
玉米大斑病菌HST生物合成基因簇的结构和表达分析
2022-07-12 14:01张淑红高凤菊武秋颖张运峰范永山
西北农林科技大学学报(自然科学版) 2022年7期
张淑红,高凤菊,武秋颖,张运峰,范永山
(唐山师范学院 生命科学系 唐山市农业病原真菌与毒素重点实验室,河北 唐山 063000)
玉米大斑病是一种严重威胁玉米生产的真菌性病害,在全球温湿地域的玉米产区均有发生,发生严重年份感病品种减产可达50%左右[1]。生产上玉米大斑病主要依赖抗病基因育种防治,目前应用的抗病基因主要是受单基因控制的Ht1、Ht2、Ht3和HtN,分别位于玉米染色体2.07,8.05,7.04和 8.06 区[2-5]。携带不同Ht基因的玉米基因型与玉米大斑病菌(Setosphaeriaturcica)不同生理小种之间存在特异性互作。寄主选择性是玉米大斑病菌与玉米之间特异性互作的决定因素。在前期研究中,Fan等[6]发现玉米大斑病的发生依赖于玉米大斑病菌产生的HT-毒素。HT-毒素是在玉米大斑病菌培养滤液中发现的一种对热稳定、非蛋白质结构的化合物[7]。研究发现,玉米大斑病菌不同生理小种间的致病性差异与HT-毒素不同毒性组分的生物学活性有关[8-10]。但是,对玉米大斑病菌HT-毒素中寄主选择性组分的化学结构和生物合成过程至今尚未明确[11]。
寄主选择性毒素(HST)是植物病原真菌产生的一类次生代谢产物,其对寄主种类或栽培品种具有特异的生理活性和高度专化性,是决定致病性的重要因素[12-13]。自1933年Tanaka等[14]从梨的菊池链格孢菌(Alternariakikuchiana)上发现第一个HST以来,已报道植物病原真菌13个属的27个种(或小种或专化型)都可以产生HST,其中16种HST的化学结构已经确定[15-17]。本研究从玉米大斑病菌次生代谢产物合成基因簇中,发现与交链链格孢(Alternariaalternata)寄主选择性毒素ACT-毒素合成酶基因高度同源的基因簇Cluster 397.3,通过比较Cluster 397.3基因簇组成基因在玉米大斑病菌接种不同时间和不同生理小种菌株间的表达差异,分析其在病菌寄主选择方面的可能功能和作用机制,以期为进一步明确玉米大斑病菌寄主选择性毒素的结构和功能奠定基础。
1 材料与方法
1.1 菌株和玉米自交系
玉米大斑病菌23号生理小种01-23菌株和1号生理小种01-11菌株,以及玉米感病自交系B37,均来自唐山市农业病原真菌与毒素重点实验室。
1.2 玉米大斑病菌Cluster 397.3的结构分析
利用antiSMASH技术[18]分析玉米大斑病菌23号生理小种et28a菌株的次生代谢产物合成基因簇,利用玉米大斑病菌基因组JGI数据库(https://mycocosm.jgi.doe.gov)进行Synteny共线性分析,获得HST生物合成基因簇Cluster 397.3,利用Blastp分析该基因簇的基因组成,利用antiSMASH技术的KnownClusterBlast(https://fungismash.secondarymetabolites.org)功能分析该基因簇与已知基因簇的同源性;利用MSA(multiple sequence alignment)分析推测HST生物合成基因StACTTS3的功能;利用MEGA 11.0软件进行系统进化分析,NCBI CDD(https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi)预测其结构域,利用TBtools软件(https://github.com/CJ-Chen/TBtools)可视化制图。
1.3 玉米大斑病菌Cluster 397.3的表达分析
将玉米大斑病菌23号生理小种01-23和1号生理小种01-11菌株在PDA平板培养基上28 ℃静置培养5 d,取直径为9 mm的菌盘,接种4~6叶期B37玉米叶片,以菌盘的菌丝面接触玉米叶片,保湿48 h后去除菌盘,按照常规方法进行栽培管理。分别于接种3,5,7,10 d,取接种部位叶片,在30 s内置于液氮中速冻,3 d内送交上海伟寰生物科技有限公司进行RNA提纯、反转录和高通量测序,制备RNA-Seq转录文库。为了保证分析数据的质量,去除接头污染Reads和低质量Reads后,得到质量较高的Clean Reads。从RNA-Seq转录文库中筛选HST生物合成基因簇基因,以β微管蛋白为内参,获得各基因的相对表达量。
1.4 数据处理与分析
利用prism软件[19]测算各基因的相对表达量并制图,对不同侵染时间和不同小种菌株间进行单因素ANOVA方差分析。
2 结果与分析
2.1 玉米大斑病菌HST生物合成基因簇的基因组成
利用antiSMASH技术分析玉米大斑病菌23号生理小种et28a菌株的基因组DNA,共发现48个次生代谢产物合成基因簇。利用玉米大斑病菌基因组JGI数据库与交链链格孢进行Synteny共线性分析,发现位于scaffold_9:1 695 105-1 735 028(39 924 bp)的Cluster 397.3基因簇与交链链格孢ACT-毒素合成基因簇(scaffold_21:760 959-776 667)有保守的共线性关系(图1-A、图1-C)。
Cluster 397.3基因簇含有9个基因(图1-A):1个氨基甲酰磷酸合成酶基因(carbamoyl phosphate synthetase,CPS)、2个细胞色素P450基因(cytochrome P450,P450)、1个短链脱氢酶/还原酶基因(short-chain dehydrogenase/reductase,SDR)、1个锌结合氧化还原酶基因(zinc binding dehydrogenase,ZOR)、2个耐药转运蛋白基因(drug resistance transporter,DRT)、1个聚酮合酶基因(polyketide synthase,PKS)和1个保守假定蛋白编码基因(conserved hypothetical protein,CHP)。
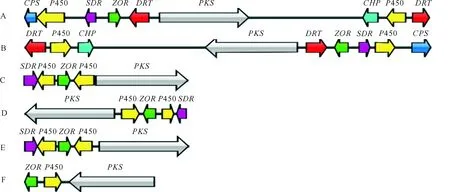
A.玉米大斑病菌et28a菌株;B.玉米大斑病菌ny001菌株;C.交链链格孢;D.芸薹生链格孢;E.壳二孢疫病病菌;F.玉米小斑病菌;CPS.氨基甲酰磷酸合成酶基因;P450.细胞色素P450基因;SDR.短链脱氢酶/还原酶基因;ZOR.锌结合氧化还原酶基因;DRT.耐药转运蛋白基因;PKS.聚酮合酶基因;CHP.保守假定蛋白编码基因
KnownClusterBlast(https://fungismash.secondarymetabolites.org)分析表明,Cluster 397.3基因簇与BGC0001264 Betaenone A/C生物合成基因簇有37%的相似性,而Betaenone A/C与链格孢(Alternariasp.)真菌产生的含EDA(epoxy-decatrienoic acid)结构的HST具有一定的结构相似性。推测Cluster 397.3基因簇也可能负责合成一种类似EDA结构的HST。
2.2 玉米大斑病菌Cluster 397.3基因簇的Synteny分析
Synteny共线性分析发现,玉米大斑病菌1号生理小种ny001菌株的Cluster 397.3同源基因簇scaffold_74:18 138―57 965也含有9个基因,与et28a菌株相同(图1-B)。
Synteny共线性分析得到交链链格孢、芸薹生链格孢(Alternariabrassicicola)、壳二孢疫病病菌(Ascochytarabiei)以及玉米小斑病菌(Cochliobolusheterostrophus)菌株的Cluster 397.3同源基因簇(图1-C-F)。其中,交链链格孢、芸薹生链格孢和壳二孢疫病病菌的Cluster 397.3同源基因簇中均包括玉米大斑病菌et28a菌株Cluster 397.3基因簇中的5个基因,分别为2个细胞色素P450基因(P450)、1个短链脱氢酶/还原酶基因(SDR)、1个聚酮合酶基因(PKS)和1个锌结合氧化还原酶基因(ZOR)。而玉米小斑病菌菌株的Cluster 397.3同源基因簇仅包括其中3个基因,与上述3种菌株相比,缺少1个细胞色素P450基因和1个短链脱氢酶/还原酶基因(图1-F)。
此外,在玉米大斑病菌基因组内还发现了交链链格孢ACTTS4、ACTT5和ACTT6的同源基因,其中玉米大斑病菌ACTTS4(StACTTS4)基因位于scaffold_4:334 954―338 493,编码的非核糖体肽合成酶具有α氨基乙二酸还原酶活性;StACTT5编码一种酰基辅酶A 合成酶(acyl-CoA synthetase);StACTT6编码一种烯酰辅酶A水合酶(enoyl-CoA hydratase);它们共同参与合成HST的EDA结构。
2.3 玉米大斑病菌Cluster 397.3 的结构分析
通过Blastp分析发现,玉米大斑病菌et28a菌株的PKS基因(scaffold_9:1 708 392-1 717 224)为ACTTS3同源基因,ACTTS3是ACT-毒素合成酶关键基因[15],ACTTS基因是链格孢菌橘致病型(Alternariaalternatatangerine pathotype)特有基因。ACTTS3编码一种聚酮合酶,玉米大斑病菌ACTTS3(StACTTS3)编码的聚酮合酶包括6个典型结构域,分别为聚酮合酶β-酮脂酰合成酶结构域(polyketide synthase,β-ketoacyl synthase domain,PksD;4-1 046)、聚酮合酶脱水酶(polyketide synthase dehydratase,PS-DH;945-1 249)、甲基转移酶-12(methyltransferase type 12,Methyltransf_12;1 427-1 528)、聚酮合酶酮还原酶(polyketide synthase,ketoreductase,PKS_KR;2 122-2 298)、脂肪酰基辅酶A还原酶NAD结合域(fatty acyl-coenzyme A reductase,NAD-binding domain,PKS_NbtC superfamily;2 126-2 477)、NAD结合Rossmann超家族(NAD-binding Rossman superfamily,NADB_Rossmann superfamily;2 583-2 870)。et28a和ny001菌株的聚酮合酶结构域排列无差异(图2)。
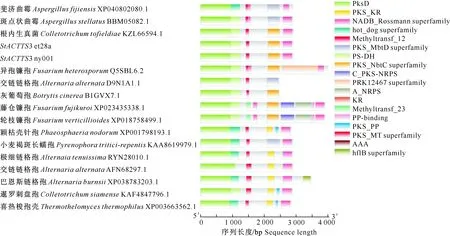
图2 玉米大斑病菌StACTTS3的结构域分析
系统进化分析结果(图3)表明,玉米大斑病菌StACTTS3与根内生真菌(Colletotrichumtofiel-diae)亲缘关系最近,其结构域分布完全相同。链格孢不同种之间以及同种不同菌株之间,StACTTS3编码蛋白的结构域种类和分布均不同,交链链格孢D9N1A1.1有4种结构域,交链链格孢AFN68297.1和极细链格孢菌RYN28010.1均有5种结构域,但结构域种类不同,而巴恩斯链格孢XP038783203.1有7种结构域(图2)。
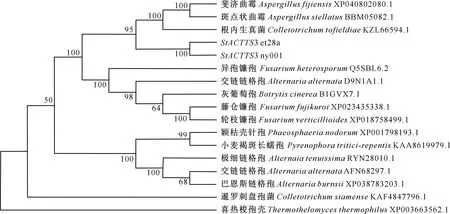
图3 玉米大斑病菌StACTTS3的系统进化树
2.4 玉米大斑病菌Cluster 397.3基因簇在病菌侵染过程中的表达分析
以玉米大斑病菌接种后的玉米叶片为样品进行转录组建库和测序,RNA-Seq转录文库数据显示共获得32 875 106~58 073 444 高质量Reads,碱基Q20(Phred数值大于20的碱基占总体碱基的比例)均大于96%,Q30(Phred数值大于30的碱基占总碱基数的比例)均大于90%,GC含量为51.67%~55.21%,表明转录组测序质量较好,可以进行下一步分析。玉米大斑病菌Cluster 397.3基因簇在病菌侵染过程中的表达分析结果见图4和图5。
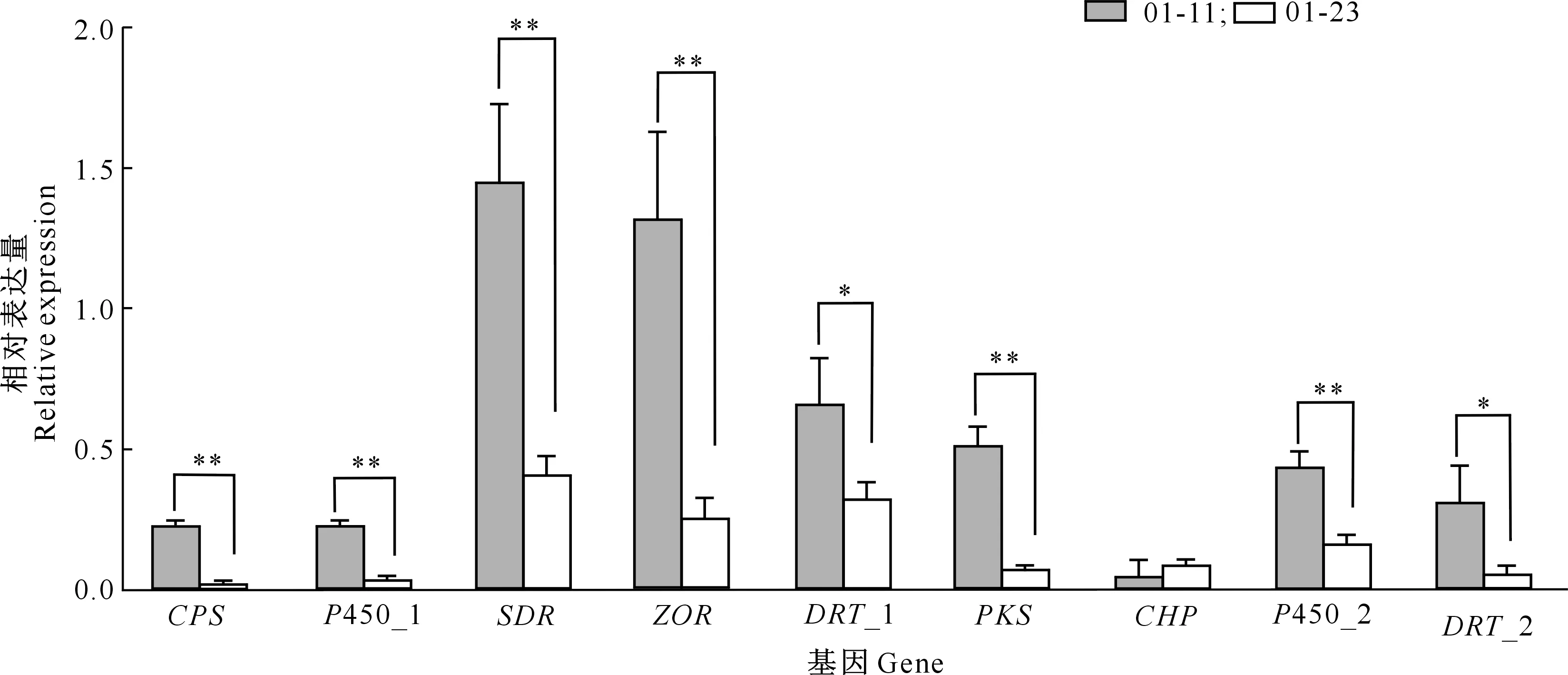
*、**分别表示01-11与01-23菌株间差异显著(P<0.05)和极显著(P<0.01)
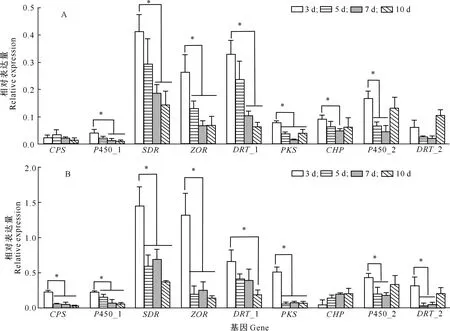
*表示接种后3 d的表达量与其他时间差异显著(P<0.05)
由图4和图5可知,在玉米大斑病菌接种感病玉米自交系B73叶片3,5,7,10 d后,Cluster 397.3基因簇基因均高效表达,但不同侵染时间和不同小种菌株间均存在差异。接种后第3天,Cluster 397.3基因簇的多数基因表达量显著高于其他侵染时间,表明Cluster 397.3基因簇主要在病菌侵染初期起作用,病菌侵入可能具有刺激毒素合成的作用;玉米大斑病菌1号生理小种菌株01-11与23号生理小种菌株01-23相比,Cluster 397.3基因簇各基因的表达规律相同,但表达量存在差异。在接种后第3天,除CHP基因外,01-11菌株其他基因相对表达量均显著高于01-23菌株,其中CPS、P450_1、SDR、ZOR、PKS及P450_2等6个基因的表达差异达到极显著水平,表明Cluster 397.3基因簇的表达存在菌株特异性。
由图4和图5还可知,在接种第3天时,Cluster 397.3基因簇的SDR、DRT_1和ZOR基因表达量均较高,说明在病菌侵染初期主要以氧化还原代谢反应和功能蛋白运输为主;其次为P450和PKS,其中P450主要参与生物氧化,而PKS主要用于合成病菌的特异致病因子;CPS表达量较低,其在病菌侵染过程中主要负责氨基酸代谢过程中NH3的转运。
3 讨 论
聚酮合酶是介导聚酮化合物生物合成的关键酶,广泛存在于细菌和真菌中。聚酮化合物是一类具有广泛生物活性的次级代谢产物,与微生物抗逆、分生孢子产生、成熟附着胞的形成、分生孢子存活、病菌毒力和致病力等密切相关[20-22]。张鑫等[23-24]在玉米大斑病菌基因组内发现了1个与黑色素合成及致病性有关的聚酮合酶基因,且该基因的表达量下降会影响 DHN黑色素的合成。本研究发现Cluster 397.3是一个与玉米大斑病菌HST合成有关的基因簇,该基因簇不仅含有1个聚酮合酶基因,还包含氨基甲酰磷酸合成酶、短链脱氢酶/还原酶、锌结合氧化还原酶、耐药转运蛋白、细胞色素P450等其他8个基因。通过共线性分析发现,交链链格孢、芸薹生链格孢、壳二孢疫病病菌和玉米小斑病菌均存在Cluster 397.3的同源基因簇,表明该基因簇具有较强的保守性。
利用HST接种植物材料可以判断植物病原真菌的致病性和寄主植物的抗病性,而且HST接种比用病原真菌接种更能迅速且大量地鉴定出感病和抗病材料[25-27],因此HST在植物抗病性鉴定和抗病育种应用方面具有极大的优越性。利用HST生物合成基因,还可以进行真菌种属及专化型的鉴定。研制钝化或抑制HST的化学或生物药剂,可以开发新型杀菌剂。某些HST是除草剂、杀虫剂、杀菌剂,甚至某些医药的重要化学成分,因此HST在农药和医药开发方面也有重要作用[28-31]。
本研究对病菌侵染过程中的表达分析结果表明,Cluster 397.3基因簇各基因的表达在01-23(23号生理小种)和01-11(1号生理小种)菌株间存在显著差异,推测该基因簇可能为玉米大斑病菌生理小种分化和致病特异性的关键基因。因此,确定Cluster 397.3为玉米大斑病菌的候选HST生物合成基因簇,该基因簇与ACT-毒素合成基因簇高度同源。
研究表明,交链链格孢ACT-毒素的作用位点主要在寄主的质膜和叶绿体上,可造成细胞膜透性改变进而引起电解质外渗,或破坏叶绿体膜结构而抑制光合作用[15]。本研究结果表明,玉米大斑病菌HST合成基因簇基因在病菌接种第3天时高效表达,表明HST的合成主要发生在病菌侵染初期,以破坏玉米叶片细胞和叶绿素合成为主。Miyamoto等[32]发现,ACT-毒素并非都引起感病反应,它还可以作为一种激发子诱导抗病相关基因表达,进而引起抗病反应,因此ACT-毒素具有极强的寄主选择性。本研究发现,玉米大斑病菌HST合成基因簇的9个基因中,除了CHP基因外,其他基因的表达量在01-11和01-23菌株间表现出了显著或极显著差异,推测HST合成基因簇的表达差异可能与病菌生理小种有关。
猜你喜欢
湖北农业科学(2022年11期)2022-07-18
中国饲料(2022年5期)2022-04-26
考试与评价·高二版(2021年3期)2021-09-10
食品安全导刊(2021年21期)2021-08-30
小天使·二年级语数英综合(2020年6期)2020-12-23
小猕猴智力画刊(2017年12期)2017-12-27
中文信息(2017年2期)2017-04-13
江苏农业科学(2016年11期)2017-03-21
百科知识(2017年2期)2017-03-17
智能计算机与应用(2016年4期)2016-09-26
