
NaCl胁迫对黑杨派无性系苗期生理生化特性的影响
2022-07-14 07:54梁青兰乔艳辉韩友吉刘桂民张升祥李善文董玉峰张兰英张富国
山东林业科技 2022年3期
梁青兰,乔艳辉,韩友吉,任 飞,刘桂民,张升祥,李善文,董玉峰,张兰英,张富国
(1.山东农业大学林学院,山东 泰安 271018;2.山东省林业科学研究院 山东省林木遗传改良重点实验室,山东 济南 250014;3.济南市国有北郊林场,山东 济南 250119;4.山东绘园地理信息有限公司,山东 济南 250101)
黑杨派(Sect.Aigeiros)树种是杨柳科(Salicaceae)杨属(Populus)落叶乔木,由于生长速度快、品质好、抗逆性强等优点,在全世界范围内广泛栽培,是我国重要的工业用材林和生态防护林树种,在盐碱地改良方面也有较好的应用前景。 盐胁迫作为影响全世界作物产量的重要非生物因子[1],给植物造成渗透胁迫,致使细胞内离子代谢紊乱[2],制约其基础代谢活动,对植物正常生理进程造成影响[3]。 研究表明,盐胁迫环境可以通过降低土壤渗透势、破坏植物根系离子平衡[4-6]、影响抗氧化酶机制[7-8]以及渗透调节方式[9-11]来影响植物生理机能,从而对植物生长发育产生抑制作用,严重时会导致植物死亡。 因此,研究盐胁迫下杨树的耐受机制,对于其在盐碱地区合理种植和推广应用具有重要意义[12]。
目前,关于杨树的耐盐性进行了大量研究报道。 程方等[13]、罗永亚[14]、杨传宝等[15-16]对不同杨树无性系的在盐胁迫下的生理特性进行了研究。 本试验利用盆栽试验方法, 对8 个初选的黑杨派无性系进行NaCl 处理,以目前推广应用品种为对照,通过测定盐胁迫下各供试无性系苗期生长指标、生理生化指标,探讨各供试新无性系对逆境的适应能力,采用主成分分析法进行综合评价,筛选出耐盐性强的无性系,为盐碱地区造林提供无性系选择和理论依据。
1 材料与方法
1.1 试验材料
试验材料为10 个黑杨派无性系, 其中8 个是山东省林业科学研究院根据3年生试验林的表型性状初选的优良无性系,以目前推广应用的I-107 杨和2025 杨为对照。
1.2 研究方法
1.2.1 试验方法和设计
采用营养液沙培法。2021年2月下旬取各无性系1年生扦插苗的苗干放于2~4℃冷库中密封储藏备用,于2021年4月中旬截取插穗进行扦插试验。 在规格为170 mm×150 mm的底部有透气孔的花盆底部放置一层无纺布,然后装入等质量消毒后的细沙, 将3 个花盆放在长×宽×高为540 mm×360 mm×150 mm 的同一塑料箱中。 将黑杨派无性系插穗扦插入花盆中,每个无性系重复3 次,扦插时应将插条顶端的芽露出沙子表面。用1/2 Hoaglang 营养液定期向箱内补充定量的营养液,保证植株正常生长。 扦插后的无性系置于山东省林业科学研究院试验苗圃的温室中,进行管理。
扦插苗生长2 个月后,于2021年6月27日进行NaCl 胁迫试验,用1/2 Hoaglang 营养液和NaCl 配制溶液,分别设置5 个盐浓度梯度:0%、0.2%、0.4%、0.6%、0.8%,即CK、T1、T2、T3、T4,将这5 种浓度的溶液(pH值为7.0)分别倒入5 个塑料箱。
1.2.2 测定指标及方法
(1)生长指标的测定
苗高和地径:于2021年6月27日在胁迫开始前测定所有供试无性系的苗高H0和地径B0,7月11日测定胁迫后各无性系的苗高H1和地径B1,试验期间的生长量增量为:
苗高增量= H1-H0
地径增量= B1-B0
其中,无性系苗高(cm)用刻度尺测量,地径(mm)使用游标卡尺测量。
生物量:在完成试验各指标取样结束后,将各处理的无性系苗木拔出洗净,去除表面杂物,将植株的根、茎、叶分开,放入烘箱在105℃杀青,并在75℃下烘至恒重,然后用电子天平称其干重。
(2)生理生化指标的测定
2021年7月11日取样,每株苗木取第4 片、第5 片和第6 片叶(从顶端数),3 次重复。 参照李合生[17]的方法进行生理生化指标的测定。 叶绿素含量采用乙醇提取比色法测定;细胞膜透性采用相对电导率法测定;丙二醛含量采用硫代巴比妥酸法测定;超氧化物歧化酶(SOD)活性测定,SOD 活性采用(NBT)光还原法测定;POD 活性采用愈创木酚法测定;游离脯氨酸(Pro)含量采用茚三酮染色法测定。 各指标测定均重复3 次。
1.3 数据统计分析
运用Excel 2010 整理数据和作图,用SPSS 22.0 软件进行多重比较(Duncan’法)、相关分析、主成分分析等,对黑杨派无性系进行耐盐性综合评价。
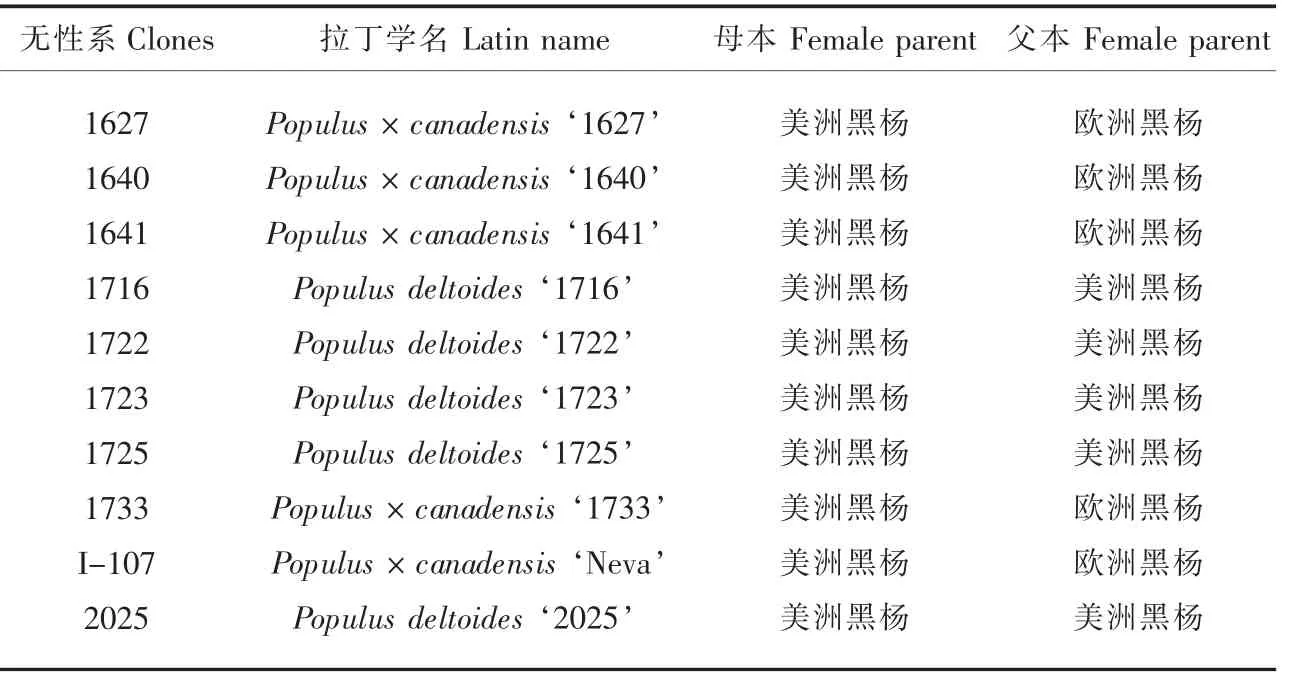
表1 试验材料Table 1 Experimemntal materials
2 结果与分析
2.1 盐胁迫对黑杨派无性系苗高的影响
通过比较NaCl 处理期间各无性系苗高的增量可以看出, 盐胁迫对各供试无性系的苗高生长有明显的抑制作用,见表2。随着盐胁迫程度加深,10 个无性系的苗高增量显著下降, 不同无性系的下降程度不同。在T1 处理时,各无性系与对照2025 相比差异显著, 在该处理下苗高增量最高的是无性系I-107、1733、1725, 增长量分别为8.0 cm、6.9 cm、6.3 cm。 在T2、T3 和T4处理下, 苗高增量排在前3 位的均是无性系1725、I-107 和1733。 说明在不同浓度盐胁迫下, 这3 个无性系具有较大的生长量,表现出较强的耐盐性。
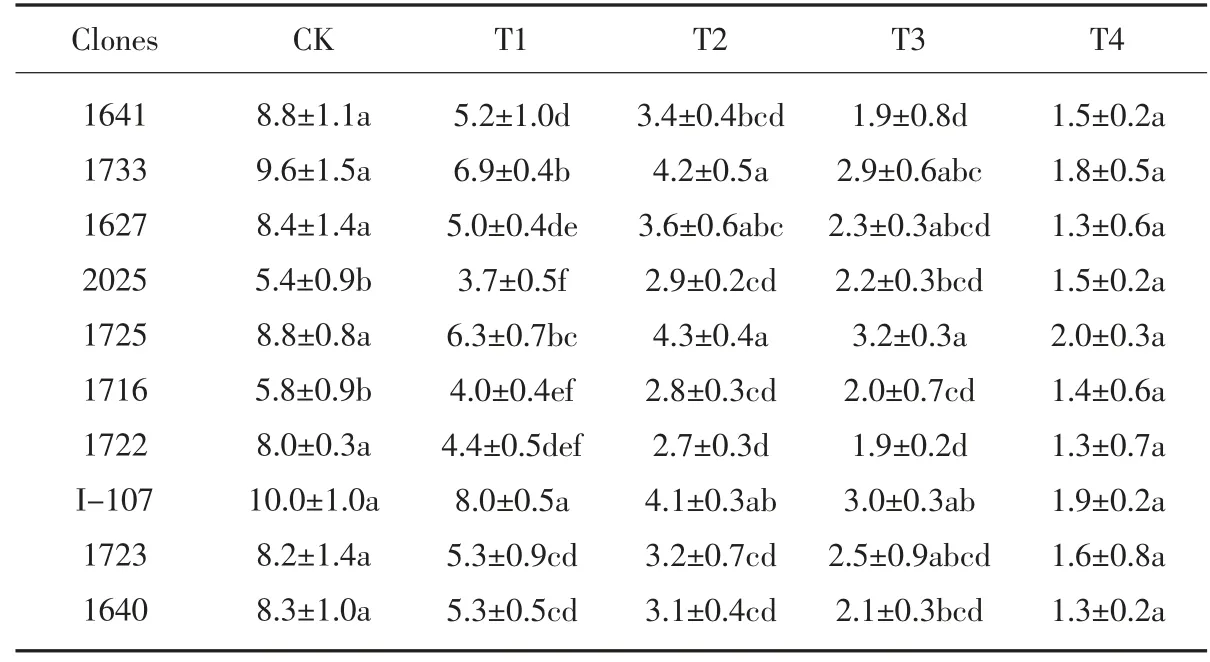
表2 盐胁迫对各无性系苗高增量(mm)的影响Table 2 Effect of salt stress on plant height growth increase of each clones
2.2 盐胁迫对黑杨派无性系地径的影响
通过比较NaCl 处理期间各无性系地径的增量可知,随着盐胁迫程度的加剧,10个无性系的地径增量显著降低, 不同无性系的降低程度不同。 如表3 所示,在T1 处理时, 与CK 相比各供试无性系的地径增量具有显著变化。 在T3 处理时, 无性系1725 和1733 的地径增量最大, 显著高于对照2025; 在T4 处理时, 无性系1725、1627、1733 的地径增量排在前3 位, 表现出较强的耐盐能力。不同浓度的NaCl 处理下,CK 处理与其他各处理浓度对地径增量的影响差异显著, 这说明盐分胁迫明显抑制了植株地径的生长。
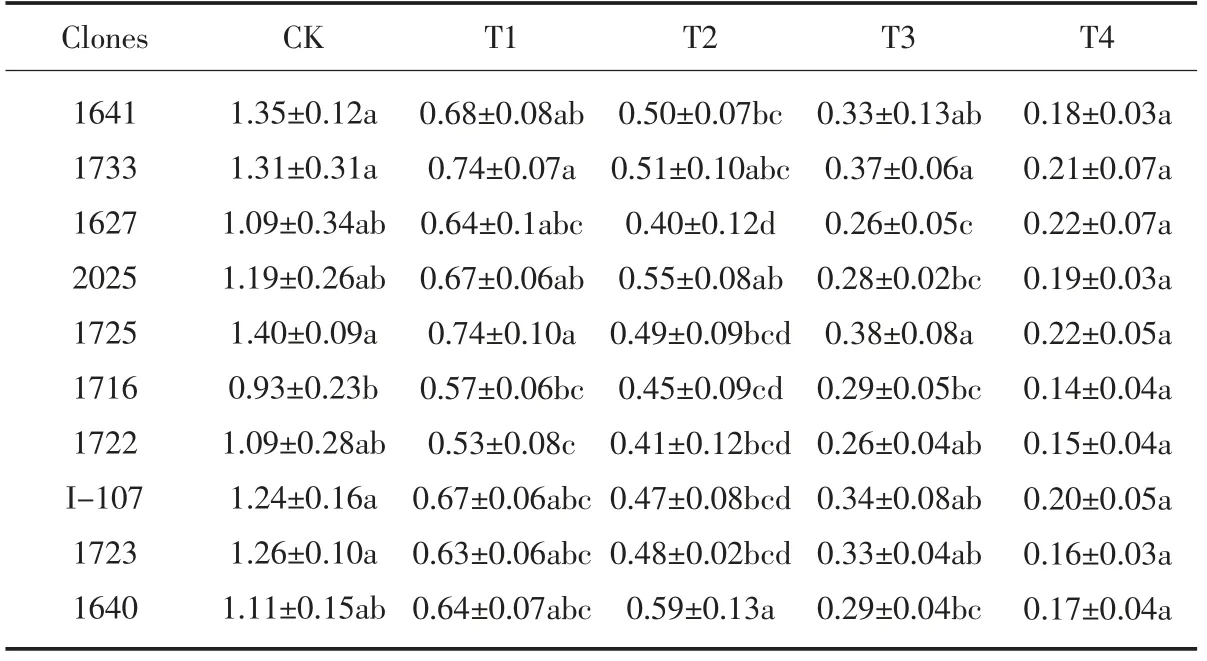
表3 盐胁迫对各无性系地径增量(mm)的影响Table 3 Effect of salt stress on diameter of stem increase of each clones
2.3 盐胁迫对黑杨派无性系生物量的影响
由表4 可以看出,随着NaCl 浓度的提高, 各供试无性系生物量的变化趋势基本一致, 均不断降低。 在不同浓度的盐处理下,无性系的生物量有所差异,其中,CK 处理下的生物量最高,其次是T1 处理,再次是T2 处理,T3、T4 处理下的生物量比较小。 在T4 处理时, 生物量最大的无性系是1733 和1725, 分别为1.49g、1.36g,与对照2025 的差异达显著水平;生物量最小的是2025,为0.85g,这说明无性系1733 和1725受到NaCl 胁迫的影响较小,2025 受到的影响较大。
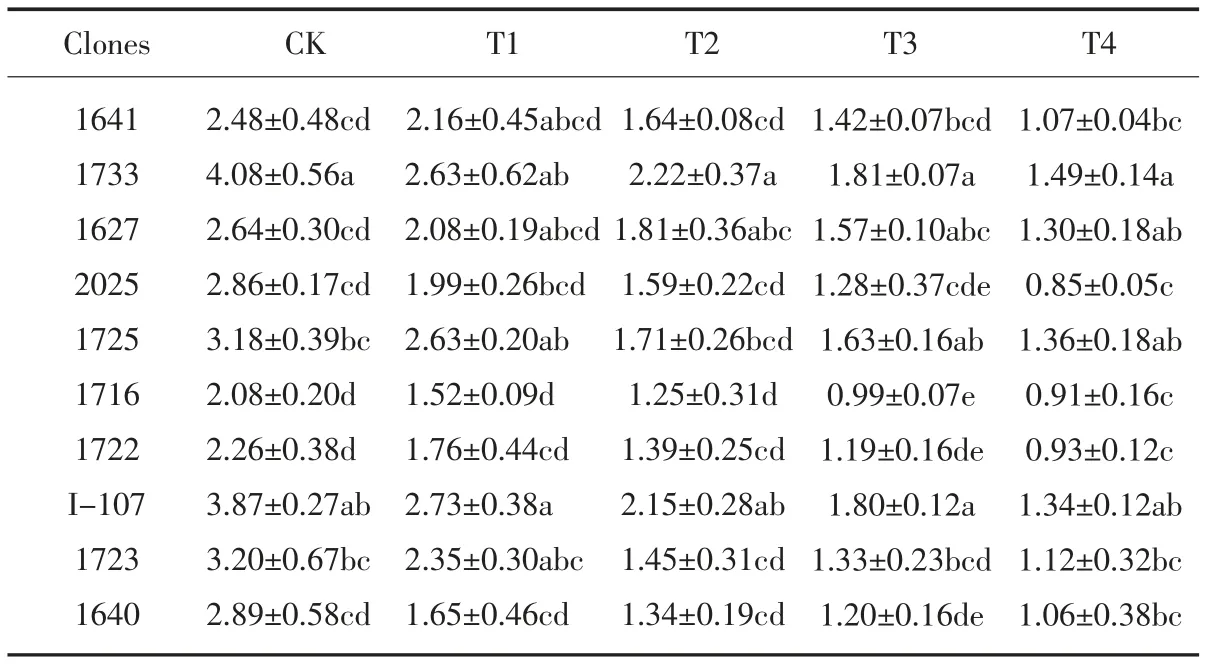
表4 盐胁迫对各无性系生物量的影响Table 4 Effect of salt stress on biomass of each clones
2.4 盐胁迫对黑杨派无性系叶绿素含量的影响
随着盐处理浓度的增加,10 个黑杨派无性系的叶绿素含量变化如表5 所示, 各供试无性系叶片的叶绿素含量随盐胁迫程度的加深呈现出下降趋势。在T1 处理下,与CK 相比无性系1733 的叶绿素含量的下降幅度最小,降幅为6.3%,1723 的下降幅度最大,降幅为28.4%22.30%;在T3 和T4 处理下,无性系2025、1725的叶绿素含量较高,均在1.51 mg·g-1以上, 1722 的叶绿素含量最小,小于1 mg·g-1,表明盐分胁迫对1722的光合作用抑制明显,对2025 和1725 的抑制作用小。试验结果说明,NaCl 处理导致植物叶绿素含量显著降低,盐分胁迫对叶绿素有明显的破坏作用。
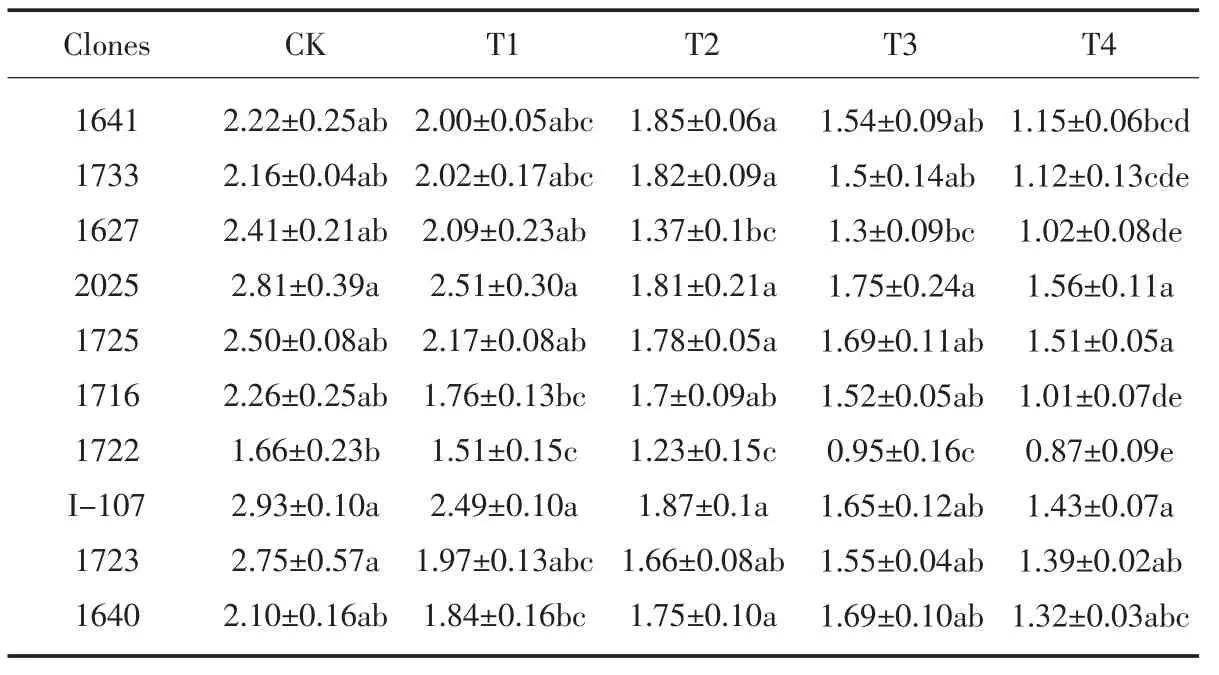
表5 盐胁迫对各无性系叶绿素含量(mg·g-1)的影响Table 5 Effect of salt stress on chlorophyll content of each clones
2.5 盐胁迫对黑杨派无性系细胞膜透性的影响
在盐处理环境下,10 个黑杨派无性系的细胞膜透性随NaCl 浓度的升高而增大(见表6), 说明盐分胁迫对细胞膜的伤害程度也随之增大。 在T1 和T2 处理时,无性系1733 和1725 细胞膜透性较低,与对照无性系2025 差异显著, 表现出较好的耐盐能力。 在T3 处理时,细胞膜通透性增加幅度最大的是1716 和1641 ,分别达到173.1%和156.2% , 增加幅度最小的是1725 和I-107,分别为88.7%和89.6%。 说明无性系1641 和1716 的细胞膜容易受到盐胁迫伤害,无性系I-107 和1725 受到盐胁迫的影响较小。
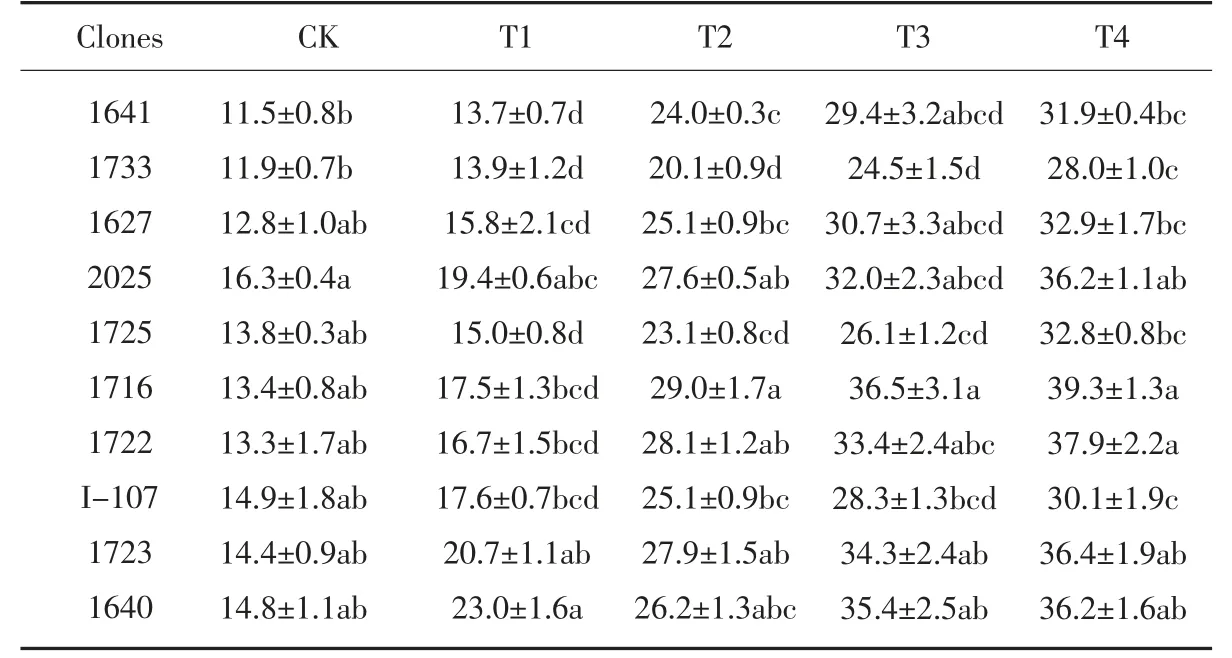
表6 盐胁迫对各无性系细胞膜透性的影响(单位:%)Table 6 Effect of salt stress on diameter of membrane permeability of each clones
2.6 盐胁迫对黑杨派无性系丙二醛含量的影响
从表7 可以看出,盐胁迫处理下的各供试无性系叶片中的丙二醛含量均较CK明显升高, 在T4 处理下增至最大。 在T1处理时, 与CK 相比,10 个无性系的叶片丙二醛含量升高幅度在38.1% ~77.7 %之间;在T2 处理时,升高幅度较小的无性系有1733、1725、2025, 增幅在87.8%以下,升高幅度较大的有1722、1716, 增幅在118.8%以上; 在T3 处理时, 无性系1725和1733 的MDA 含量和升高幅度均最小,均低于对照I-107、2025 及其他参试无性系,说明其受到盐胁迫的影响较小;在T4高盐浓度处理下,所有无性系的丙二醛含量都达到较高水平,盐胁迫对植物细胞膜产生的破坏加重,无性系的受到的盐害作用增大。
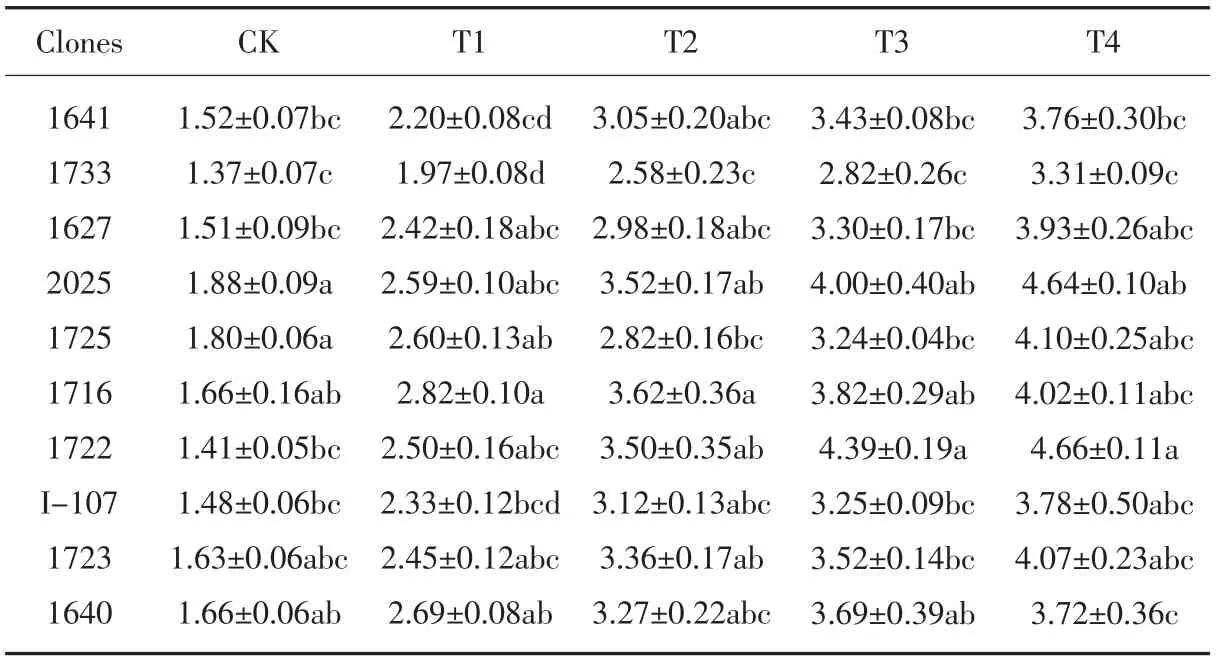
表7 盐胁迫对各无性系MDA 含量的影响Table 6 Effect of salt stress on MDA content of each clones
2.7 盐胁迫对黑杨派无性系SOD 活性的影响
各黑杨派无性系的SOD 活性变化规律如表8 所示, 随着NaCl 浓度增加SOD活性表现为先升后降。 在T1 处理浓度时,各供试无性系的SOD 活性处于最高水平,其中,升高幅度最大的无性系是1723(53.0%)、I-107(44.6%),升高幅度最小的是1641(21.2%)、1716(22.3%);但在T2 至T4 处理下,无性系的SOD 活性开始下降,这表明,在低浓度盐胁迫下10 个无性系能够通过增强SOD 的活性抵抗盐分伤害,当胁迫加重,抗氧化酶活性降低,植物会受到盐害作用。T2-T4 浓度处理下,无性系1725 的SOD 活性始终较高,居于前两位,说明其在高浓度的盐分胁迫下受影响较小。
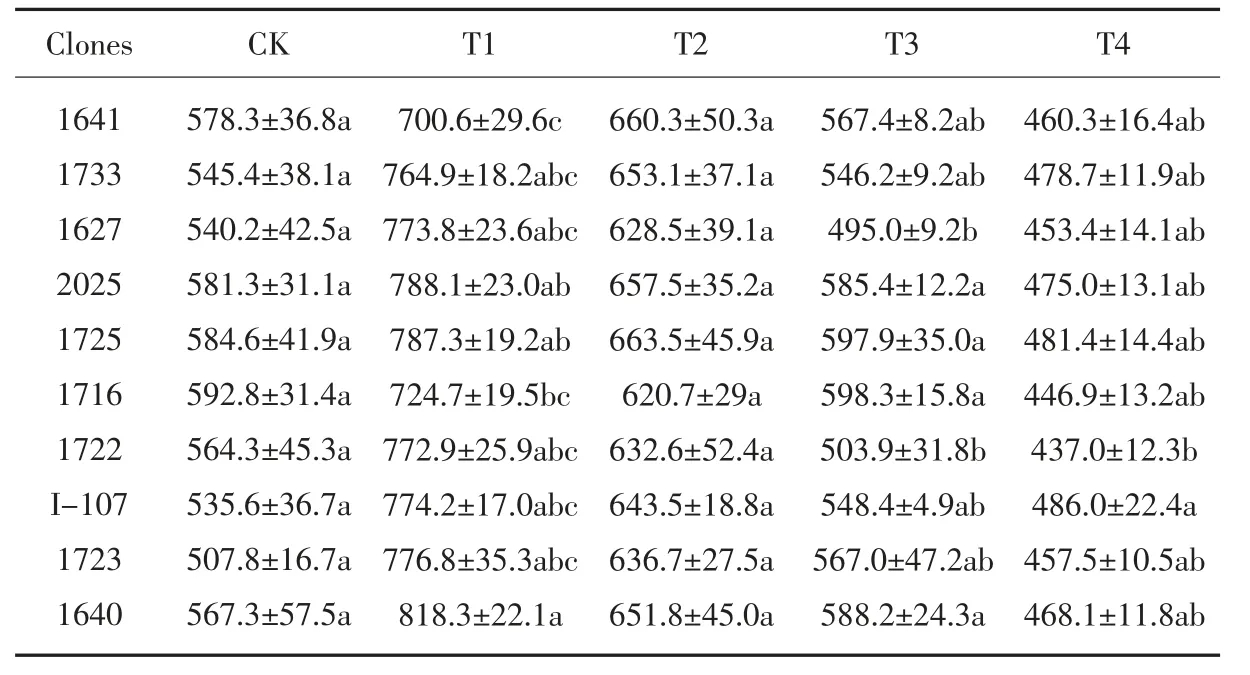
表8 盐胁迫对各无性系SOD 活性的影响Table 8 Effect of salt stress on SOD activity of each clones
2.8 盐胁迫对黑杨派无性系POD 活性的影响
各供试无性系叶片POD 活性的变化规律与SOD 大体一致(见表9),所有无性系的POD 活性在T1 处理下与CK 相比显著升高,POD 活性达到最高值, 其中无性系1733、I-107 的POD 活性和升高幅度最大, 增幅为202.3%和201.3%。 随后随着NaCl 处理浓度增加, 各无性系POD 活性降低, 在T4 处理时降至最低, 此时POD活性最高的无性系为1733 和1725, 与对照2025 达到显著性差异水平。 试验结果表明, 不同浓度的NaCl 处理对于黑杨派无性系POD 活性产生的影响明显不同。

表9 盐胁迫对各无性系POD 活性的影响Table 8 Effect of salt stress on POD activity of each clones
2.9 盐胁迫对黑杨派无性系游离脯氨酸含量的影响
如表10 所示, 各供试无性系叶片的游离脯氨酸含量各有差异,除无性系1640的叶片脯氨酸含量呈先下降后上升外,其余无性系的脯氨酸含量随着NaCl 浓度的增加表现为不断上升趋势, 并在T4 处理时达到最高值。 在T2 处理时, 无性系1641、1733 和1725 的脯氨酸含量增加幅度最大,均在97.1%以上,明显高于对照2025、I-107 及其他无性系;在T4 处理时,相比于其他无性系, 1733、1725 的游离脯氨酸含量处于较高水平,在60.8mg·g-1以上,说明在盐胁迫条件下,其参与渗透调节的能力较强。
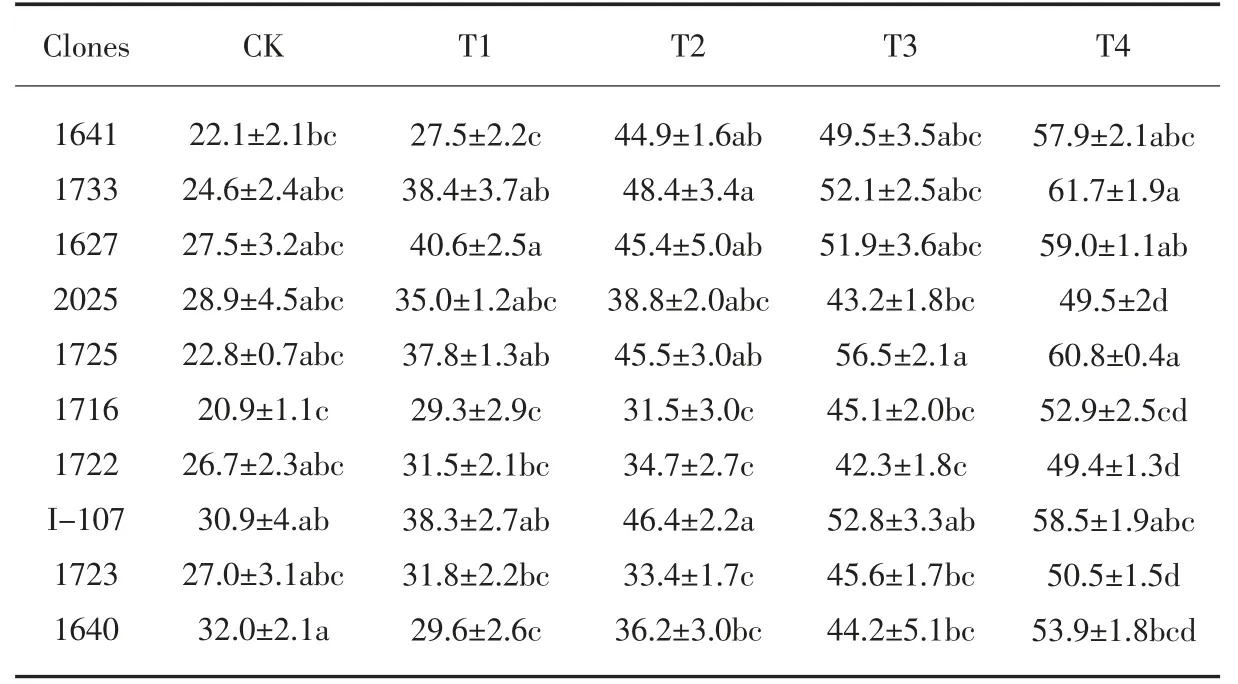
表10 盐胁迫对各无性系游离脯氨酸含量的影响Tab.10 Effect of salt stress on Proline content of each clones
2.10 主成分分析
对T2、T3 和T4 处理下10 个无性系的各项指标进主成分分析,见表11。在T2处理下, 第1 主成分贡献率为65.6%,第2 主成分贡献率为21.7%, 累计贡献率达87.3%, 表明这2 个主成分可以把原来9项指标87.3%的信息表达出来。 第1 主成分中苗高增量、生物量、细胞膜透性、POD活性、丙二醛含量、脯氨酸含量的特征向量绝对值较大,这表明如果无性系的第1 主成分值大,其苗高增量、生物量、POD 活性、脯氨酸含量值也大,而细胞膜透性、丙二醛含量值小,说明该无性系耐盐性强。第2 主成分中地径增量、叶绿素含量、SOD 活性的特征向量绝对值较大。

表11 NaCl 胁迫下各指标的主成分特征向量、特征值及累计贡献率Tab.11 Principal component characteristic vector, characteristic value and cumulative contribution rate of each index under NaCl stress
在T3 处理下, 第1 主成分贡献率为64.6%,第2 主成分贡献率为22.0%, 累计贡献率达86.7%, 可以反映原来86.7%的信息。 第1 主成分中苗高增量、地径增量、生物量、细胞膜透性、丙二醛含量、POD 活性、脯氨酸含量的特征向量绝对值较大,其中除细胞膜透性和丙二醛含量的向量为负值外, 其余均为正值;这表明如果无性系的第1 主成分值大,其苗高增量、地径增量、生物量、POD 活性、脯氨酸含量值也大,而细胞膜透性、丙二醛含量值小,说明该无性系耐盐性强。 第2 主成分中叶绿素含量、SOD 活性的特征向量较大。
在T4 处理下,第1 主成分贡献率为68.7%,第2 主成分贡献率为16.8%,累计贡献率达85.5%,可以反映原来9 项指标85.5%的信息。 在T4 处理下,第1 主成分中苗高增量、地径增量、生物量、细胞膜透性、SOD活性、POD 活性、脯氨酸含量的特征向量绝对值较大,其中除细胞膜透性的向量为负值外,其余均为正值;这表明如果无性系的第一主成分值大,其苗高增量、地径增量、生物量、SOD 活性、POD 活性、脯氨酸含量值也大,而细胞膜透性值小,说明该无性系耐盐性强。 第2 主成分中叶绿素含量、SOD 活性、丙二醛含量的特征向量较大。
计算T2、T3 和T4 处理下10 个无性系的前2 个主成分的综合值(表12),并计算每个无性系这3 个处理的综合主成分值的均值,根据均值大小对各个无性系的耐盐性进行排序,各供试无性系的耐盐能力强弱依次为: 1733、1725、I-107、1641、1627、2025、1640、1723、1716、1722。
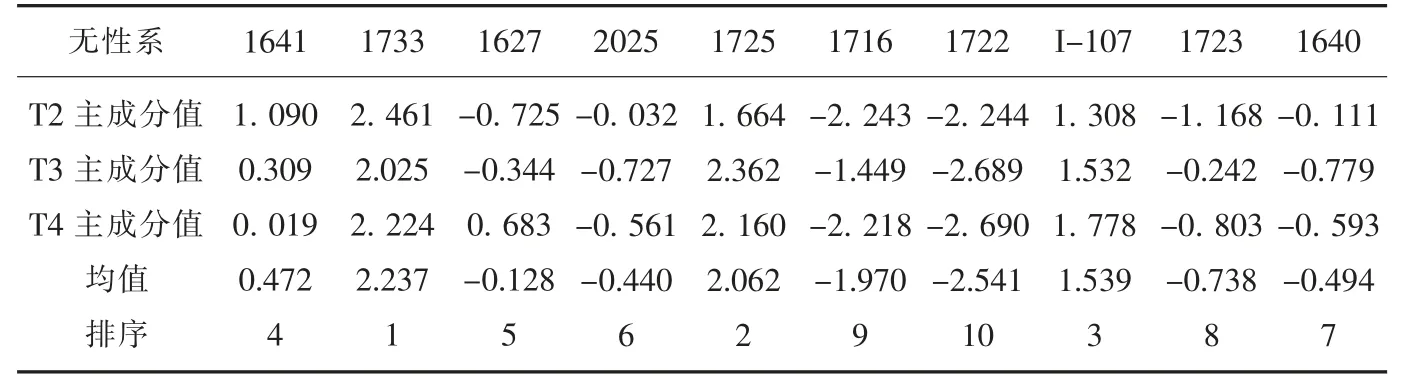
表12 NaCl 胁迫下10 个无性系的综合主成分值Tab.12 Intergrated principal component values of 10 clones under NaCl stress
3 讨论与结论
植物处于盐渍环境下,其形态发育、光合作用、渗透调节等都会受到影响,其中最直观的就是抑制植株的生长,植物的生长量变化是判断其受到盐害程度的重要测量标准[18]。在本试验中,随着盐胁迫程度的加深,10 个无性系的苗高增量、地径增量和生物量均呈下降趋势。说明过量的盐进入植物体内对其造成损伤,影响植株生长发育。 这与杨传宝等[15-16]对白杨派(Leuce)无性系、罗永亚[14]对胡杨(P.euphratica)的研究结果一致。
叶绿素在植物的光合作用过程发挥着重要的作用,在正常情况下,植物通过光合作用制造有机物,维持植株正常生长[19]。而盐胁迫会严重影响植物叶片中叶绿素的含量,降低植物的光合效率,抑制植物正常生长。孙伟博等[20]以杨树为试验材料,对其进行NaCl 胁迫,研究结果发现,杨树植株叶绿素含量随盐浓度的升高呈下降趋势。 这与本试验结果一致,随着盐胁迫的加深,各供试无性系的叶片绿素含量与对照比较均逐渐降低。
植物在逆境胁迫下细胞膜是重要的生理结构,可以控制细胞内外物质进出,维持细胞代谢稳定,维持正常生理活动[21]。盐分胁迫会破坏细胞膜结构,引起丙二醛含量增加,对细胞产生毒害作用,导致细胞膜的结构与功能受损[22-23]。 在本试验中,10 个无性系的细胞膜透性和MDA 含量呈逐渐上升的趋势,这与对金银花(Lonicerae japonicae )[24]、猕猴桃(Actinidia deliciosa)[25]等植物的研究结论相似。 说明较高的盐浓度会导致植物细胞膜脂质过氧化作用加强,细胞膜受害加深。
超氧化物歧化酶(SOD)和过氧化物酶(POD)是植物抗氧化防御体系中重要的保护酶,二者在逆境环境下能够将植物体内过多的活性氧清除,起到减轻细胞膜氧化损伤的作用。 在本试验中,在T1 处理下,各无性系SOD 活性与对照比较明显增加,在T2 处理时SOD 活性出现降低,这可能是由于植物在低盐浓度下通过提高酶活性来抵御盐胁迫,减轻活性氧带来的氧化伤害,当胁迫程度加深,达到植物自身所承受的最大限度,细胞膜系统受到伤害,保护酶活性就会降低。 这与李翀等[26]对‘南林895’杂交杨(P.× euramericana‘Nanlin895’)研究结果一致,
游离脯氨酸是一种重要的渗透调节物质,能够调节组织细胞的渗透压,维护细胞结构和功能稳定。 本试验中, 随着处理盐胁迫程度加重,10 个黑杨派无性系的脯氨酸含量不断增加, 这说明在遇到高浓度盐胁迫时,为了维持自身生命活动的正常进行,植物会通过增加脯氨酸的含量来缓解盐分伤害,抵抗盐害导致的渗透胁迫。 这与史军辉等[27]对胡杨的研究结果相同。
本研究对10 个黑杨派无性系进行不同浓度的NaCl 处理, 通过分析各无性系苗期的各项指标变化,探究盐胁迫对各供试无性系生长和生理生化特性的影响。 根据主成分分析,得到各无性系的耐盐性由强到弱依次为1733、1725、I-107、1641、1627、2025、1640、1723、1716、1722。 初步筛选出1733 和1725 两个耐盐能力较强的无性系,可以作为耐盐试验材料进一步在盐碱地开展田间试验研究。
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
农业工程学报(2022年10期)2022-08-22
作物学报(2022年8期)2022-05-29
现代园艺(2022年6期)2022-04-08
安徽化工(2021年3期)2021-05-29
现代农业科技(2020年10期)2020-06-04
安徽农学通报(2020年7期)2020-05-26
科学与财富(2019年15期)2019-10-21
分析化学(2018年11期)2018-01-16
课程教育研究·新教师教学(2016年14期)2017-04-12
