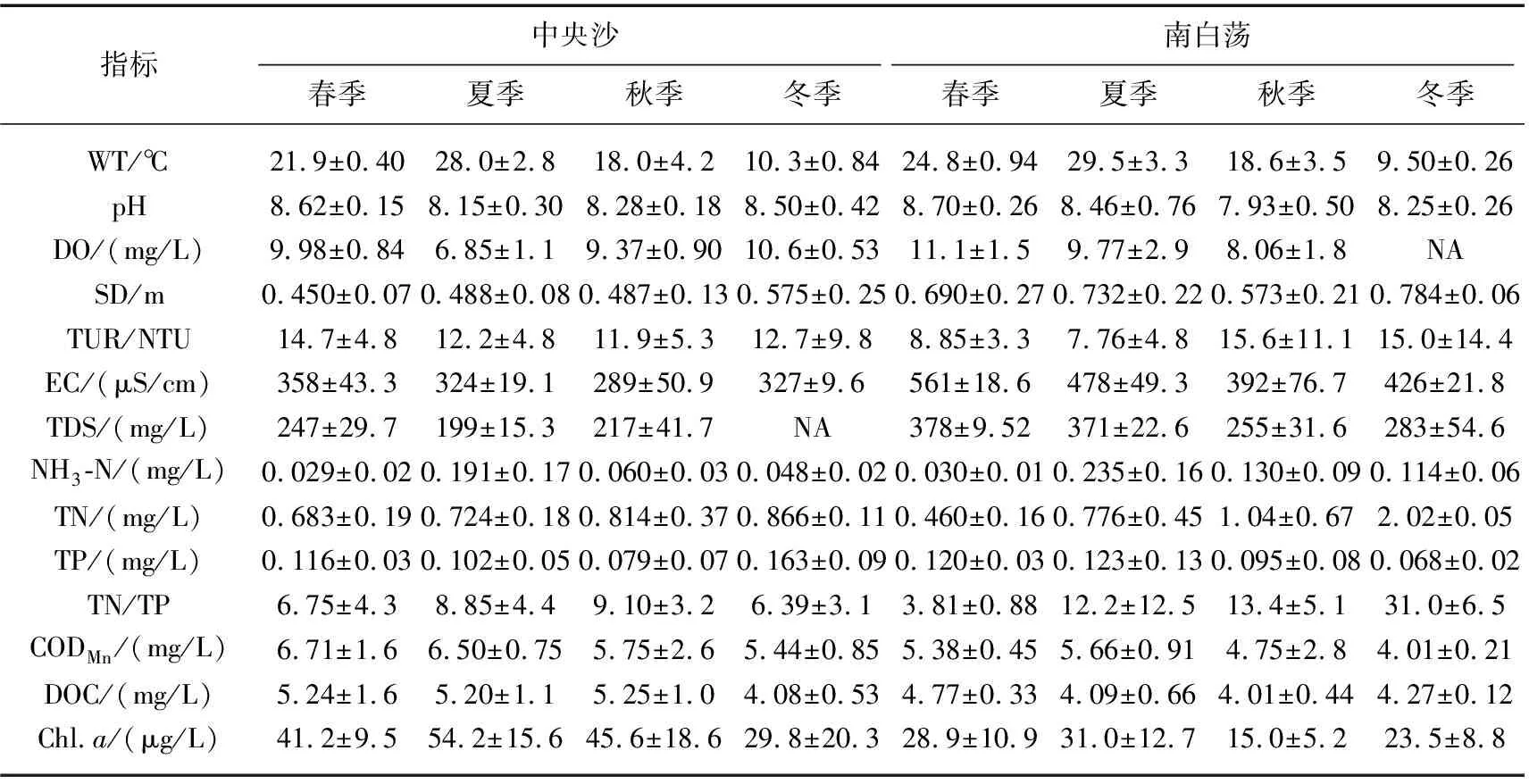
表1 中央沙水域与南白荡水体理化因子的季节平均值与标准偏差值*
2.2 综合营养状态评价
中央沙水域和南白荡水体的季节平均TLI(∑)分别介于57.5~59.0、54.1~56.1之间,全年均处于轻度富营养状态,季节变化不显著,中央沙水域的营养状态要高于南白荡(图2).
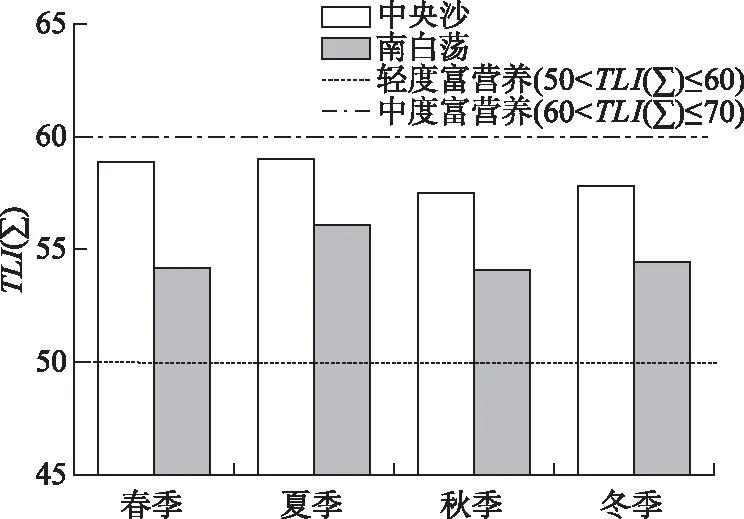
图2 中央沙水域和南白荡综合营养状态指数的季节变化
2.3 浮游植物的群落结构特征
2.3.1 浮游植物种类组成 在中央沙水域检出浮游植物7门104属184种(鉴定到属的按1个种计算,下同).其中,蓝藻门有24属35种,占总种数的19.0%;硅藻门有23属37种,占总种数的20.1%;绿藻门有41属85种,占总种数的46.2%;裸藻门、隐藻门、甲藻门、金藻门共占总种数的14.7%.
在南白荡检出浮游植物8门96属172种.其中,蓝藻门有20属34种,占总种数的19.8%;硅藻门有20属39种,占总种数的22.7%;绿藻门有39属73种,占总种数的42.4%;裸藻门、隐藻门、甲藻门、金藻门和黄藻门共占总种数的15.1%.南白荡与中央沙水域浮游植物种类组成相似,绿藻门的种类数最多,其次是硅藻门和蓝藻门,甲藻门、隐藻门、裸藻门和金藻门的种类数相对较少,黄藻门仅在南白荡检出.
2.3.2 中央沙浮游植物细胞密度及时空分布 中央沙水域浮游植物细胞密度各季节平均值变化范围为 3.00×107~1.61×108cells/L(图3a).蓝藻细胞密度最大,季节平均细胞密度在2.25×107~1.24×108cells/L之间,占总细胞密度的82.8%;其次为硅藻和绿藻,占比分别为7.99%、7.21%;隐藻、裸藻、金藻和甲藻总共占比2.00%,浮游植物细胞密度表现为夏季>秋季>春季>冬季.浮游植物细胞密度组成在时空上具有一定差异,蓝藻细胞密度在各季节占比均最大,是构成中央沙水库浮游植物的绝对优势类群,绿藻细胞密度在夏秋季显著高于其他季节,金藻主要出现在冬季(图3b).
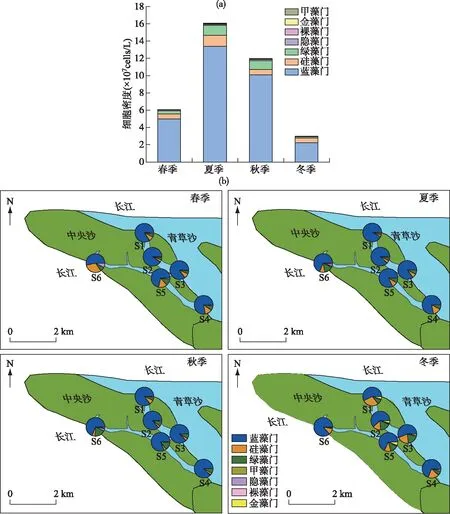
图3 中央沙水域浮游植物细胞密度的季节变化及其时空分布
2.3.3 南白荡浮游植物细胞密度及时空分布 南白荡浮游植物细胞密度季节平均值变化范围为4.29×106~6.59×107cells/L(图4a).蓝藻细胞密度在夏秋季显著增加,变化范围为2.56×105~4.91×107cells/L,占总细胞密度的71.2%;其次为硅藻、隐藻和绿藻,占比分别为11.3%、9.19%、7.16%;裸藻、金藻、甲藻和黄藻总共占比1.15%,浮游植物细胞密度也表现为夏季>秋季>春季>冬季.如图4b所示,南白荡浮游植物群落结构发生了显著季节性演替,春季以硅藻和隐藻为主,蓝藻和绿藻次之;夏季以蓝藻为主,硅藻、隐藻和绿藻次之;秋季以蓝藻为主,隐藻占比有所增加;冬季以隐藻为主,其次是硅藻、金藻和绿藻.总体上,南白荡的浮游植物春冬季以隐藻和硅藻为主,夏秋季以蓝藻为主,各样点浮游植物密度组成存在显著差异.

图4 南白荡浮游植物细胞密度的季节变化及时空分布
2.4 浮游植物优势类群分析
中央沙水域共鉴定出浮游植物优势类群4门17属(表2).全年以蓝藻门的优势类群为主,假鱼腥藻属是全年优势类群,长孢藻属和束丝藻属是春、夏、秋季优势类群,微囊藻属是秋季优势类群;硅藻门的直链藻属和小环藻属主要出现在春冬季节,针杆藻属主要出现在冬季;绿藻门的十字藻属和单针藻属分别出现在秋季和冬季;隐藻门的隐藻属和蓝隐藻属主要出现在春季.总体上,蓝藻门的假鱼腥藻属和长孢藻属是主要优势类群,假鱼腥藻属是全年的绝对优势类群(Y≥ 0.1),硅藻门和隐藻门的优势属主要出现在春冬季.
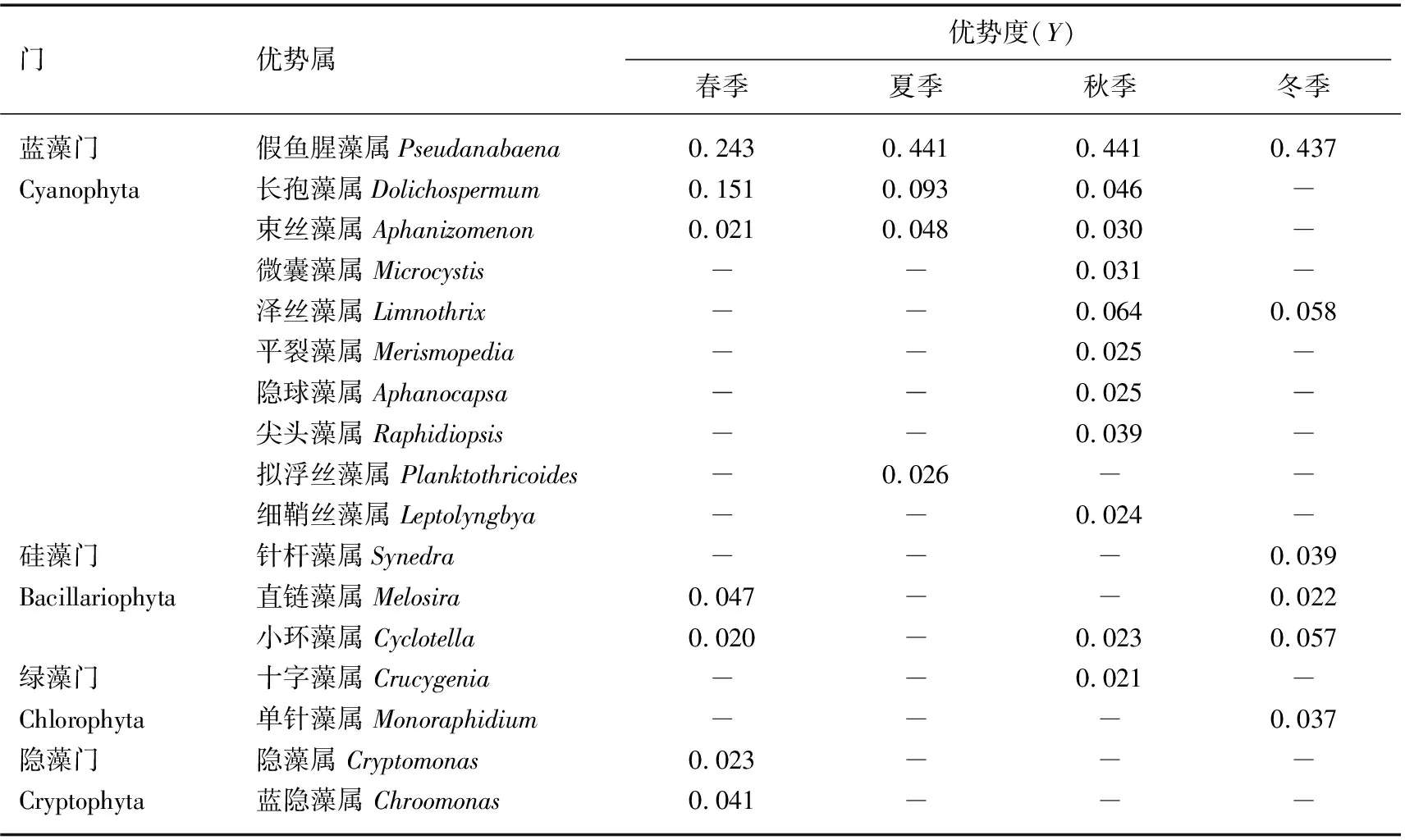
表2 中央沙水域浮游植物优势属及优势度*
南白荡浮共鉴定出浮游游植物优势类群5门13属(表3).春季硅藻门的小环藻属和隐藻门的隐藻属、蓝隐藻属的优势度较大,为主要优势种;夏季主要优势类群为蓝藻门的假鱼腥藻属和平裂藻属;秋季南白荡P1点水面出现明显的水华现象,优势类群主要为微囊藻属;冬季水体的主要优势类群为蓝隐藻属.假鱼腥藻属、直链藻属、隐藻属和蓝隐藻属在四季都是优势属,春冬季优势类群主要为隐藻门和硅藻门,夏秋季为蓝藻门.
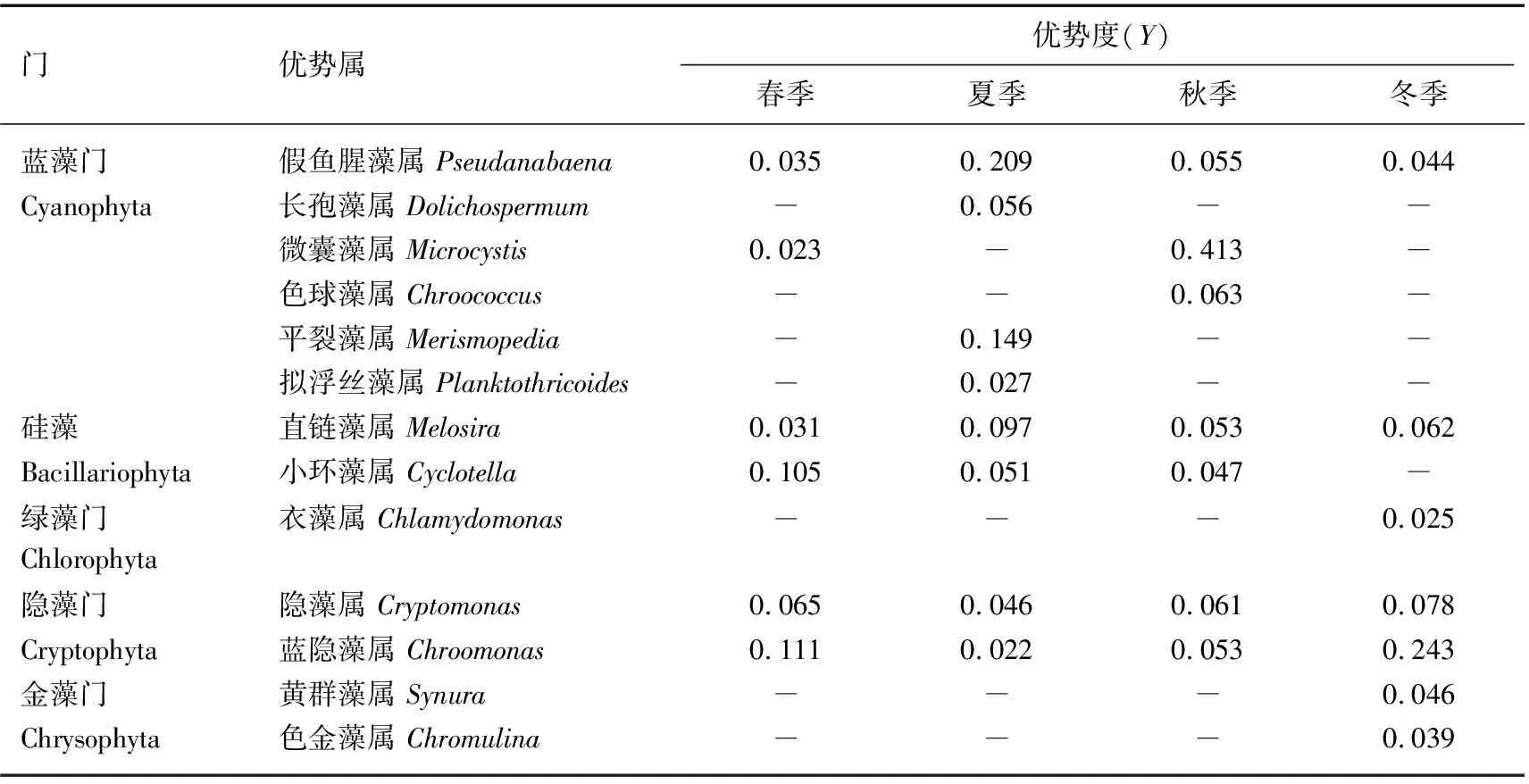
表3 南白荡浮游植物优势属及优势度*
2.5 影响浮游植物的环境因子主成分分析
PCA分析结果显示(图5),8个环境因子变量在前4轴上解释了82.9%的数据变异性(轴1=31.3%,轴2=25.8%).决定轴1主要的环境因子为TN(0.50)、WT(0.48)、NH3-N(0.32)和TDS(0.30),轴2的主要环境因子为pH(0.55)和TP(0.55).PCA得分结果表明,样本主要按照季节和不同水体进行聚类,第1、2主成分主要描述了研究区域的季节变化和营养盐浓度2个主要的环境变化.从季节变化来看,同一采样点的样品表现出一定的分离,水质变化显著,夏季影响中央沙水域的主要环境因子为WT和NH3-N,可能与库区内大型植物生长吸收营养盐有关,其余季节各环境因子对库区水质影响较均匀,水体环境较稳定,而南白荡各季节水质变化显著,水体环境稳定性较弱,秋季水域内暴发了大量的水葫芦、野菱等大型水生植物,吸收了较多的营养盐浓度,夏季和冬季主要环境影响因子分别为TP和TN.
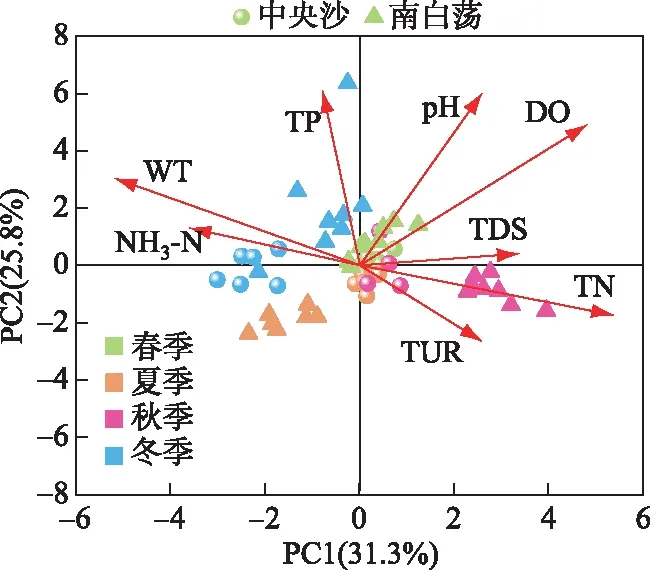
图5 中央沙和南白荡水域环境因子的PCA分析
2.6 浮游植物种属与环境因子的冗余分析
中央沙水域的RDA分析结果如图6a所示,前2个排序轴物种与环境因子的相关关系分别达到0.8242和0.8337,前4轴共解释了86.5%的环境因子信息,前2轴占63.7%,所有特征值解释了浮游植物群落变异程度的46.7%.第1排序轴与TN呈显著正相关(r=0.58,P<0.01),与TP(r=-0.49,P<0.05)、CODMn(r=-0.48,P<0.05)呈负相关;第2排序轴与WT(r=0.57,P<0.05)呈负相关,与DO呈正相关.因此,总氮、总磷、水温可被认为是影响中央沙水域浮游植物群落分布的主要环境因子,而浮游植物生长繁殖也可影响水体中DO的浓度.
南白荡的RDA分析结果如图6b所示,前2个排序轴物种与环境因子的相关关系达到0.8400和0.8740,前4轴共解释了92.5%的环境因子信息,前2轴占78.1%,所有特征值解释了浮游植物群落变异程度的55.5%.第1排序轴与WT、DO和pH呈正相关(r=0.61、0.51、0.48,P<0.01);第2排序轴与TDS呈负相关(r=-0.73,P<0.01).因此,水温、总溶解性盐可被认为是影响南白荡浮游植物群落分布的主要环境因子,而其浮游植物生长繁殖也可导致DO和pH值的变化.
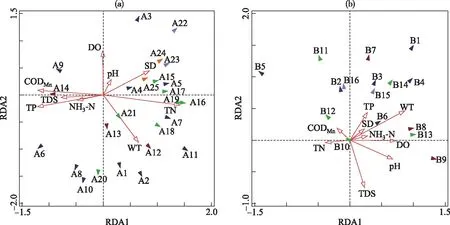
图6 中央沙水域(a)和南白荡(b)浮游植物与环境因子的RDA分析(A1、B1=假鱼腥藻属,A2、B2=长孢藻属,A3=隐球藻属,B3=色球藻属,A4、B4=平裂藻属,A5、B5=微囊藻属,A6=浮鞘丝藻属,A7=拟浮丝藻属,A8=浮丝藻属,A9=泽丝藻属,A10、B6=束丝藻属,A11=尖头藻属,A12、B7=针杆藻属,A13、B8=直链藻属,A14、B9=小环藻属,A15、B10=栅藻属,A16、B11=小球藻属,A17、B12=十字藻属,B13=衣藻属,B14=集星藻属,A18=单针藻属,A19=四角藻属,A20=转板藻属,A21=盘星藻属,A22、B15=隐藻属,A23、B16=蓝隐藻属,A24=裸藻属,A25=囊裸藻属)
3 讨论
3.1 浮游植物群落结构的季节演替规律
中央沙与南白荡水域浮游植物群落结构季节性演替存在显著差异,中央沙水域浮游植物群落全年以蓝藻为主,主要优势类群为假鱼腥藻属、长孢藻属,而南白荡浮游植物群落季节性演替规律为:春季以小环藻属、隐藻属和蓝隐藻属为主,夏季以假鱼腥藻属和平裂藻属为主,秋季以微囊藻属为主,冬季以蓝隐藻属为主,这些浮游植物优势类群均属于富营养化水体指示种[25,34],表明中央沙水域与南白荡水体均处于富营养化状态,这与图2所示的综合营养状态指数评价结果一致.
中央沙水域成库后停止运行初期浮游植物群落结构组成主要为绿藻和硅藻[18],近年来库区水域的封闭状态使水体环境条件趋于稳定,调查期间浮游植物群落结构演替成全年以蓝藻为主的绝对优势类群.与毗邻的青草沙水库有着相似的演替规律,青草沙水库建成初期,绿藻种数最多,蓝藻为库区的绝对优势类群[35].青草沙水库正式运行后,成为半开放型水体,浮游植物群落发生了明显的季节性演替:春冬季以硅藻、隐藻和黄藻为主,夏秋季以蓝藻、硅藻、绿藻为主[18,36-37].Oliver等[38]有关研究表明,在亚热带地区温度和光照随季节变化不明显,若水体中具有较高浓度的氮和磷,则蓝藻可能成为全年的优势藻种,对于青草沙水库及其备用的中央沙水域,长江口径流输入以及库区湿地植被释放的营养盐导致库区富营养化程度严重[39],有利于蓝藻生长.研究表明,多数蓝藻适宜在较稳定的水体中生长,而绿藻和硅藻则更适应紊乱的水体条件,且流动水体中的浮游植物密度要低于静水水体[40].中央沙筑堤围堰蓄水多年后,水流速度减缓,水力停留时间长,淹没的植被形成腐殖质进入沉积物中,导致水体环境恶化,自净能力减弱[41-42],较稳定水环境条件形成了常年以假鱼腥藻属、长孢藻属和束丝藻属等为主的优势类群,这些都是产毒、产嗅的重要藻源,对饮用水安全产生了巨大威胁.南白荡浮游植物群落演替规律与太湖[10]、太平湖[13]、淀山湖[14]等长三角地区湖泊的季节演替规律相似,夏秋季以蓝藻为主要优势类群,春冬季节以硅藻和隐藻为优势类群,金藻主要出现在冬季.南白荡属于开放型水域,水流自北向南汇入太浦河,水力停留时间短,常年与外界水体进行交换不仅改变了南白荡水体的理化因子特征,同时还会引进新的浮游植物,难以形成常年稳定的浮游植物优势类群[41].因此,中央沙与南白荡水域的封闭或开放特征引起的水文水质条件差异可能是造成2个备用水源地季节性浮游植物群落演替差异的重要原因之一.
3.2 浮游植物群落结构与环境因子的关系
浮游植物群落在水生态系统中生长和繁殖的主要影响因子包括水温、光照、营养盐浓度、盐度、水体动力等非生物因素[43-45],浮游动物捕食[46]等生物因素,及地理空间和土地利用类型[47-48].RDA分析结果表明,水温是共同影响南白荡和中央沙水域浮游植物群落分布的显著因素,与众多湖库研究一致.水温可以通过影响浮游植物的生理和代谢过程直接影响浮游植物,也可以通过影响浮游植物生长的水生环境间接影响浮游植物[49-50].夏季2个水体温度均值可达28℃(表1),细胞分裂速率加倍,适宜高温生长的蓝藻、绿藻迅速繁殖[51],浮游植物细胞密度显著增加,春冬季温度降低,喜低温的硅藻、金藻和黄藻占比增加,隐藻属、蓝隐藻属和小环藻属成为南白荡的主要优势类群.近年来全球气候变暖,为蓝藻的生长提供了有利的环境条件[52],中央沙水域蓝藻的绝对竞争优势抑制了其余藻类的生长,假鱼腥藻属在2个备用水源地中四季均是优势类群,与温度呈显著正相关,夏季繁殖成为库区主要优势类群.
氮和磷是浮游植物生长繁殖所必需的营养物质,与浮游植物多样性、丰度和生物量等息息相关.RDA结果显示,TN和TP是影响中央沙水域浮游植物群落分布的主要环境因子,这与张家路[18]、严文逸等[37]在邻近的青草沙水库的研究结果一致.多数浮游植物优势类群与TN呈正相关,而小环藻属、浮鞘丝藻属、泽丝藻属、浮丝藻属、束丝藻属和转板藻属与TP呈正相关,氮/磷比可以反映营养盐对浮游植物的限制,中央沙水域TN/TP比值全年小于16,说明中央沙水域浮游植物生长可能受到氮限制,长孢藻属、束丝藻属等固氮蓝藻生长占优势[53];而南白荡仅在冬季TN/TP比值大于16,浮游植物生长受到磷限制[54-56],与岸边的农耕作业及生产养殖活动减弱有关.南白荡筛选出的浮游植物均与TP呈正相关,微囊藻属、长孢藻属、十字藻属、栅藻属、小球藻属和蓝隐藻属与TN呈正相关,这些藻属在中央沙水域也与TN呈正相关,表明营养盐在不同水体对浮游植物群落影响的异同性.PCA分析结果显示(图5),中央沙水域各季水质较稳定,但夏季因降雨量较大而有波动,而南白荡水体环境因子季节变化显著,与周围居民的生产活动及外来输入水源有关,外源氮磷污染物通过生活污水排放或降雨冲刷耕地等途径进入湖泊,致使水域内营养盐浓度较高,但不是浮游植物群落生长的主要驱动因子.有研究表明,在水体流动较强时,营养盐浓度不再是影响浮游植物群落结构分布的主要驱动因素,而是水动力条件,水体流通性可能是造成驱动2个备用水源地浮游植物群落分布的主要环境因子存在差异的关键因素[57].南白荡的优势类群直链藻属、小环藻属有较重的硅质外壳,在湖泊水体流动期间生长占据优势[58].此外,小环藻属、直链藻属和衣藻属与pH呈显著正相关,在弱碱性水体中浮游植物更容易吸收空气中的CO2,加速自身光合作用和水生系统的能量传递与物质转化[59],浮游植物生长同时也消耗了水体中的CO2导致pH值升高,并释放出O2.因此pH与DO变化也可能由大量繁殖的浮游植物所致,本研究不作为影响浮游植物生长的关键环境因子.
中央沙与南白荡水域不同的开放程度及周围土地利用类型的差异,导致2个备用水源地浮游植物群落结构分布的环境影响因子存在显著差异.总体来说,营养盐浓度、水温和水体流通性等因素是影响浮游植物生长的重要因素,针对中央沙水域可重点控制水体中氮、磷浓度,开放库区闸门,让水体流动起来.南白荡可通过促进大型水生植被生长,吸收水体中的养分,降低光照条件,抑制浮游植物过度繁殖[60],同时控制周围耕地农肥使用及生活污水的排放,减少外源营养盐过量输入.
4 结论
中央沙与南白荡水域水质整体均处于轻度富营养状态,浮游植物细胞密度呈一致的季节性变化,而群落结构季节性演替规律存在显著差异,中央沙水域封闭的水域状态及较高的营养盐浓度为蓝藻的生长提供了稳定的水环境条件和营养物质,库区全年蓝藻占绝对优势,与之前的研究相比,库区浮游植物群落结构的组成发生显著变化.而南白荡开放的水体与外界水质交换频繁,浮游植物群落结构季节性演替明显,春冬季以隐藻和硅藻为主,夏秋季以蓝藻为主.
中央沙水域浮游植物群落分布主要受总氮、总磷和水温等因素控制,而南白荡则主要受水温和总溶解性盐影响,湖库水质管理不仅要关注水体中的营养盐浓度,还要关注水体流通性,筑堤围堰蓄水后,需要经常开放闸门与外界进行水质交换,增加水域内的水体流通性.蓝藻细胞密度过高是上海典型湖库面临的共同挑战,管理者应根据影响浮游植物群落结构分布的不同环境因子,分别采取适宜的措施进行水质管理.
猜你喜欢
当代水产(2022年6期)2022-06-29
发明与创新·中学生(2022年3期)2022-03-14
农业科技与信息(2020年14期)2020-12-18
流行色(2019年10期)2019-12-06
绿色科技(2019年14期)2019-11-19
装饰装修天地(2019年9期)2019-10-21
江苏农业科学(2019年5期)2019-09-02
绿色科技(2018年19期)2018-11-29
海洋世界(2016年2期)2016-03-11
科学24小时(2015年3期)2015-09-10
