
堆型艾美耳球虫重组亚单位疫苗的制备及其免疫效果评价
2022-07-19 12:31毕天奇肇英池马丹程淑琴王晓岑李新李建华张西臣宫鹏涛张楠张媛媛
中国生物制品学杂志 2022年7期
毕天奇,肇英池,马丹,程淑琴,王晓岑,李新,李建华,张西臣,宫鹏涛,张楠,张媛媛
1.吉林大学动物医学学院,吉林 长春 130062;2.吉林大学人兽共患病研究所,吉林 长春 130062
鸡球虫病是一种寄生性原虫病[1],主要侵害鸡肠道上皮细胞,分布广泛,是严重危害养鸡业的寄生虫病之一[2]。目前,国际公认危害鸡的艾美耳球虫有7 种,即柔嫩艾美耳球虫(Eimeria tenella)、堆型艾美耳球虫(Eimeria acervulina)、毒害艾美耳球虫(Eimeri anecateix)、巨型艾美耳球虫(Eimeria maxima)、缓艾美耳球虫(Eimeria mitis)、早熟艾美耳球虫(Eimeria praecox)和布什艾美耳球虫(Eimeria brunetti)[3]。堆型艾美耳球虫主要影响肉用仔鸡及后备母鸡,导致母鸡增重率及产蛋量下降[4]。以往对鸡球虫的防控主要依赖抗球虫药物和活疫苗[5],但化学药物的大量使用可导致药物残留,影响动物源性食品安全,现已禁用部分抗球虫药物;而活疫苗毒力存在返强和散毒的风险[6]。因此,为促进养鸡业更加健康地发展,研制稳定安全的抗球虫疫苗十分必要。
近年,有研究已筛选出较多球虫保护性基因,如SO7、cSZ1、NA4、TA4、AMA1等[7],其中cSZ1基因具有高度的保守性,且cSZ1 蛋白免疫原性良好,是堆型艾美耳球虫疫苗研究的候选蛋白[8],但目前尚无毕赤酵母系统表达cSZ1基因的相关报道。本研究通过毕赤酵母表达堆型艾美耳球虫cSZ1基因,制备了重组亚单位疫苗,并评价其免疫效果,以期为研制安全高效的堆型艾美耳球虫重组亚单位疫苗提供候选抗原。
1 材料与方法
1.1载体、菌株及虫株 克隆载体pMD18-T 购自宝日医生物技术(北京)有限公司;感受态毕赤酵母菌株GS115、酵母表达载体pPICZαA 及E.acervulina孢子化卵囊均由吉林大学寄生虫实验室保存;感受态E.coliDH5α 购自沈阳天根生物技术有限公司。
1.2主要试剂 反转录试剂盒、限制性内切酶SacⅠ、EcoRⅠ、SalⅠ、鼠源His 标签单克隆抗体、DNA T4 连接酶、HRP 标记的山羊抗鼠及山羊抗鸡IgG 均购自日本TaKaRa 公司;胶回收试剂盒、质粒小提试剂盒、DL2000 DNA marker、DL5000 DNA marker 及rTaq酶均购自沈阳天根生物技术有限公司;鸡淋巴细胞分离试剂盒购自北京索莱宝科技有限公司;大刀豆素(ConA)、CCK-8、弗氏完全佐剂及弗氏不完全佐剂均购自美国Sigma 公司;其他试剂均由吉林大学动物医学学院寄生虫实验室提供。
1.3实验动物 健康海兰褐雏鸡,雄性,150 只,7 日龄,购自长春市农业科学院,均于无球虫污染的环境中饲养,自由饮食。本实验对雏鸡的所有处理均以科研为目的进行养殖和使用,且按照动物伦理相关规定进行(文件号为:SY202007001)。
1.4目的基因的扩增 根据GenBank 中登录的cSZ1基因序列(AY513879.1),结合克隆载体pMD18-T 和酵母表达载体pPICZαA 多克隆位点序列,应用Primer5.0 软件设计引物,cSZ1-F:5'-GAATTCATGGGTGAAGAGGCTGATACTCA-3',cSZ1-R:5'-GTCGACTTAGAAGCCGCCCTGGTACA-3',扩增产物大小为513 bp。引物由吉林省库美生物科技有限公司合成。以堆型艾美耳球虫cDNA 为模板,进行PCR 扩增。PCR 扩增条件为:95 ℃5 min;95 ℃30 s,57 ℃30 s,72 ℃60 s,共30 个循环;72 ℃10 min,4 ℃保存。扩增产物经1%琼脂糖凝胶电泳分析。
1.5重组表达质粒pPICZαA-cSZ1的构建 胶回收目的基因片段,连接至克隆载体pMD18-T,转化至感受态E.coilDH5α 中,筛选阳性菌落,用DNA质粒小提试剂盒提取重组质粒。将酵母表达载体pPICZαA 和重组质粒pMD18-T-cSZ1经EcoRⅠ和SalⅠ双酶切,回收载体片段及目的基因片段,以T4 DNA 连接酶于37 ℃连接过夜;转化至感受态E.coilDH5α 中,挑取单克隆,进行EcoRⅠ和SalⅠ双酶切鉴定。鉴定正确的质粒送吉林省库美生物科技有限公司测序,测序正确的重组表达质粒命名为pPICZαA-cSZ1。
1.6重组菌GS115-pPICZαA-cSZ1的构建 用SacⅠ酶线性化重组表达质粒pPICZαA-cSZ1,无菌取线性化质粒20 μL,加入感受态毕赤酵母菌株GS115 80 μL,混匀,冰浴15 min;电穿孔转化,取90 μL,涂布于YPD 平板,于30 ℃恒温培养箱培养2 ~3 d;出现白色菌落后,用酵母通用引物AOX1(上游:5'-GACTGGTTCCAATTGACAAGG-3',下游:5'-GGCAAATGGCATTCTGACATCCT-3')进行菌落PCR 检测。阳性菌落送吉林省库美生物科技有限公司测序,测序正确的重组菌命名为GS115-pPICZαA-cSZ1。
1.7目的基因的诱导表达 将重组菌GS115-pPICZαAcSZ1用6 mL YPD 液体培养基于30 ℃,200 r/min 培养16 h;加至50 mL BMGY 培养基中扩大培养,继续培养约12 h;菌液A600为5 时,2 000×g离心5 min;将菌体沉淀移至100 mL BMMY 液体培养基中,继续培养,每隔24 h 加入1 mL 甲醇,并取1 mL 菌液,2 000×g离心3 min,取上清,进行10% SDS-PAGE 分析,直至144 h。取甲醇诱导120 h 的上清,经10%SDS-PAGE分离蛋白后,转移至PVDF 膜,用5%脱脂奶粉37 ℃封闭2 h;加入His 标签单克隆抗体(1 ∶1 000 稀释),4 ℃孵育过夜;TBST 洗涤3 次,加入HRP 标记的山羊抗鸡IgG(1 ∶5 000 稀释),37 ℃孵育1 h;H2SO4终止反应,ECL 显色。
1.8重组蛋白的大量制备及纯化 将重组菌GS115-pPICZαA-cSZ1于BMMY 液体培养基中培养120 h;收取上清液,2 000×g离心5 min,取上清,加入硫酸铵(472 g/L),4 ℃放置14 h;5 000×g离心30 min;取沉淀,用1 × PBS 重悬,饱和硫酸铵溶液透析;透析蛋白经镍柱纯化,获得纯化蛋白cSZ1,BCA 法测定蛋白浓度,-20 ℃保存。
1.9动物分组及免疫 将150 只雏鸡随机分为5 组,每组30 只。其中3 个免疫组分别按20、40、60 μg / 只(低、中、高剂量)经肌肉注射重组蛋白;7 d后进行加强免疫,免疫剂量及途径同首次免疫。末次免疫7 d 后,经口感染E.acervulina孢子化卵囊,剂量均为1 × 104个/ 只。同时设阳性对照组(未免疫,攻虫剂量同免疫组)和阴性对照组(未免疫未攻虫)。
1.10免疫效果评价
1.10.1临床观察 攻虫后3 d 内,观察并记录各组雏鸡的饮食、精神状态及粪便性状等。
1.10.2抗球虫效果 攻虫后7 d,统计各组雏鸡的存活情况,按下式计算存活率;称量雏鸡体重,按下式计算相对增重率。每组取3 只雏鸡,解剖,观察十二指肠组织,根据Johnson 计分方法统计各组雏鸡的十二指肠病变计分[9],计分总和即为病变值;麦氏计数法统计雏鸡十二指肠组织中的卵囊数[3],2 个计数室中卵囊数的平均值(单位为106个)乘以200,即为克粪便卵囊数(Oocysts per gramme,OPG),并换算为卵囊值,换算关系为:粪便排卵囊数(单位为106个)<0.1、0.1 ~1.0、2.0 ~5.0、6.0 ~10.0、>11.0 时对应的卵囊值分别为0、1、10、20、40,按下式计算抗球虫指数(anticoccidial index,ACI)。


1.10.3血清抗体水平检测 采用间接ELISA 法。于初次免疫后7 d 及加强免疫后7 d,取低、中、高剂量免疫组和阴性对照组雏鸡,每组3 只,经心脏采血,分离血清。以制备的cSZ1 蛋白作为抗原包被96 孔板,1 μg /孔,4 ℃包被过夜;PBST 洗涤3 次,用5%脱脂奶粉37 ℃封闭2 h;PBST 洗涤3 次,加入待测血清(1 ∶2 000 稀释),100 μL/孔,37 ℃孵育2 h;PBST 洗涤3 次,加入HRP 标记的兔抗鸡IgG(1 ∶5 000 稀释),100 μL / 孔,37 ℃孵育1 h;PBST 洗涤3 次,加入显色液,50 μL / 孔,37 ℃避光孵育15 min;加入终止液,50 μL / 孔,用酶标仪检测A490。
1.10.4T 淋巴细胞增殖情况的检测 攻虫后7 d,取低、中、高剂量免疫组和阴性对照组雏鸡,每组3只,无菌取脾脏,采用鸡淋巴细胞分离试剂盒分离T淋巴细胞,通过CCK-8 法检测T 淋巴细胞增殖情况(A450)[5]。每组样品设3 个重复。
1.11统计学分析 应用SPSS 26.0 软件进行统计学分析,试验数据以均值± 标准差(±s)表示,组间比较采用单因素方差分析,以P<0.05 为差异有统计学意义。
2 结 果
2.1cSZ1基因PCR 产物的鉴定cSZ1基因PCR产物经1%琼脂糖凝胶分析,可见513 bp 目的基因条带,大小与预期一致,见图1。测序结果表明,扩增序列与GenBank 中登录的序列完全一致。

图1 cSZ1 基因扩增产物电泳图Fig.1 Electrophoretic profile of PCR product of cSZ1 gene
2.2重组表达质粒pPICZaA-cSZ1的鉴定 重组质粒pPICZaA-cSZ1的双酶切产物(EcoRⅠ/ SalⅠ)经1%琼脂糖凝胶电泳分析,可见3 593 和513 bp 的载体片段及目的基因片段,见图2。测序结果显示,碱基序列无突变和移码。表明重组表达质粒构建正确。

图2 重组表达质粒pPICZaA-cSZ1 双酶切(EcoRⅠ/SalⅠ)鉴定Fig.2 Restriction map of recombinant plasmid pPICZaAcSZ1(EcoRⅠ/SalⅠ)
2.3重组菌GS115-pPICZaA-cSZ1的鉴定 重组菌GS115-pPICZaA-cSZ1经菌落PCR 鉴定,可见2 200(GS115)和1 093 bp(目的基因513 bp+载体580 bp)的基因条带,见图3。表明重组菌构建正确。

图3 重组菌GS115-pPICZaA-cSZ1 的菌落PCR 鉴定Fig.3 Identification of GS115-pPICZaA-cSZ1 by colony PCR
2.4重组蛋白的鉴定 甲醇诱导的阳性重组菌株GS115-pPICZaA-cSZ1经10% SDS-PAGE 分析,均可见相对分子质量约19 000 的目的蛋白条带,大小与预期相符,见图4。诱导120 和144 h 的蛋白浓度较高,为节约诱导时间,选择120 h 为诱导时间。甲醇诱导120 h 的蛋白可与鼠抗His 标签单克隆抗体发生特异性结合,于相对分子质量约19 000 处可见特异性结合条带,大小与理论值相符,且条带清晰,无杂带,见图5,表明在毕赤酵母中成功表达重组蛋白cSZ1。
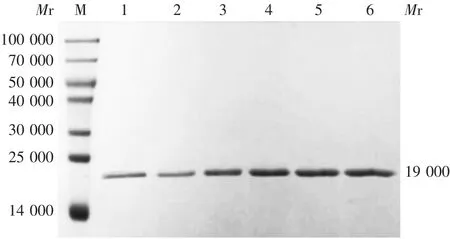
图4 重组蛋白的SDS-PAGE 分析Fig.4 SDS-PAGE profile of recombinant protein

图5 重组蛋白的Western blot 检测Fig.5 Western blotting of recombinant protein
2.5免疫效果
2.5.1临床表现 攻虫后4 d,阳性对照组雏鸡开始出现血便、稀便、羽毛逆立、精神萎靡等症状,其他各组雏鸡偶有稀便,但食欲正常,均未出现死亡。
2.5.2抗球虫效果 与阳性对照组比较,低、中、高剂量免疫组雏鸡的相对增重率明显升高(F分别为7.111、5.740、5.880,P均<0.05),卵囊值明显降低(F分别为63.593、54.795、61.780,P均<0.01),十二指肠病变值显著降低(F分别为10.286、12.250、5.444,P均<0.05)。低剂量免疫组动物的十二指肠病变值高于中、高剂量组,但差异无统计学意义(F分别为0.400 和0.133,P均>0.05)。经计算,低、中、高剂量免疫组ACI 分别为171.25、176.42、174.33,保护效果较好,其中中剂量免疫组保护效果最好。见表1。

表1 重组亚单位疫苗的抗球虫效果Tab.1 ACIs of recombinant subunit vaccine
2.5.3血清抗体水平 随着免疫次数的增加,与阴性对照组比较,低、中、高剂量免疫组雏鸡于初次免疫后7 d 及加强免疫后7 d,血清抗体水平明显升高(F= 1.522 ~13.00,P<0.01),见图6。

图6 各组雏鸡血清中的抗体水平Fig.6 Serum antibody levels of chicks in various groups
2.5.4T 淋巴细胞增殖情况 阴性对照组及低、中、高剂量免疫组雏鸡T 淋巴细胞A450分别为0.291、0.487、0.522、0.507。与阴性对照组比较,低、中、高剂量免疫组雏鸡T 淋巴细胞增殖率显著升高(F分别为4.342、2.594、8.025,P均<0.01)。
3 讨 论
堆型艾美耳球虫是流行性最广的鸡球虫之一[8],虽然感染后不会导致大范围的死亡,但严重影响感染鸡的发育,导致产蛋率及对其他疾病的抵抗力下降[9]。郭衍冰等[4]将构建的重组卡介苗pMV361-cSZ1经滴鼻免疫鸡,发现卵囊产量下降62.7%,ACI高达182.93,呈较好的保护效果。本研究将cSZ1基因连接至酵母载体,获得重组蛋白,经肌肉注射免疫鸡,结果表明,重组蛋白对免疫鸡可产生较好的保护效果,但疫苗的免疫保护效果与注射剂量不呈剂量依赖性,该结果与文献[10]的结论相符。
毕赤酵母表达系统具有高表达,安全无毒的特点,目前已有多种基因在其中得到高效表达[11-13]。毕赤酵母表达系统还具有其他优点:可稳定表达外源蛋白[14];培养基所需成分单一,操作方便[15];分泌上清表达,可对所表达的蛋白进行空间折叠和一定的糖基化修饰,更接近天然蛋白质[16]。ZHANG等[17]将EtMIC2基因分别用毕赤酵母和E.coil进行表达,用获得的重组蛋白免疫雏鸡,结果表明,免疫毕赤酵母表达的蛋白后,增重率、卵囊排出量、盲肠病变上明显优于E.coil表达蛋白组,特异性抗体水平和脾细胞增殖也高于E.coil表达蛋白组,表明毕赤酵母表达的重组蛋白EtMIC2 能够诱导更高的体液免疫,提供更好的免疫保护力。本研究结果表明,与空白对照组比较,用构建的重组亚单位疫苗免疫雏鸡后,体液和细胞免疫水平均显著上升(P<0.05),20、40 和60 μg 剂量免疫组的ACI 值分别为171.25、176.42、174.33,显示了较好的免疫保护效果。
本研究成功构建了真核表达质粒pPICZαA-cSZ1,于毕赤酵母中表达制备的重组亚单位疫苗经不同剂量免疫雏鸡,攻虫试验均显示良好的保护效果。本研究为堆型艾美耳球虫疫苗的大规模生产奠定了基础。
猜你喜欢
中学生物学(2022年8期)2022-10-13
——一道江苏高考题的奥秘解读和拓展
中学生物学(2022年7期)2022-09-07
伴侣(2022年3期)2022-04-08
三农资讯半月报(2020年11期)2020-06-21
江苏农业学报(2019年1期)2019-09-10
家禽科学(2019年3期)2019-07-08
中国动物保健(2018年4期)2018-12-27
中国动物保健(2017年10期)2018-01-29
名人传记·财富人物(2017年9期)2017-11-02
名人传记·财富人物(2017年9期)2017-11-02
