
芽孢杆菌蛋白酶基因的比较基因组学分析
2022-07-21 06:51贾仲昕赵佳男王承民秦建华
广东农业科学 2022年5期
贾仲昕,赵佳男,季 芳,王 雪,李 刚,王承民,秦建华
(1.河北农业大学动物医学院,河北 保定 071000;2.广东省科学院动物研究所/广东省动物保护与资源利用重点实验室/广东省野生动物保护与利用公共实验室,广东 广州 510260)
【研究意义】2020 年农业农村部全面禁止除中药外一切促生长类药物饲料添加剂的使用,标志着我国畜牧生产和养殖业“无抗时代”的到来。而蛋白酶一方面可以提高动物对蛋白质的消化并促进对养分的吸收,另一方面还可以有效减少进入后肠中的未消化蛋白数量,减少肠道菌群失衡的可能,从而保障动物肠道健康,因此蛋白酶成为抗生素促生长剂替代品的选择之一[1-2]。目前市场上的饲用蛋白酶主要由芽孢杆菌产生,因此探究影响芽孢杆菌产蛋白酶能力的重要基因显得尤为重要。【前人研究进展】芽孢杆菌产生的细胞外蛋白酶被认为参与细菌对营养物质的获取过程。研究发现,枯草芽孢杆菌(Bacillus subtilis)可以产生至少8 种独特的胞外或细胞壁相关蛋白酶[3],包括枯草杆菌蛋白酶AprE、中性金属蛋白酶NprE、丝氨酸蛋白酶Epr 和Vpr、杆状肽酶F(Bpr)、中性蛋白酶B(NprB)、细胞壁相关蛋白酶WprA 和金属蛋白酶Mpr。Yang 等[4]研究表明,蛋白酶基因缺失的唯一表型效应是蛋白酶活性的丧失。目前,芽孢杆菌蛋白酶基因的相关报道多集中于研究蛋白酶基因的调控机制以及高产蛋白酶突变株的诱导。Liu 等[5]发现degU和spo0A以及kinA的上调部分导致B.pumilus突变株SCU11 碱性蛋白酶的高产量。Zhou 等[6]研究发现,在地衣芽孢杆菌(B.licheniformis)中反应调节因子Spo0A 可能直接参与aprE的转录与激活。Zolfaghari 等[7]通过点突变等系统生物学方法,使aprE基因的表达水平和枯草杆菌蛋白酶E(Subtilisin E)的稳定性得到提高。【本研究切入点】随着测序技术的发展,细菌全基因组信息可以更加容易获得。通过比较不同菌种的基因组信息并结合细菌的相关表型,可以深入分析造成细菌表型差异的重要基因[8]。此外,芽孢杆菌全基因组注释信息的完善也为利用比较基因组学分析影响芽孢杆菌产蛋白酶能力的重要基因提供了研究基础。【拟解决的关键问题】本研究对15 株芽孢杆菌进行全基因组测序,通过检测15株芽孢杆菌的产蛋白酶能力,利用比较基因组学,挖掘影响芽孢杆菌产蛋白酶能力的重要基因,分析芽孢杆菌基因组中胞外蛋白酶基因与产蛋白酶能力的关联性,为产蛋白酶芽孢杆菌的开发提供依据。
1 材料与方法
1.1 试验材料
供试15 株菌株来源于广东省科学院动物研究所,具体如下:
枯草芽孢杆菌(B.subtiliis)N1282-4at、N1108-5at、N1303-2Ay,贝莱斯芽孢杆菌(B.velezensis)C1,蜡状芽孢杆菌(B.cereus)N435-1,高地芽孢杆菌(B.altitudinis)N435-3,巨大芽孢杆菌(B.megaterium)N447-3、B1,假真菌样芽孢杆菌(B.pseudomycoides)N451-2,海洋芽孢杆菌(B.oceanisediminis)N497-ZHONG,阿耶波多氏芽孢杆菌(B.aryabhattai)B2,副短短芽孢杆菌(Brevibacillus parabrevis)B3,芽孢杆菌(Bacillussp.)N470-1、N447-1、A1。
培养基配方参照文献[9-10]并稍作优化。蛋白酶发酵培养基配方(g/L):酪蛋白10.00、KH2PO40.360、NaH2PO4·2H2O 0.360、MgSO4·7H2O 0.500、FeSO4·7H2O 0.002、CaCl2·2H2O 0.480、NaCl 5.00。筛选培养基配方(g/L):酪蛋白10.0、牛肉膏粉3.0、Na2HPO42.0、NaCl 5.0、琼脂15.0、溴麝香草酚蓝0.05。MHB培养基配方(g/L):酪蛋白水解物17.5、牛肉浸粉5.0、淀粉1.5。NA 培养基配方(g/L):蛋白胨10、牛肉膏粉3.0、NaCl 5.0、琼脂17.0。
1.2 芽孢杆菌产蛋白酶能力检测
1.2.1 产蛋白酶菌株初筛 取出保藏的细菌冻存液接种于NA 培养基上,35 ℃培养24 h,挑取单菌落接种至MHB 培养基中,32 ℃、160 r/min 振荡培养24 h。将菌液梯度稀释,吸取100 μL 涂布于酪蛋白琼脂上,35 ℃培养48 h。用游标卡尺测量水解圈直径(H)和菌落直径(C)并计算H与C 的比值H/C。
1.2.2 蛋白酶活性测定 挑取初筛时酪蛋白琼脂上的单菌落,接种至MHB 培养基中,32 ℃、160 r/ min 振荡培养24 h 后,将菌液按2%(V/V)接种至发酵培养基中,32 ℃、160 r/min 振荡培养24 h,调整菌液浓度OD600为0.500,9 000 r/min离心15 min,上清液即为粗酶液。蛋白酶活性测定参考GB/T 23527-2009福林酚法并做适当修改。测定前先将粗酶液与酪蛋白溶液于40 ℃水浴预热5 min。
(1)试验组:取1 mL 粗酶液加 入1 mL 10 g/L 酪蛋白溶液,40 ℃水浴10 min。之后立即加入3 mL 0.4 mol/L 三氯乙酸溶液终止反应。静置10 min 后12 000 r/min 离心10 min。吸取上清液1 mL,加入5.0 mL 0.4 mol/L 碳酸钠溶液和1 mL福林试剂使用溶液,40 ℃水浴显色20 min,用紫外分光光度计在680 nm 处测定样品的OD680。
(2)对照组:先加三氯乙酸使蛋白酶失活再加酪蛋白,其他操作同上。
计算酶活性(X):
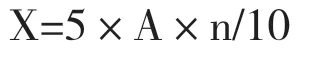
式中,X 为样品酶活性(U/mL),A 为由样品测得的净OD 值,查标准曲线所得相对应的酪氨酸含量,n 为酶液的稀释倍数,10 代表反应10 min,5 代表5 mL 反应液。
酶活性定义:在40 ℃下,每分钟内酪蛋白被水解产生1 μg 酪氨酸为1 个蛋白酶活性单位(U/mL)。
1.3 芽孢杆菌全基因组测序
将15 株菌株送至北京百迈客生物科技有限公司,基于Nanopore 测序技术平台进行全基因组测序。试验流程[11-12]按照ONT 公司提供的标准执行,包括样品质量检测、文库构建、文库质量检测和文库测序等流程。利用预测得到的基因序列与COG、KEGG、Swiss-Prot、TrEMBL、Nr 等功能数据库进行BLAST 比对,得到基因功能注释结果。
1.4 芽孢杆菌蛋白酶基因分析
查找芽孢杆菌中主要报道的8 种特征化细胞外或细胞壁相关蛋白酶[13](AprE、NprE、Bpr、Epr、Vp、NprB、WprA、Mpr),以及从细菌全基因组数据注释结果中发现的丝氨酸蛋白酶(Serine protease,AprX[14-15])的相关基因数据,包括菌株基本信息,蛋白酶基因的种类、数量和转录方向,蛋白酶基因间的相对距离等。利用IBS 线上作图工具(http://ibs.biocuckoo.org)进行 绘图。从全基因组数据结果中获取菌株的16S rDNA 序列,结合NCBI 数据库中已有的数据,利用MEGA X 软件绘制系统发育树。
1.5 菌株蛋白酶基因的 mRNA 水平检测
按照菌株产蛋白酶的相关表型,选取菌株B.altitudinisN435-3、B.oceanisediminisN497-ZHONG、B.velezensisC1、B.subtiliisN1282-4at进行蛋白酶基因的mRNA 水平检测。根据细菌相应蛋白酶基因序列,由生工生物工程(上海)股份有限公司设计与合成菌株引物(表1)。提取上述菌株在不同培养基(NB 培养基和发酵培养基)条件下的总RNA,以总RNA 为模板反转录为cDNA,以cDNA 为模板进行qPCR,16S rDNA为内参基因。
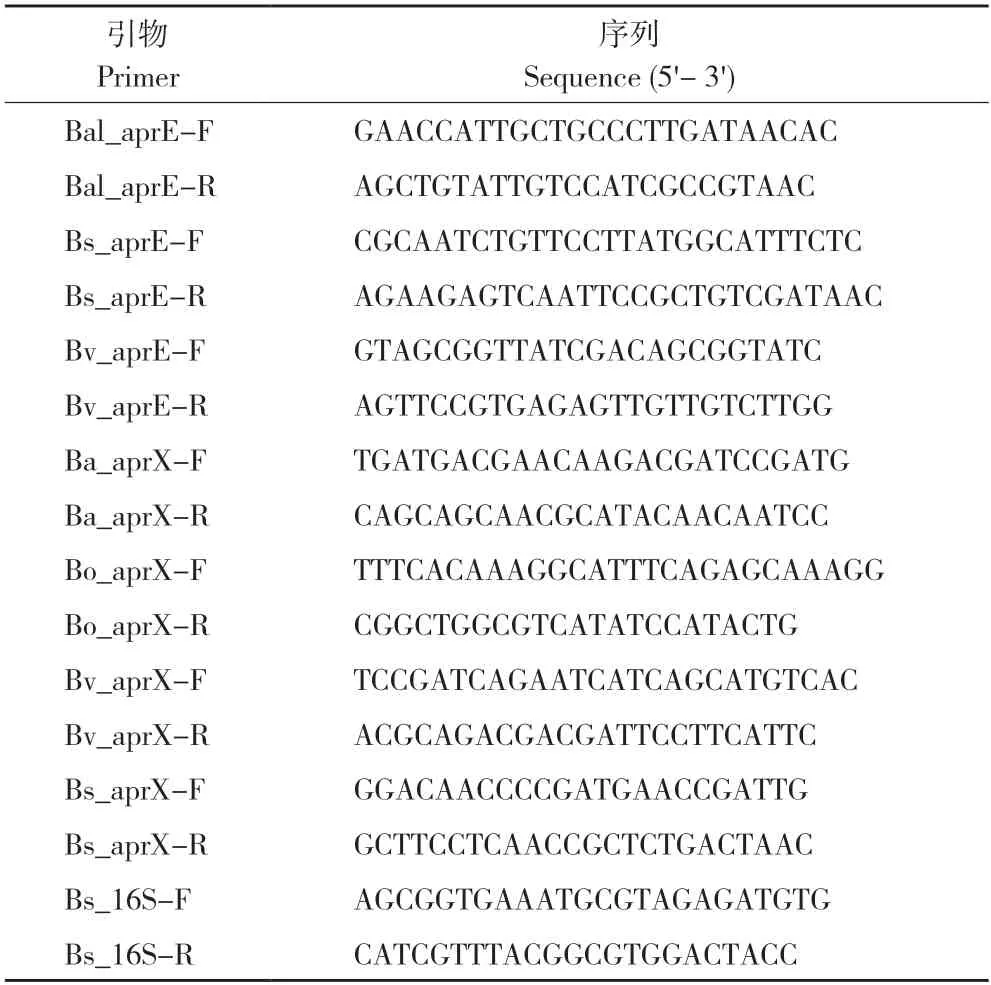
表1 菌株引物Table 1 Sequences of primers for strains
试验数据采用SPSS 19.0 软件进行单因素方差分析(One-way ANOVA)。
2 结果与分析
2.1 Bacillus sp.A1 和B.velezensis C1 产蛋白酶能力较强
在选取的15 株菌株中,有6 株菌株(Bacillussp.A1、B.velezensisC1、B.altitudinisN435-3、B.subtiliisN1282-4at、B.subtiliisN1108-5at、B.subtiliisN1303-2Ay)在接种培养 48 h,菌株透明圈直径与菌落直径的比值(H/C)显著高于其他9 株菌株。Bacillussp.A1 和B.velezensisC1的H/C 值最高,其次为B.subtiliisN1282-4at、B.subtiliisN1108-5at、B.subtiliisN1303-2Ay 和B.altitudinisN435-3(图1)。由于H/C 值在一定程度上反映了细菌产蛋白酶能力[16],因此上述6株菌株的产蛋白酶能力相对较强。

图1 产蛋白酶菌株初筛的H/C 值Fig.1 H/C value of primary screening of protease production strains
由图2 可知,15 株菌株中蛋白酶活性最高的2株菌株分别为Bacillussp.A1(31.40±0.40 U/mL)和B.velezensisC1(29.75±0.43 U/mL ),蛋白酶活性均达到 30 U/mL,显著高于其他13 株菌株;其次为B.subtiliisN1282-4at、B.subtiliisN1108-5at、B.subtiliisN1303-2Ay 等3 株菌株,蛋白酶活性在7.40~8.57 U/mL 之间,三者差异不显著;B.megateriumN447-3 和B.megateriumB1,蛋白酶活性仅为5 U/mL 左右 ;其他8 株菌株的蛋白酶活性均小于2 U/mL。菌株在不同培养基条件下酶活性不同,使用发酵培养基(底物酪蛋白为唯一氮源)后,15 株菌株的中性蛋白酶活性普遍有所提升,但菌株间蛋白酶活性的相对大小并未发生改变。
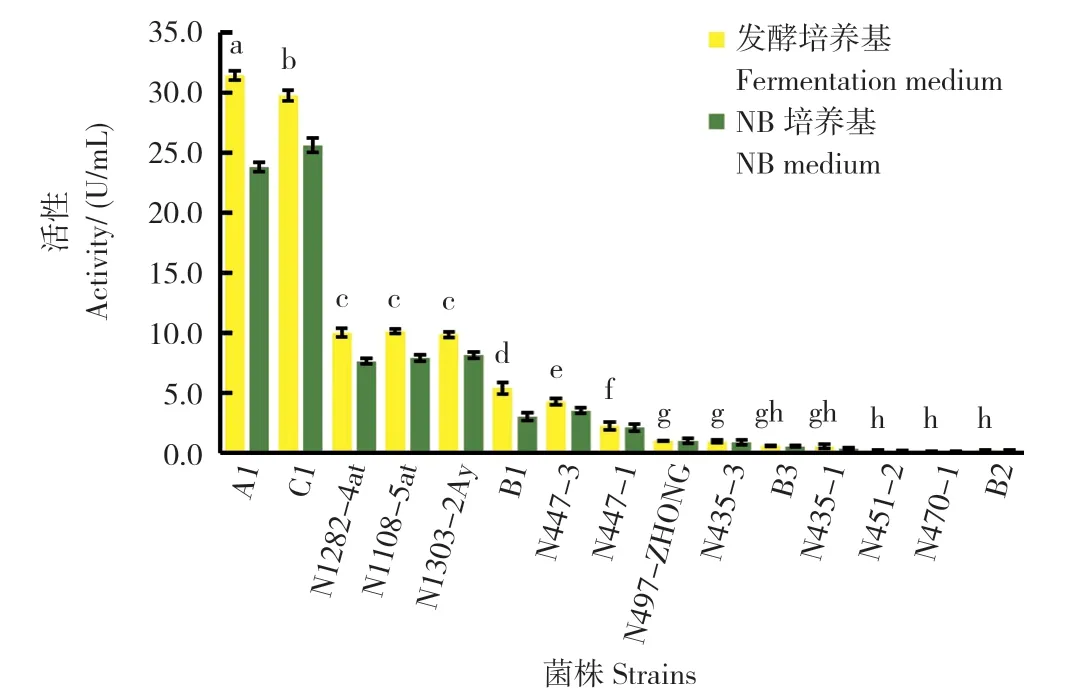
图2 菌株在不同培养基条件下的蛋白酶活性检测结果Fig.2 Detection results of protease activity of strains under different medium conditions
2.2 不同菌种携带不同的胞外蛋白酶相关基因组合
通过对菌株全基因组上蛋白酶相关基因的筛查与统计,芽孢杆菌属细菌携带有8 种胞外蛋白酶相关基因,包括aprE、aprX、nprE、epr、vpr、nprB、wprA和mpr。对上述基因的种类、转录方向、序列长度和染色体组中相对位置进行分析,可将其分为7 种基因型(G1~G7,图3)。3 株B.subtiliis菌株N1282-4at、N1108-5at、N1303-2Ay 属于G1 型,特点是染色体组上携带有上述8 种胞外蛋白酶基因。Bacillussp.A1、B.velezensisC1 属于G2 型,染色体组上携带aprX、vpr、epr、mpr。B.altitudinisN435-3 属于G3 型,相比G1 型缺少mpr、nprB和nprE。B.oceanisediminisN497-ZHONG 属于G4 型,染色体组上携带aprX、vpr、nprB。B.megateriumN447-3、B.megateriumB1、B.aryabhattaiB2 属于G5 型,染色体组上仅携带wprA、vpr,但同时存在2 个vpr。G6 型可分为3 个亚型G6.1、G6.2 和G6.3,其中B.cereusN435-1 属于G6.1 型,菌株染色体组上携带vpr、nprB和mpr;BacillusN447-1属于G6.2型,相比G6.1型缺少mpr;而BacillusN470-1 则属于G6.3 型,染色体组上仅携带vpr。B.pseudomycoidesN451-2 属于G7 型,染色体组上仅携带nprB。菌株Brevibacillus parabrevisB3 的染色体上则没有携带上述8 种基因。
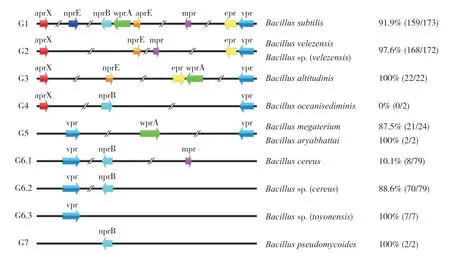
图3 芽孢杆菌属菌株蛋白酶的基因型分类结果F ig.3 Genotype classification results of Protease in Bacillus strains
通过与NCBI 数据库现有芽孢杆菌属细菌的全基因组数据(截至2021 年11 月21 日)进行比对,结果(图3)发现,在NCBI 数据库有173 株B.subtiliis具有全基因数据,其中159 株符合G1型基因分布(91.9%,159/173);B.velezensis有97.6%(168/172)符合G2 型;B.altitudinis100%(22/22)符 合G3 型;没 有B.oceanisediminis符合G4 型,考虑到NCBI 数据库中仅有2 株B.oceanisediminis的全基因组数据,因此存在较大偶然性;B.megaterium有87.5%(21/24)、B.aryabhattai有100%(2/2)符 合G5 型;而B.cereus中仅有10.1%(8/79)符合G6.1 型,更多的是88.9%(70/79)符合G6.2 型,B.toyonensis100%(7/7)符 合G6.3 型;B.pseudomycoides100%(2/2)符合G7 型。
2.3 细菌蛋白酶基因分型与其进化关系存在关联
绘制基于细菌16S rDNA 序列的系统发育树(图4),15 株菌株(红色)可分为Bacillus和Brevibacillus两个属,其中14 株属于Bacillus,1 株属于Brevibacillus。Bacillus属可分为3 个分支,其中G1、G2、G3 型所属菌株的亲缘关系接近为一个分支(绿色),G4、G5 型所属菌株的亲缘关系接近为一个分支(黄色),G6、G7 型所属菌株的亲缘关系接近为一个分支(蓝色)。B revibacillus parabreviB3 作为一个单独的分支出现,表明其与其他细菌物种的亲缘关系较远。通过与细菌的蛋白酶基因分型(图3)进行比对可发现,菌株与枯草芽孢杆菌亲缘关系越近,其染色体上携带的胞外蛋白酶基因种类越齐全。

图4 基于细菌16S rDNA 基因序列的系统发育树Fig.4 Phylogenetic tree based on 16S rDNA sequences of strains
2.4 芽孢杆菌蛋白酶基因分析
结合菌株的基因型分型结果(图3)与产蛋白酶能力检测结果(图2)发现,mpr、epr、vpr、nprB和wprA等5 个蛋白酶基因在产蛋白酶能力高和产酶能力低的菌株中均有携带情况。nprE基因仅有G1 型菌株携带,但其产酶能力低于不携带nprE的G2型。存在aprX和aprE的G1 型和G2 型菌株,其H/C 值和蛋白酶活性也均高于其他菌株,而也携带aprX和aprE的G3 型和携带aprX的G4 型菌株虽然其蛋白酶活性较低,但在酪蛋白琼脂上的H/C 也是高于除G1 型和G2型菌株外的其他菌株。G1~G4 型菌株的共同特征是携带aprX或aprE基因,因此本研究将影响菌株产蛋白酶能力的重要基因初步确定为aprE和aprX。
2.5 不同菌种间蛋白酶基因aprX 表达差异显著
为了探究影响菌株产蛋白酶能力的重要基因是否为aprX和aprE,本研 究检测了B.velezensisC1、B.subtiliisN1282-4at、B.altitudinisN435-3、B.oceanisediminisN497-ZHONG在不同培养基(NB 培养基和发酵培养基)条件下aprX、aprE的mRNA 水平。在N B 培养基(图5A,以B.subtiliisN1282-4at 作为对照),B.velezensisC1 的aprXm RNA 相对水平显著高于其他3 株菌株,为B.subtiliisN1282-4at 的2.7倍左右;B.altitudinisN435-3 则仅为N1282-4at 的60%;而B.oceanisediminisN497-ZHONG 则无表达或相比B.subtiliisN1282-4at 表达量过小。在发 酵培养基(图5C),相比NB 培养基,B.velezensisC1 的aprXmRNA相对水平上升为B.subtiliisN1282-4at的3.6倍左右;B.altitudinisN435-3 也有所升高,为B.subtiliisN1282-4at 的80%;而B.oceanisediminisN497-ZHONG 仍无表达或相比B.subtiliisN1282-4at 表达量过小。由图 5B、图5D 可见,发酵 培养基中B.altitudinisN435-3 的aprE相对表达水平为B.subtiliisN1282-4at 的约1.2 倍,差异显著,其余差异均不显著。根据 结果推测,aprX是影响芽孢杆菌产蛋白酶能力的重要基因。

图5 不同培养基菌株aprX 和aprE 的相对表达水平Fig.5 Relative expression levels of aprX and aprE under different medium conditions
3 讨论
本研究通过对15 株菌株进行全基因组测序,获得了菌株的全基因组信息,并根据菌株染色体上携带的蛋白酶基因数量、种类、转录方向将其分为7 种基因型,并进行比较基因组学分析。在15 株菌株中仅有G1 型的B.subtiliis拥有最全的8种蛋白酶基因,其他菌株都缺乏多种蛋白酶基因。通过对NCBI 数据库中已有菌株全基因组信息的比对,发现不同来源的相同菌种绝大多数归为同一基因型,可以认为在芽孢杆菌属不同来源的相同菌种间,蛋白酶基因携带情况保守。
有研究指出,枯草芽孢杆菌的主要胞外蛋白酶分别是aprE编码的枯草杆菌蛋白酶和nprE编码的中性金属蛋白酶[17-18]。但本研究结果显示,仅G3 型菌株B.altitudinisN435-3 的aprEmRNA水平在发酵培养基条件下是G1 型菌株B.subtiliisN1282-4at 的约1.2 倍,其他情况差异不显著。而nprE仅G1 型菌株携带,但缺乏nprE的G2 型菌株的产蛋白酶能力高于G1 型菌株。可能是因为G1 型菌株的nprE表达水平过低使NprE 的活性没有表现出来,同时aprE表达水平与G2 和G3 型细菌处于同一水平,而G2 型的aprX表达水平却显著高于G1 型菌株所致。
芽孢杆菌属菌株通常在指数生长期后期开始大量产生胞外蛋白酶,这使得细菌可以高效利用外界的营养物质,从而确保其正常的生长发育[19]。控制芽孢杆菌胞外蛋白酶表达的调节途径非常复杂,如AbrB、SinR 和ScoC 等调节因子可以抑制芽孢杆菌蛋白酶基因的表达,但Spo0A 可以抑制AbrB 和SinR 的活性从而促进蛋白酶的产生[19]。此外,aprE和nprE还受到全局转录调节因子CodY 的强大且直接的负反馈调节,但由于CodY同时也抑制ScoC[20],因此CodY 除了是aprE和nprE的直接负调节因子外,还通过抑制scoC的高水平表达充当相同基因的间接正调节因子。由于CodY 和ScoC 都是aprE和nprE的强阻遏物,并且CodY 会在含有过量氨基酸的培养基中具有活性,但随着氨基酸耗尽会失去活性[21]。考虑到G1 型和G2 型菌株的进化关系接近,这或许可以解释为何G1 型细菌的产蛋白酶能力低于G2型细菌。本研究在细菌生长的后期阶段检测了细菌产蛋白酶的能力和基因的表达,而此时营养物质的消耗导致CodY 的部分失活,从而减轻了对ScoC 的抑制,使ScoC 水平升高,持续抑制aprE和nprE的表达。因此结合荧光定量PCR 结果,aprE和nprE由于CodY 的失活导致对ScoC 抑制的减弱,从而受到ScoC 的强抑制,使细菌aprE和nprE的mRNA 水平降低。因此除aprE和nprE外,aprX是影响芽孢杆菌属细菌产胞外蛋白酶能力的又一重要基因。
蛋白酶是一种诱导酶,其在细菌内部的生物合成受到多种因素诱导(如蛋白酶底物和底物类似物),同时也会受到细菌生存环境中易利用氮源(如氨基酸和铵盐等)的阻遏[22-23]。因此,当外界环境中存在大量营养物质时,细菌只会产生少量蛋白酶来维持必要的生理活动,只有在外界环境缺乏易利用氮源时,细菌才会大量产生蛋白酶,去获取环境中其他难利用的大分子蛋白质。这也解释了为何细菌在发酵培养基(唯一氮源为酪蛋白)条件下aprX和aprE的表达水平比在营养肉汤培养基条件下高。
4 结论
本研究对15 株芽孢杆菌进行全基因组测序获得了菌株的全基因组信息,并根据菌株携带的蛋白酶基因情况进行了基因分型,通过对NCBI数据库中已有菌株全基因组信息的比对,发现芽孢杆菌不同来源的相同菌种间蛋白酶基因携带情况保守。通过构建进化树并与基因分型结果进行比对,发现芽孢杆菌属菌株与枯草芽孢杆菌亲缘关系越近,携带的胞外蛋白酶基因种类越齐全。通过对15 株芽孢杆菌进行产蛋白酶能力检测,并进行比较基因组学分析和qRT-PCR 验证,发现aprX是影响芽孢杆菌属细菌产蛋白酶能力的重要基因。
猜你喜欢
食品科学(2022年16期)2022-09-01
江西水产科技(2022年2期)2022-05-17
江西水产科技(2022年2期)2022-05-17
浙江农业学报(2022年1期)2022-02-18
大科技·百科新说(2020年4期)2020-06-19
食品安全导刊·中旬刊(2020年2期)2020-06-01
文苑(2018年22期)2018-11-19
百科知识(2018年20期)2018-10-18
环球市场(2018年19期)2018-09-10
食品与生活(2018年10期)2018-01-15
