
小相岭山系非飞行小型兽类物种多样性海拔分布格局
2022-07-28 08:26刘芝秀周恩华张勘王大勇胡杰李艳红吉晟男
四川动物 2022年4期
刘芝秀,周恩华,张勘,王大勇,胡杰*,李艳红*,吉晟男
(1.西华师范大学西南野生动植物资源保护教育部重点实验室,四川 南充 637009;2.四川栗子坪国家级自然保护区管理局,四川 石棉 625400;3.四川冶勒自然保护区管理处,四川 冕宁 615600;4.中国环境科学研究院,北京 100012)
小型兽类因种类多、分布广、适应能力强,在生态系统中占据着重要地位(秦岭等,2007),同时在维持食物链、食物网、营养循环以及生态系统的结构和功能方面也发挥着重要的作用(Liu,2009)。另外,小型兽类迁移性小,对环境变化敏感且种群数量稳定(Umetsu,2008),其群落结构的变化不仅可以反映生境变化,还可以反映人类活动的干扰程度,是评估环境质量的良好实验对象(傅琰华等,2015)。非飞行小型兽类(简称小型兽类)主要是指除翼手类外的所有体型较小、体质量较轻的哺乳动物。本文研究的对象主要是劳亚食虫目Eulipotyphla、啮齿目Rodentia和兔形目Lagomorpha的物种。
物种多样性包括α、β和γ 3个不同的空间尺度,相互影响、相互作用、不可分割。在对物种多样性海拔空间分布格局的研究上,α、β和γ多样性出现并驾齐驱的趋势(权寿瑛等,2009;吴永杰等,2012;Krasnov,2018)。全球物种丰富度海拔梯度分布格局主要包括4种模式,即单调递减模式、前峰模式、驼峰模式和单调递增模式(Rahbek,2005)。在小型兽类中,前峰模式和驼峰模式更普遍(Rahbek,2005;McCain,2006)。我国大多数学者对小型兽类海拔梯度格局的研究也表明,物种丰富度在中海拔地区最高(权寿瑛等,2010;王作波等,2020)。相邻海拔之间的物种更替地理格局则表明,多峰和不规则模式最普遍(Tang,2012;Wang,2012)。目前,对小型兽类多样性海拔分布格局形成机制的探讨尚未有定论,因此需要在更多的地区开展相关研究。
小相岭山系位于全球36个生物多样性热点区域之一的西南山地(Habel,2019),区域内植被类型丰富、动植物种类繁多,在生物多样性保护和维持方面具有重要的研究价值。迄今为止,该区域内物种多样性的相关研究着重于大中型兽类和鸟类(李健威等,2020),小型兽类仅有简单的描述性报道(张君,胡锦矗,2004;李艳红等,2007)。为此,在小相岭山系开展有关小型兽类的研究,一方面可以进一步丰富该区域小型兽类多样性的基础信息,另一方面,可以探讨小相岭山系小型兽类的海拔梯度分布格局,进而为该区域小型兽类生物多样性的保护和管理提供科学依据。
1 研究地区和研究方法
1.1 研究区概况
小相岭山系属大雪山支脉,面积约11 500 km,介于石棉、冕宁、越西、喜德和九龙5县交界处(图1),境内地形崎岖,山岭重叠,地势西高东低,地貌切割破碎强烈,以中山和高山峡谷为主,南北长约147 km,东西宽约107 km,总体呈南北走向,且北高南低(冉江洪等,2004;胡锦矗,吴攀文,2007)。小相岭山系由焦顶山、铧头尖、窝洛几峨等山峰组成,主山脊海拔大多在4 000 m以上,最高峰为则尔山,海拔5 299 m,最低海拔处于大洪村板栗树沟(1 330 m),海拔平均相对高差2 000 m以上(Hu,2010)。山系四周由于高山屏障常产生强烈的焚风效应,且有明显的干湿季节变化,属于以亚热带季风为基带的山地气候,冬春多风,夏秋多雨,光照少,年均日照率为30%左右。由于地势原因,“雨屏效应”显著,年均降水量800~1 250 mm,年均气温11.7~14.4℃,年均相对湿度87.9%,垂直变化明显(青菁等,2016)。

图1 小相岭山系样方布设Fig.1 Sample sites in the Xiaoxiangling Mountains
小相岭山系植被类型丰富多样,发育形成了完整的植被垂直带谱:海拔1 330~2 400 m为常绿阔叶林,山楠、曼青冈、巴东栎、卵叶钓樟等为优势种;海拔2 400~2 700 m为落叶阔叶林,野核桃细叶青冈槲栎、华西枫杨、大叶杨等为优势种;海拔2 500~2 700 m为针阔叶混交林,扇叶槭、红桦、铁杉、麦吊云杉等为优势种;海拔2 700~3 700 m为亚高山针叶林,铁杉、麦吊云杉、川滇冷杉、冷杉等为优势种;海拔2 800~3 700 m为亚高山灌丛,大白杜鹃、小叶栒子、四川嵩草等为优势种;海拔3 700~4 400 m为高山灌丛草甸,凝毛杜鹃、金露梅、峨眉蔷薇、高山嵩草、高原毛茛等为优势种;海拔4 400 m以上为高山流石滩稀疏植被,风毛菊、水母雪莲花、梭砂贝母等为优势种(刘守江等,2011)。
1.2 试验设计
2019年7—8月和2020年7—9月,根据小相岭区域的生境类型和地形地貌特征等因素,采用夹日法对该区域内的小型兽类进行野外调查。选取海拔1 300~4 100 m为研究区域,每400 m设置样方,在7个连续海拔段共设置53个样方。样方间距大于100 m,并尽可能覆盖所有生境类型。样方大小为100 m×100 m,夹线行距25~30 m,夹距3~5 m,每个样方放置66~88个鼠夹。以新鲜花生米为诱饵,每日下午放置,翌日上午收夹,连捕3 d。收夹时,收集捕获动物并补放诱饵,同时对捕获到标本的鼠夹进行清洗和烘干。将捕获的标本进行编号、称重和常规测量(包括头体长、尾长、吻长、耳长和后足长),鉴定性别和年龄段并记录数据。所有标本用99%无水乙醇浸制,带回实验室鉴定。
1.3 数据处理
小型兽类物种鉴定主要参考《中国兽类野外手册》(Smith,解焱,2009)和《四川资源动物志(第二卷)》(胡锦矗,王酋之,1984);分类系统参考《中国哺乳动物多样性(第2版)》(蒋志刚等,2017)。
通过物种累积曲线判断野外调查是否满足取样需求(Ugland,2003)。
物种丰富度用观察到的物种数表示,插值物种丰富度指的是如果一个物种在较低和较高海拔都有记录,那么就认为该物种在这2个海拔之间的所有区域都有分布。Chao2丰富度作为另外一种矫正指数,其计算原理是将实际捕获的物种数通过非参数的统计方法,估算出总的物种数量(Colwell&Coddington,1994)。物种多度为每个样方的个体数之和,相对多度由物种捕获率表示。
群落多样性指数采用Shannon-Wiener多样性指数(′)和Simpson多样性指数();优势种判断采用Berger-Parker优势度指数();群落相似性采用Chao-Sørensen相似性指数()。利用一元线性方程评估随海拔距离增加的衰减率,斜率的绝对值表示衰减率,值越大表示物种相似性随海拔距离的增加衰减越快,说明该地区β多样性越高(Soininen,2007)。

式中,为物种数,P为种的个体数占全部个体数的比例,N为优势种的个体数,为群落中全部物种的个体数,分别为2个群落各自的物种数,为2个群落共有物种数。
所有数据利用Excel 2016、SPSS 21.0和EstimateS 9.0进行处理分析。
2 结果
2.1 物种组成
累计布设11 902个夹日,捕获小型兽类标本317只,隶属于3目7科15属23种,总捕获率为2.7%。其中,劳亚食虫目3科(鼩鼱科Soricidea、鼹科Talpidae、猬科Erinaceidae)5属6种;兔形目鼠兔科Ochotonidae 1属2种;啮齿目3科(鼠科Muridae、松鼠科Sciuridae、仓鼠科Cricetidae)9属15种。啮齿目数量最多,有225只,占总捕获量的71.0%;劳亚食虫目和兔形目较少,分别占18.3%和10.7%。在物种组成上,优势种为康定绒鼠(18.3%)、中华姬鼠(12.9%)和高山姬鼠(10.1%)。不同物种具有不同的海拔分布区:分布在2 000 m及以上的4种,分别是高山姬鼠、中华姬鼠、大耳姬鼠和川西白腹鼠;其次是分布在1 500 m左右的微尾鼩、小纹背鼩鼱、康定绒鼠和藏鼠兔(表1)。以上物种为宽域分布物种(海拔分布范围超过研究区域的一半),其余物种为狭域分布。

表1 小型兽类物种组成及海拔分布Table 1 Species composition and elevational distribution of small mammals
结合文献资料(张君,胡锦矗,2004;李艳红等,2007;李健威等,2020)和调查访问情况,小相岭山系记录小型兽类45种,隶属于3目8科28属。其中,劳亚食虫目3科12属16种,占总物种数的35.6%;啮齿目4科15属27种,占60.0%;兔形目1科1属2种,占4.4%。中国特有种16种,占35.6%。列入《中国生物多样性红色名录(第一卷哺乳动物)》(蒋志刚,2021)的濒危(EN)物种1种,即滇攀鼠;易危(VU)物种2种,为宽齿鼹和峨眉鼩鼹;近危(NT)物种4种,为白尾鼹、纹背鼩鼱、中华绒鼠和康定绒鼠。新增分布记录物种1种,即滇攀鼠。
2.2 物种丰富度取样评估
在捕获了15种物种后,物种累积曲线由直线上升变为舒缓上升,逐渐成一渐近线(图2),说明本次野外调查取样较为充分,可以满足后续的处理分析需要。

图2 物种累积曲线Fig.2 Species accumulation curves
2.3 物种多样性海拔分布格局
本研究使用3种不同的物种丰富度指数验证小相岭山系小型兽类的海拔分布格局,均得到一致的研究结果,即物种丰富度在海拔2 500~2 900 m段最高。实际物种丰富度和插值物种丰富度的分布格局为单峰型,且变化趋势较为一致,都在达到峰值后,呈递减趋势。Chao2丰富度的海拔分布格局为双峰型,除了在海拔2 500~2 900 m段出现峰值外,在3 300~3 700 m也出现了峰值。另外,捕获率作为反映群落物种相对多度的指标,在中低海拔段变化明显,呈“W”形,随后在高海拔段缓慢上升,最高点出现在3 700~4 100 m处(图3)。

图3 小相岭山系的物种丰富度指数和物种多度海拔分布格局Fig.3 Elevational distribution patterns of species richness indices and species abundance in the Xiaoxiangling Mountains
小相岭区域小型兽类的生物多样性指数(Shannon-Wiener和Simpson多样性指数)最大峰值出现在中海拔段,即2 500~2 900 m处,与物种丰富度海拔分布格局相似。Chao-Sørensen相似性指数结果表明,低海拔区域(1 700~2 100 m vs.2 100~2 500 m)和高海拔区域(3 300~3 700 m vs.3 700~4 100 m)的物种相似性最高,达到0.93和0.95,而中海拔段的物种相似性较低。群落相似性随海拔的衰减率表明,小相岭山系整个海拔梯度小型兽类的物种更替随海拔增加而明显衰减(图4)。
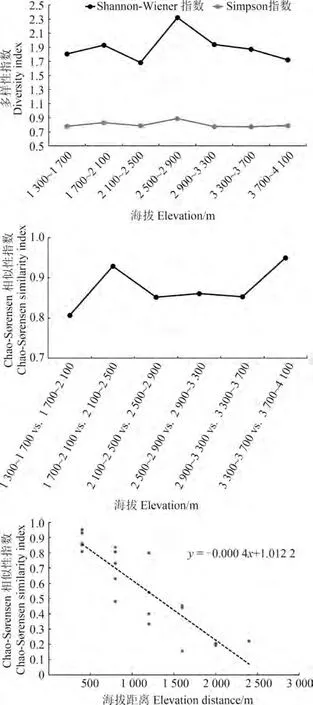
图4 小相岭山系物种多样性海拔分布格局Fig.4 Elevational patterns of species diversity in the Xiaoxiangling Mountains
2.4 不同海拔小型兽类群落组成分析
在1 300~1 700 m内捕获小型兽类9种,第一优势种为高山姬鼠(0.40),第二优势种为微尾鼩、北社鼠和川西白腹鼠(0.13);在1 700~2 100 m捕获9种,第一优势种为北社鼠(0.26),第二优势种为微尾鼩(0.22),第三优势种为中华姬鼠和高山姬鼠(0.15);在2 100~2 500 m捕获7种,第一优势种为中华姬鼠(0.32),第二优势种为微尾鼩(0.24),第三优势种为大耳姬鼠(0.21);在2 500~2 900 m捕获13种,第一优势种为中华姬鼠和高山姬鼠(0.17),第二优势种为微尾鼩和大耳姬鼠(0.11);在2 900~3 300 m捕获12种,第一优势种为康定绒鼠(0.43),第二优势种为中华姬鼠(0.13);在3 300~3 700 m捕获10种,第一优势种为康定绒鼠(0.43),第二优势种为小纹背鼩鼱(0.12);在3 700~4 100 m捕获8种,第一优势种为高原松田鼠(0.35),第二优势种为大耳姬鼠和藏鼠兔(0.17)(表2)。
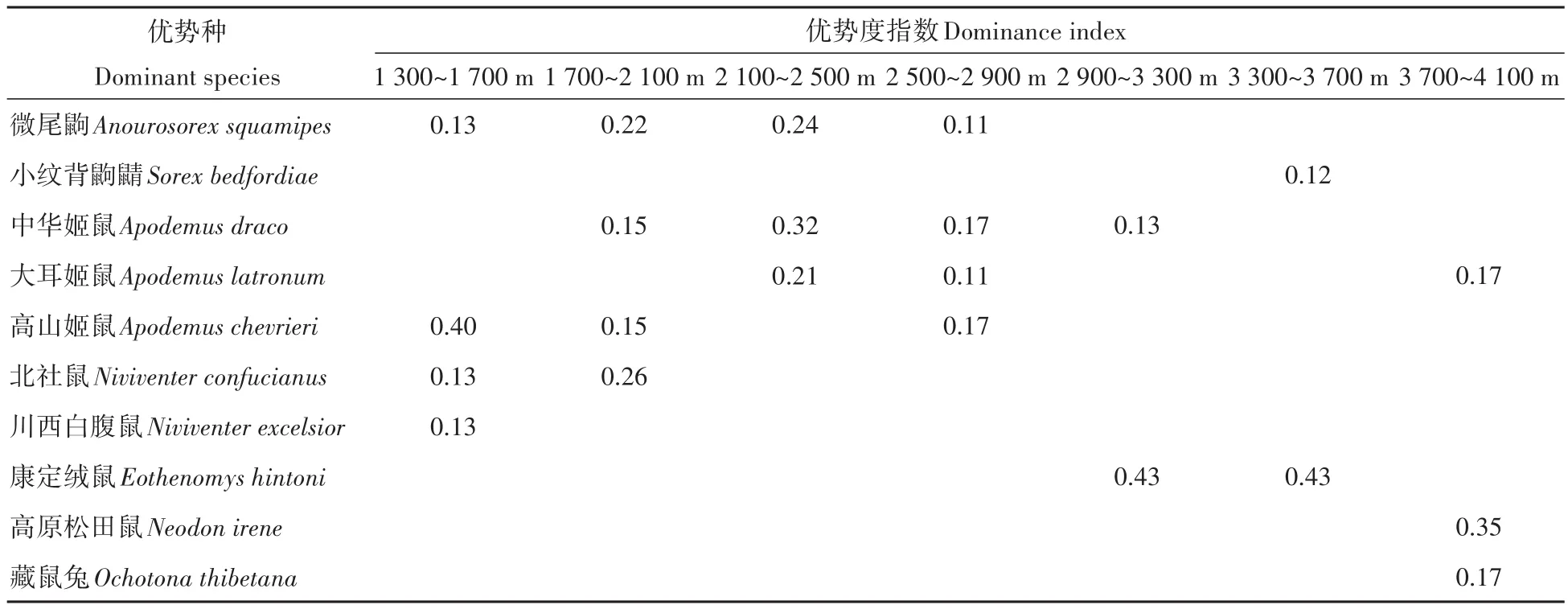
表2 不同海拔段小型兽类优势种组成Table 2 Composition of dominant species of small mammals at different elevations
3 讨论
3.1 物种组成
本次调查捕获小型兽类23种,与以往调查相比(张君,胡锦矗,2004),有22种小型兽类未被调查到,但新增新分布物种滇攀鼠,核实原有的兽类名录发现,部分物种的分布值得商榷,如根田鼠和黑唇鼠兔:根田鼠仅分布于新疆(刘少英等,2019),而黑唇鼠兔在四川仅分布于若尔盖、甘孜、德格、石渠等川西高原地区(王酉之,胡锦矗,1999)。陈鹏(2012)关于姬鼠属的研究表明,长尾姬鼠并非有效种,因此,此处的长尾姬鼠实为中华姬鼠。康定绒鼠原为西南绒鼠康定亚种,Liu等(2012)将该亚种提升为种。汪巧云等(2020)研究表明,喜马拉雅水鼩仅分布在云南,而四川等地的实为利安德水鼩。此外,Liu等(2019)通过分子学实验,对绒鼠属的几个亚种进行鉴定研究,认为石棉绒鼠和美姑绒鼠应为独立有效种。因此,本文对这些物种进行了删除或补充修订。部分物种可能因其数量稀少或栖息于特殊生境内,未被捕获,如林跳鼠s、蹼足鼩利安德水鼩等。
3.2 不同海拔的优势种
小相岭山系不同海拔的小型兽类优势种主要由高山姬鼠、微尾鼩、北社鼠、川西白腹鼠、中华姬鼠、大耳姬鼠、康定绒鼠、小纹背鼩鼱、高原松田鼠和藏鼠兔组成。其中,中华姬鼠和大耳姬鼠在所有海拔段内均有分布。广域分布种在不同海拔优势种中的占比高达80%,说明其具有很强的繁殖、生存和适应能力(张云智等,2005)。对大相岭山系小型兽类群落及生境的研究结果显示,广布种在群落优势种中占比较大,且优势种的种类(齐敦武,2005)与本研究结果较为相近,这表明大、小相岭山系的小型兽类可能具有相似的群落特征。随着海拔的升高,第一优势种出现由高山姬鼠、北社鼠替换成中华姬鼠,再到康定绒鼠和高原松田鼠的现象,而第二与第三优势种的变化无明显规律。四川黑竹沟地区(岳先涛等,2020)和云南农耕区(陈星等,2019)小型兽类群落组成的海拔变化也表明,优势种存在随海拔升高而发生替换的现象。说明海拔的变化是影响小型兽类群落结构组成的重要因素,但地形、气候和食物资源等其他因素的作用同样不可忽视(胡一鸣等,2018)。
3.3 小型兽类物种多样性海拔梯度分布格局
物种丰富度和物种多度作为反映物种生物多样性最直接的2个指标,常被用于预测山地物种的分布格局,但由于其对常见种和稀有种的评价存在一定的缺陷,通常将Shannon-Wiener和Simpson多样性指数综合起来以弥补这一不足(栗忠飞,高吉喜,2018)。本研究中,小型兽类的物种数明显在2 500~2 900 m最高,而物种相对多度的峰值则出现在2 900 m以上,二者的海拔梯度分布格局存在一定的差异。Shannon-Wiener和Simpson多样性指数独立于物种多度分布格局外,可以较为准确地评价常见种或稀有种的贡献率,本研究中,这2种多样性指数的海拔分布格局峰值都出现在海拔2 500~2 900 m段,因此将这2种多样性指数与物种丰富度相结合,可以更好反映物种的垂直分布变化情况。对西南山地小型兽类物种丰富度海拔梯度分布格局的研究结果表明,该区域小型兽类物种丰富度的海拔格局存在明显的空间变化,权寿瑛等(2009)发现横断山区100~5 000 m范围内的小型兽类物种丰富度海拔分布格局为宽峰型;马俊等(2010)发现螺髻山中海拔段的小型兽类物种多样性最高;吴永杰等(2012)发现贡嘎山东坡小型兽类的物种丰富度在中海拔地区最高。本研究结果与上述结论一致,小相岭山系的小型兽类物种丰富度在2 500~2 900 m段最高,符合中峰模式。这可能是由于西南山地2 500~3 000 m的中海拔地区位于最大降水带,该海拔范围内湿度大、植被类型多样、食物丰富,相对于低海拔和高海拔结构简单的生境,更利于小型兽类生存(张荣祖等,1997)。
3.4 小型兽类物种更替海拔梯度分布格局
对于物种β多样性的分布格局,既往的研究主要通过相邻海拔之间的物种更替和相似性距离衰减率2种不同空间尺度来分析:Mena和Vázquez-Domínguez(2005)发现鼠类的相邻海拔间物种更替海拔格局分布模式均为多峰与不规则型;Wen等(2014)对横断山区小型兽类的物种更替地理格局的研究同样表明,贡嘎山、唐家河和响古菁3条样线的相邻海拔间物种更替格局均不一致。本研究对小相岭山系海拔1 300~4 100 m段小型兽类相邻海拔间的物种更替格局分析发现,低海拔区域和高海拔区域的物种相似性最高,而中海拔段的物种相似性相对较低,说明中海拔段的物种更替更快、β多样性更高。Rapoport法则预测,物种组成在高海拔地区的变化小于低海拔地区(Stevens,1992,1996),高海拔地区存在的物种分布区域重叠导致β多样性随海拔的升高而降低。在更大尺度上,不同海拔间的物种相似性随海拔距离的增加而减小,表现为衰减率越大,β多样性越高(Soininen,2007)。小相岭山系小型兽类的相似性随海拔距离的衰减率比西南山地其他地区的(Wen,2014)更低,这可能是由于小相岭山系的群落优势种主要为广域分布种,扩散能力较强、物种聚集度较低,因而β多样性相对较低。
本研究得到四川栗子坪国家级自然保护区管理局和四川冶勒自然保护区管理处的大力支持和协助;感谢张德军、吴大鹏、李勇、陈红、魏勇等人在野外采集工作中提供的帮助。
猜你喜欢
华人时刊(2022年13期)2022-10-27
当代水产(2022年8期)2022-09-20
军事文摘(2022年14期)2022-08-26
中国音乐学(2022年2期)2022-08-10
华人时刊(2022年5期)2022-06-05
华人时刊(2022年3期)2022-04-26
散文诗(2021年22期)2022-01-12
当代陕西(2020年23期)2021-01-07
学校教育研究(2020年7期)2020-04-09
环球时报(2019-05-23)2019-05-23
