
珍稀濒危植物独花兰研究进展
2022-08-10 02:00刘旭阳王仕宝朱志凯
陕西农业科学 2022年6期
张 慧,刘旭阳,王仕宝,朱志凯,吴 鹏
( 1.汉中职业技术学院 药学院;陕西 汉中 723000;2.汉中职业技术学院 秦巴山区药(食)用植物研究所,陕西 汉中 723000;3.汉中市南郑区食品药品检验检测中心,陕西 汉中 723000 )
兰科独花兰(ChangnieniaamoenaS. S. Chien)是我国特有的单种属植物,其药用价值较高,在《全国中草药汇编》中记载:其药材名为长年兰,以假鳞茎或全草入药,其功效为清热、凉血、解毒,主治咳嗽,痰中带血,热疖疔疮[1]。独花兰的外部形态特殊、色彩美丽,全株只有一叶一花,叶片宽卵状椭圆形至宽椭圆形,一般长6.5~11.5 cm,宽5~8.2 cm,叶片上面绿色,背面紫红色。花葶顶生,长10~17 cm,紫色,花白色、带淡紫色或肉红色晕,唇瓣上有紫红色斑点[2]。近年来,由于环境的变化、过度的采挖及自身繁育困难等因素,导致独花兰资源急剧减少,1992年被《中国植物红皮书—稀有濒危植物( 第一册) 》列为珍稀濒危种,成为国家二级保护植物[3]。文章主要针对独花兰的分布区域、群落特征、繁育特征、组织培养育苗技术和遗传与变异五个方面方面进行综述,以期为独花兰的深入研究提供参考依据。
1 分布区域
查阅中国植物志[2]、FRPS图库照片分布图(图1)及相关文献[4~8],得出独花兰分布在13个省(市)区,主要以湖北、湖南等华中地区为中心,北至河南、陕西南部,南至浙江、江西及广西等地,西至四川,甘肃南部,东至安徽、河南一带。此外,还包括重庆、贵州等地。另有报道[9]:独花兰于1970年在陕西省平利县首次发现,之后四十多年再未发现,直到2011年和2012年分别在陕西化龙山国家级自然保护区内被重新发现。2018-2020年,进行中药资源普查期间,在陕西宁强县域发现多个独花兰野生分布群落[10]。
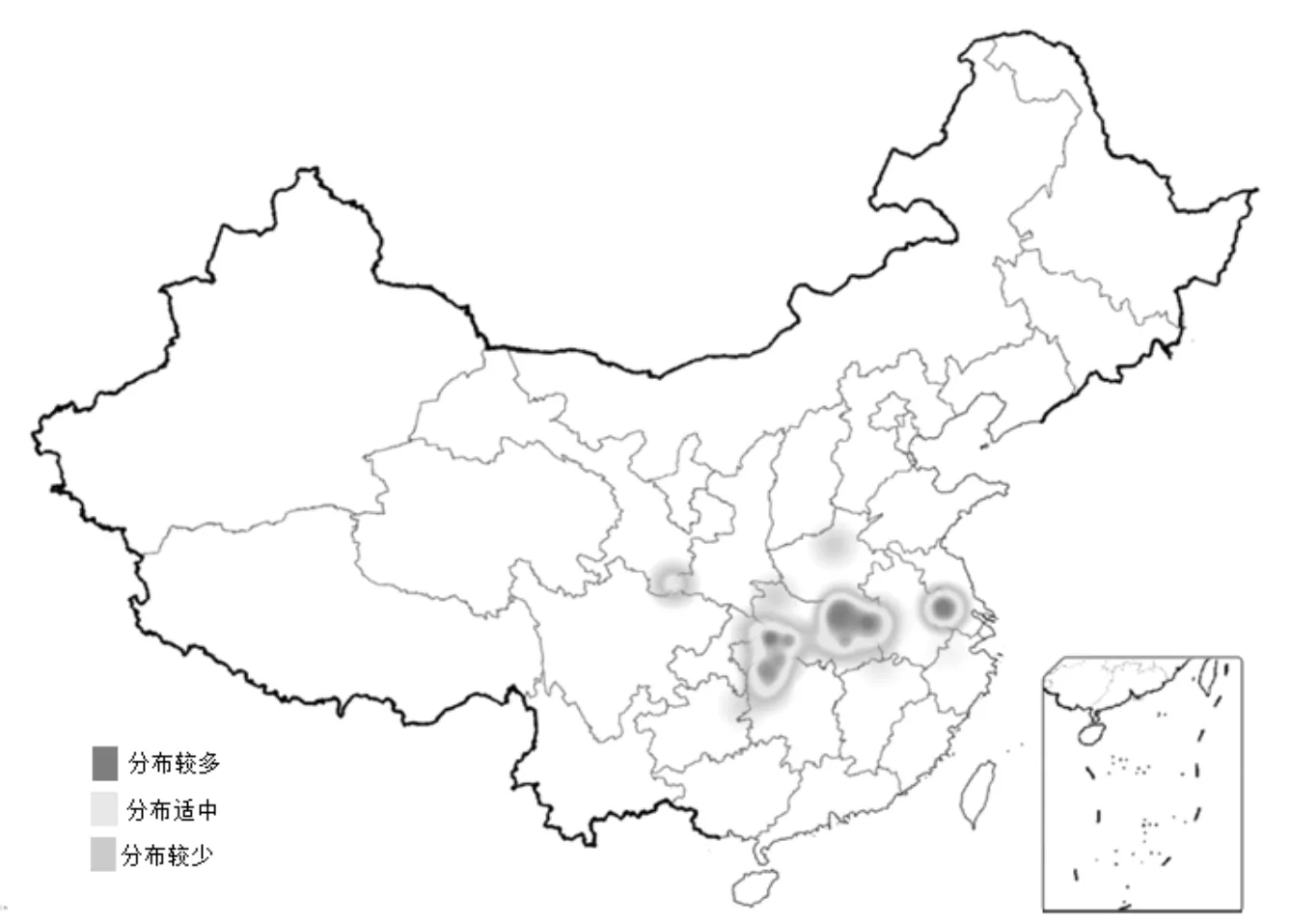
图1 FRPS图库照片分布
2 群落特征
生物群落特征主要包括群落中物种的多样性、群落的生长形式、结构、优势种等多个方面。独花兰群落一般分布于坡度15~45°的阴坡,海拔范围常为400~1 700 m,山地黄棕壤,郁闭度40%~75%,林下腐殖质丰富、水分充足的土壤,同时排水性良好。邬刚等基于邻接格子法和直接计数法,研究了英山县现有的168株独花兰的种群数量及分布格局,利用方差/均值比率法,得出草盘镇桃花冲村独花兰小种群为集群分布[11]。神农架南坡独花兰主要分布的伴生植物,包括以化香树、四照花、华千金榆等为主的落叶阔叶林及箬竹组成的灌丛下,北温带物种为群落中的优势物种[12]。鸡公山保护区独花兰主要分布在落叶阔叶林和杉木林中,主要植被有水杉、朴树、元宝槭、常春藤、络石、山胡椒、绞股蓝等[13]。贵州台江县独花兰的生境调查发现其生于针阔混交疏林下,主要乔木树种有杉木、金叶子、鹅掌楸、楠木等,主要灌木有栎属,主要草本有苔草属、百合属、虾脊兰、金兰等植物[14]。王仕宝等在陕西宁强县区域内落叶阔叶林或针阔混交林中发现了独花兰野生种群,其植被乔木层有壳斗科、松科及桦木科;灌木层主要有小檗科、山茱萸科、忍冬科、漆树科、冬青科、禾本科(竹亚科)等;草本层主要包括百合科、唇形科、兰科、鹿蹄草科、薯蓣科、禾本科(禾亚科)、龙胆科等植物[10]。
3 繁育特征
3.1 繁育与传粉
独花兰是自交亲和的虫媒花,正常情况下需借助前来吮吸花粘液分泌物的昆虫触及隐藏在药帽下的花粉或粘盘,将花粉块带到柱头凹陷处的腔室内完成授粉。虽然独花兰具有花距,但不分泌花蜜,不能为传粉者提供回报,是欺骗性传粉模式[15],并且独花兰花期在4月份,海拔较高区域温度较低,受访昆虫偏少,传粉者较少。研究表明在独花兰群落内出现的昆虫有15种,为其传粉的只有熊蜂。三条熊蜂是神农架龙门河群体传粉者,仿熊蜂是神农架关门山群体的唯一传粉者。可见,传粉者限制是导致自然结实率低的主要原因[16]。此外,结合文献[17],对独花兰的生理构造研究表明,花药帽较大,并且包盖紧密,不利于花粉块的外露和传播;花粉块柄不发达,不能将花粉块托起将花药帽顶开;粘盘不发达,无法将花粉块带出药帽。因此,整个花期内,如果没有昆虫的帮助,花粉块将一直包藏在花药帽下,无法正常完成授粉过程。独花兰的这些生物学缺陷导致了其只开花不结果,繁殖率低下。琚煜熙[12]跟踪调查鸡公山自然保护区内14个分布群的340株独花兰,发现其开花株数为215株,开花率为 63.2%,结果株数仅有12株,结实率仅为5.6%。桂先群[18]分别于2008年4月24日和2014年4月25日对天堂寨自然保护区内的独花兰进行人工授粉。结果显示,自花受粉后花被枯萎凋落,子房会伸长,果实成熟期结果,未受粉的花被鲜艳,子房不会伸长,果实成熟期不结果,表明独花兰是自交亲和的异交种。同时,该试验除进一步证实了独花兰的生物学特性是其致危的重要因素外,还对人工授粉技术提高其种群密度的应用提供了依据。
3.2 繁育与菌根
兰科植物作为最进化复杂的类群之一。在自然环境下,兰科植物的大多数种子极为细小,缺乏胚乳组织,需要和适宜的真菌共生才能萌发。独花兰作为兰科植物也不例外,种子的胚发育不完全,没有贮藏种子萌发所需的营养组织,在自然条件下,独花兰的种子不能正常萌发;当独花兰的种子与特定的真菌形成稳定的共生关系后,才使种子利用真菌提供的营养物质, 继而萌发并生长成植株。武静宇根据独花兰菌根真菌培养的特点及其形态学特征,初步将其菌根真菌鉴定为兰科共生丝核菌,后采用分子生物学方法,将其菌根真菌的rDNA ITS序列进行Blast同源比对,进一步得出该真菌属于胶膜菌属[19]。有文献报道独花兰具有典型的兰科植物菌根构造;分离得到的独花兰的菌根真菌主要属于担子菌和有丝孢菌的菌株,镰刀菌属为优势种属;利用回接试验法来判断从独花兰根中分离到的真菌是否为菌根真菌时发现大部分菌株对华石斛组培幼苗生长有不同程度的促进作用[20~21]。基于此,通过筛选适合的真菌进行独花兰种子的共生萌发试验,为有效实现独花兰植物的种质保育,进行濒危野生居群的生态恢复显得尤为重要。
3.3 繁育与营养体
花的发育和开花对物种繁殖具有重要意义。对于多年生植物, 其生殖生长需要建立在若干年的营养生长的基础之上。因此,植株营养生长状况影响着花的发育和开花。独花兰的地下部生长有假鳞茎,假鳞茎作为一种变态茎,是独花兰植物体的重要营养器官,每个假鳞茎的顶端生有1个叶片,并且是花芽着生的地方。独花兰叶片大小能够体现植株个体大小,反映植株所处的发育阶段。叶片大小与地下茎体积和假鳞茎数目呈高度正相关,同时,依据叶片的大小、假鳞茎的数量及地下茎的体积大小可以判断独花兰是否达到有性生殖的状态;其叶片越大,光合作用产物就会越多,可以满足有性生殖的需要。熊治廷对安徽天堂寨自然保护区内野生独花兰进行了两月有余的连续定株观测和群体统计,发现独花兰植株开花与否和地下假鳞茎的数目、地下茎的总体积和叶片的面积具有极显著相关性,绝大多数开花的独花兰地下假鳞茎数为3,地下茎总体积通常达8 cm3,叶片的面积达 33 cm3[22]。个体较大的植株,叶片大,光合产物多,开花持续期较长。此外,独花兰可依靠自身的假鳞茎进行缓慢的无性生殖[23],其假鳞茎具新老更替特点,经过1个生长季后即不再生叶开花,以后由新的假鳞茎继续展叶开花,因老的假鳞茎所储存的营养物供应了抽葶开花的需要,在花谢后老的假鳞茎呈现出萎缩坏死状[22]。陈戈[24]等在春秋两季利用假鳞茎,采取组织个体分株法进行独花兰繁育,独花兰生长2~3年可进行1次分株,分株后至少需要有5个连接的假球茎。
4 组织培养育苗
为解决独花兰自然情况下的繁殖困难,传粉受精与结实率较低的问题,相关学者通过研究,成功提出了独花兰的快速繁育方案。Jiang[25]等分别以4月和6月份采集的假鳞茎和节段化假鳞茎为外植体,1/2MS+1.0 mg/L6-BA+0.05 mg/LNAA+10%椰子汁培养基为芽诱导培养基,结果显示,节段化假鳞茎上没有芽产生,认为这与假鳞茎切面上有大量酚类物质的渗出,导致节段化的假鳞茎坏死有关;4月份采集的假鳞茎为外植体培养12周可产生腋芽,6月份所采的假球茎为外植体,培养7周后,每个外植体产生1.4个芽,这一现象与独花兰4月花期营养物质的消耗有关,同时认为以独花兰假鳞茎为外植体时,适宜采收期为6月上旬。最佳生根诱导培养基为1/2MS+1.0mg/LNAA+0.1 mg/L6-BA,平均每芽生根4.5条,最高生根率为88.9%。将4~5 cm高的试管苗移栽到腐殖质:沙土(1:1)混合物的盆栽中,在温室中生长7周后移栽到大田,之后在野外生长两个月后成活率约为75%。此外,刘国顺[3]、高丽[26]、刘赛思[27]等也研究了独花兰的组织培养技术。刘国顺发现75%酒精消毒30 s,然后0.2%HgCl消毒6~7 min为外植体消毒的最佳方式,试验外植体的污染率为14.3%[3]。结合相关文献,对独花兰不同外植体、培养基、诱导率和生根率进行了汇总(见表1)。目前,组织培养育苗技术是实现独花兰快速规模化繁殖的一种有效手段。

表1 文献中记载的独花兰组织培养情况
5 遗传变异特征
基于BLAST1鉴定、遗传变异、建树分析,巫伟峰[28]等对12个兰属本地样品和250条GenBank下载的兰科ITS2序列(其中81条属于兰属)进行了评估。结果表明,ITS2序列在兰属植物的分子鉴定、亲缘关系、系统发育关系等方面具有一定的应用价值,可作为兰属植物DNA条形码鉴定的辅助条形码。独花兰作为兰科的单种属植物,可以结合ITS2序列对同族植物进行研究DNA条形码和系统发育的关系。为了研究独花兰的分类地位,Yi[29]利用下一代测序技术测定了独花兰的叶绿体全基因组序列,结果表明独花兰与布袋兰亲缘关系较近。Li[30~31]结合传粉和生态学的研究结果,利用用随机扩增多态DNA(RAPD)技术对独花兰的遗传多样性水平和分配进行了研究,结果表明,可能是由于生境破坏致使独花兰数量减少甚至局部灭绝,基因流动受限导致独花兰群体内变异低而群体间变异大;并提出了独花兰栖息地保护和传粉者保护、人工授粉以及迁地保护的保护管理措施。孙海芹[32]等采用(单因素方差分析法)和UPGMA聚类分析等方法,研究了分布于神农架、庐山和新宁的12个自然居群独花兰的形态变异,发现庐山居群13个性状的平均观测值均高于新宁和神农架居群,在物种水平上各性状存在较多的变异,并且营养器官变异性较高。单因素方差分析结果显示3个地区间多个形态性状存在极显著差异;UPGMA聚类分析表明这3个地区分别形成明显不同的分支。在以传粉者为媒介自然选择的影响下,神农架龙门河和关门山两个地点间存在明显的形态分化。
6 展望
目前,关于独花兰的研究报道相对较少,主要集中在分布区域、群落特征、繁育特征、组织培养育苗技术及遗传与变异等方面,尚未见关于独花兰的化学成分、药理作用等方面的报道,这可能与独花兰为濒危植物,资源稀缺有较大关系。独花兰是一种珍贵的濒危药用植物,具有较高的药用价值;且独花兰耐阴又耐寒,植株矮小,花色艳丽,具有较高的观赏价值,是室内培养的良好的花卉品种,这便导致了独花兰采挖过度,资源减少。同时,独花兰对传粉者的高度依赖和专一性,致使传粉困难,结实率低,还有种子的萌发也需要与菌根真菌形成共生关系,在适宜的温度和湿度下才能萌发,生态环境的破坏可能影响其传粉者和菌根真菌,从而影响独花兰的自然生长与繁育,这又进一步导致了独花兰资源的稀缺。针对现状,应该积极采取应对措施保护独花兰生境,利用人工授粉和组织培养育苗技术提高独花兰的繁育率,丰富其资源。在此基础上,对独花兰进行更深入的研究,以期在药用、生态、遗传及观赏等方面体现更高的综合价值。
猜你喜欢
中国兽医学报(2022年5期)2022-11-16
科学之友(2021年12期)2021-12-23
老年博览·上半月(2021年12期)2021-12-21
安徽农业科学(2018年15期)2018-05-14
大科技·百科新说(2018年3期)2018-04-11
科学种养(2017年11期)2017-11-28
成才之路(2016年27期)2016-09-30
湖北农业科学(2014年19期)2014-12-02
农村农业农民·B版(2011年5期)2011-06-16
农业工程技术·温室园艺(2009年9期)2009-05-17
