
大亚湾夏季浮游植物群落结构的长期变化及其与环境因子的关系
2022-08-17 06:51杨熙李开枝谭烨辉吕意华
海洋学报 2022年8期
杨熙 ,李开枝,谭烨辉,吕意华
( 1.国家海洋局南海环境监测中心,广东 广州 510300;2.自然资源部海洋环境探测技术与应用重点实验室,广东 广州510300;3.海南南沙珊瑚礁生态系统国家野外科学观测研究站,广东 广州 510300;4.中国科学院南海海洋研究所,广东广州 510301)
1 引言
随着人类活动的增加和气候变化的影响,海洋环境已发生了深刻的变化,如污染物的大量排放导致水体营养盐浓度上升以致出现富营养化[1],全球变暖导致海水温度上升等[2]。浮游植物是海洋生态系统中最主要的初级生产者,约占地球初级生产量的一半[3],在生态系统的物质循环与能量流动中起着十分重要的作用[4−5],当海洋环境发生变化时其能作出相应的响应。海湾处于陆地和海洋之间的交会地带,受人类活动和气候变化的双重影响。国际上研究历史比较长的几个典型的海湾中,切萨皮克湾和东京湾浮游植物群落结构的长期变化是由人类活动所引起的水体富营养化导致的[6−7],而纳拉甘西特湾浮游植物群落结构的长期变化则受气候变化和人类活动两个方面影响[8]。
大亚湾是南海北部沿岸一个较大的山地溺谷型半封闭海湾,其周边区域以石油化工工业、港口航运业、海水养殖业和旅游业最为发达。由于区域经济的飞速发展和人类活动的加强导致大亚湾已由贫营养状态发展到中营养状态,且湾内部有些区域已出现富营养化的趋势[9]。研究显示,河流污染物的输入是引起湾内部水体出现富营养化的主要原因[10],其中引起富营养化的主要成分是溶解性无机氮(Dissolved Inorganic Nitrogen,DIN),其浓度由 1985 年的21.42 μg/L增加至2004年的75.60 μg/L[11]。大亚湾浮游植物以硅藻为主,其次为甲藻,硅藻的主要优势种有角毛藻属(Chaetocerossp.)、伪菱形藻属(Pseudonitzschiasp.)、翼根管藻纤细变型(Rhizosolenia alataf.gracillima)、中肋骨条藻(Skeletonema costatum)、菱形海线藻(Thalassionema nitzschioides),叉角藻(Ceratium furca)是甲藻中的主要优势种[12]。水体富营养化导致了浮游植物种类数和丰度的下降、优势种向小型化发展、生物多样性降低以及甲藻赤潮的频繁发生[13−14]。此外,气候因素也是影响大亚湾浮游植物群落的重要因素,如厄尔尼诺现象和降雨量的增加等会给海湾带来营养盐的输入,导致浮游植物生物量的增加和赤潮的暴发[10,15]。
已有的大亚湾浮游植物群落结构长期变化相关报道主要是针对1990−2004年间的情况,而2004年至今的10多年里随着粤港澳大湾区建设的开展,大亚湾区域经济高速发展,人口数量激增,人类活动对大亚湾环境的影响愈发强烈,如海水养殖面积增大、核电站温排水、大量工业废水的排放等,这些因素均会对浮游植物群落产生影响,而此期间大亚湾浮游植物群落的长期变化情况和影响因素还少见报道。夏季为一个较为特殊的季节,该时期大亚湾浮游植物生长旺盛,且大亚湾处于丰水期,降雨量充沛,一年中大亚湾营养盐的输入主要集中在该时期,而且还存在由粤东沿岸上升流所引起的外海水入侵现象,本文利用1999−2017年历年夏季大亚湾环境因子和网采浮游植物的调查数据,对大亚湾整体区域进行分析,以期获得夏季大亚湾浮游植物群落结构的长期变化规律及其变化原因。
2 材料和方法
2.1 数据来源
关于大亚湾浮游植物的研究,开始于20世纪80年代初期,徐恭昭[16]对大亚湾整个海域浮游植物的分布做过多次调查。进入90年代后,中国科学院大亚湾海洋生物综合实验站和国家海洋局南海环境监测中心均对大亚湾进行了长期调查,积累了大量的调查数据,其中中国科学院大亚湾海洋生物综合实验站在大亚湾内布设了12个固定的调查站位,国家海洋局南海环境监测中心在大亚湾内布设了18个固定的调查站位,具体调查站位的分布情况见图1。本文所分析的数据为1999−2017年大亚湾夏季的调查数据,其中1999−2003年的调查数据由中国科学院大亚湾海洋生物综合实验站提供,2004−2017年的调查数据由国家海洋局南海环境监测中心提供,调查要素包括水温、盐度、营养盐(NO3-N、NO2-N、NH4-N和PO4-P)浓度和网采浮游植物数据,数据基本信息见表1。
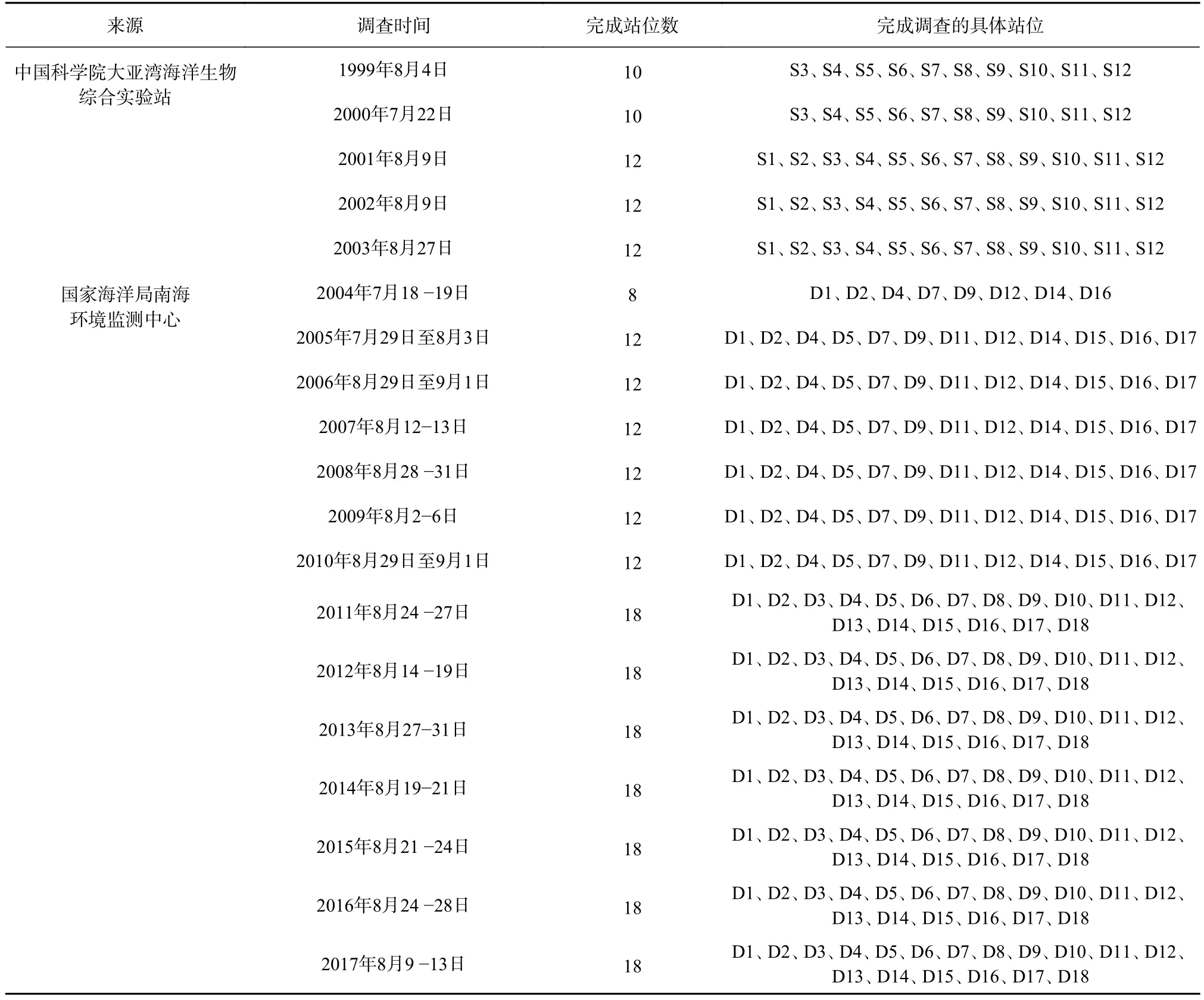
表1 调查数据基本信息Table 1 Basic information of survey data
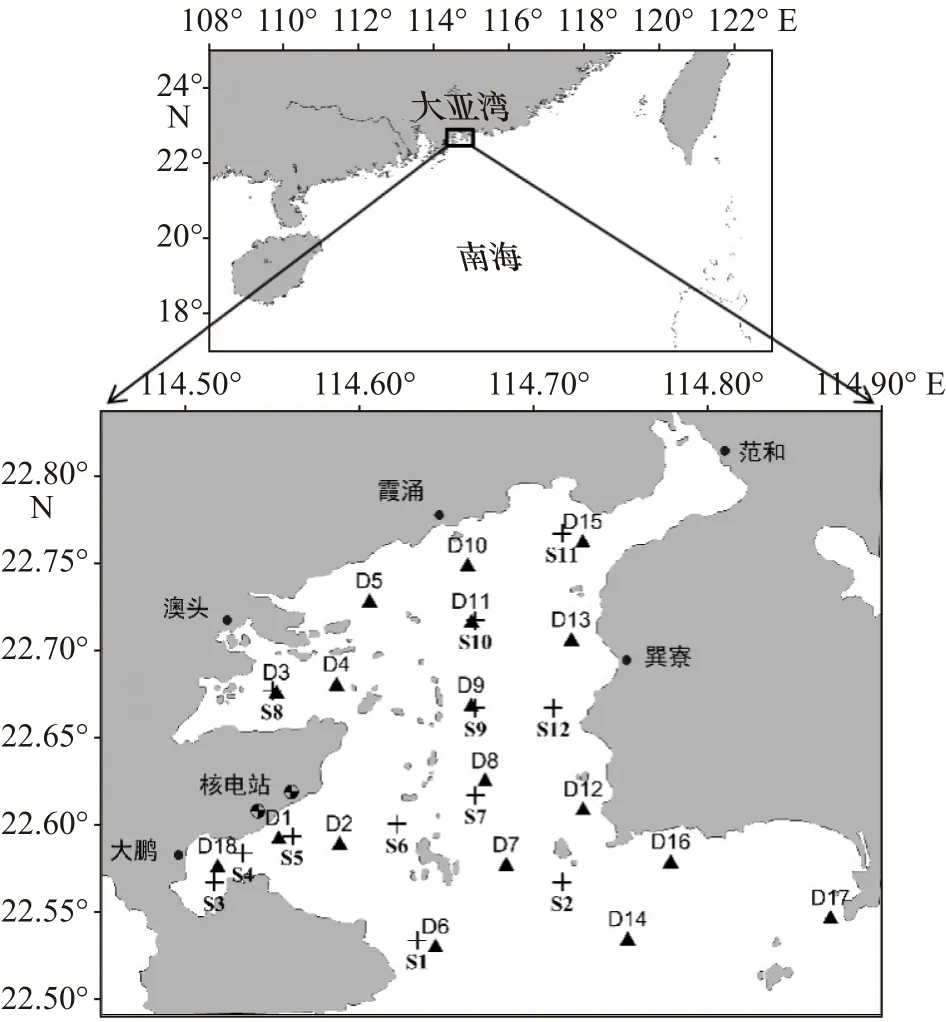
图1 调查区域和站位分布Fig.1 Distribution of study area and monitoring stations
2.2 样品的采集和分析
历年数据中样品的采集和分析方法均一致。其中表底层水温和盐度数据获取采用CTD数据和仪器测量相结合的方法。采集的表底层营养盐水样,经孔径为0.45 μm的醋酸纤维滤膜(使用前经过45~55℃烘烤 6~8 h)过滤,4项营养盐(包括 NO3-N、NO2-N、NH4-N和PO4-P)浓度的测定参考《海洋调查规范》(GB/T 12763.4−2007),文中 DIN 浓度为 NO3-N、NO2-N和NH4-N浓度之和。浮游植物使用标准的浅水III型浮游生物网(网长为140 cm、网口内径为37 cm、网口面积为0.1 m2、筛绢孔径为77 μm)进行采集。采集浮游植物样品时,放网速度应控制在0.5 m/s,下放至离底2 m处时静置1 min左右,然后再以0.1 m/s的速度向上垂直拖网,将所采集的样品装入1 L不透明广口PE瓶中,用甲醛固定后置于阴凉处保存(甲醛体积终浓度为2%)。浮游植物样品鉴定计数之前先利用虹吸法进行浓缩处理。鉴定计数时,先将浓缩样品摇匀,然后用200 μL的移液枪吸取0.1 mL的溶液,均匀分散在0.1 mL的浮游植物计数框中,置于光学显微镜下进行种类鉴定和定量计数,每个样品计3片。
2.3 数据处理方法
浮游植物群落特征参数:优势度(Y)、香农−维纳指数(H′,又称多样性指数)[17]和Pielou均匀度指数(J)[18]的计算公式分别为
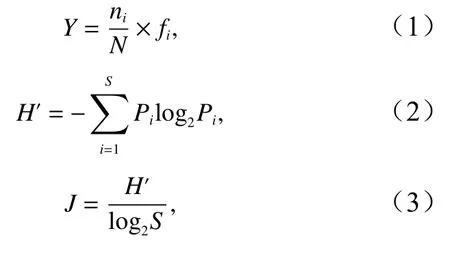
式中,ni为第i种的数量;fi为该种在各站出现的频率;N为群落中所有种的数量;S为浮游植物种类数;Pi=Ni/N,Ni是第i个物种的个体数。Y≥0.02即判定该种为该区域的优势种。
利用Mann-Kendall 非参数检验法来分析大亚湾环境因子和浮游植物群落时间序列的单调变化趋势,其中浮游植物丰度数据经lg(x+1)转化,该方法无需数据符合正态分布,也不受少量异常值和缺失值的影响,因此广泛应用于水文、海洋等数据的趋势分析中。利用 Canoco 5(Microcomputer Power, 美国)软件分析浮游植物与环境变量间的相关性,由于去趋势对应分析(DCA)结果显示4个轴中梯度最大值小于3,因此选择冗余分析(RDA)方法,不同种类浮游植物丰度取各年度所有站位平均值进行分析,所有浮游植物种类丰度数据均经过lg(x+1)加以转化,以降低优势种类对排序的影响。文中环境因子和浮游植物的年际变化点线图、柱状图等采用Origin 8.0软件完成绘制。在进行数据分析时,我们将缺失年份数据忽略,只对存在数据的年份进行统计分析;温度、盐度和营养盐浓度取表底层平均值进行分析。方差分析、t检验等数据统计处理及分析均通过SPSS 13.0统计软件完成,所有统计分析结果中p<0.05时即视为显著。
3 结果
3.1 环境因子的长期变化
1999−2017 年各环境因子的年际变化情况见图2。由图可见,海水温度和盐度年际波动较大,Mann-Kendall检验结果显示,海水温度呈显著下降趋势(p<0.05),海水盐度则与温度相反,呈现显著上升趋势(p<0.05)(表2)。营养盐方面,DIN 和 DIP 浓度的长期变化趋势均不显著(p>0.05),从营养盐的年际变化图上可以看到,大亚湾DIN浓度以NO3-N为主,NH4-N次之,NO2-N浓度最低。DIN浓度的长期变化走势与NO3-N浓度较为一致,在1999−2007年间,溶解无机氮浓度变化幅度较小(26.54~134.94 μg/L),平均值为(52.91±19.69)μg/L,2008−2017 年间,其浓度变化幅度较大(42.84~203.21 μg/L),平均值为(91.39±49.70)μg/L,相比于1999−2007年间的平均值提升了72.73%。溶解无机磷(Dissolved Inorganic Phosphorus,DIP)浓度的年际波动较大,其在2007−2009年间出现较高值(15.84~17.79 μg/L),其他年份 DIP 浓度在 1.86~13.87 μg/L 间波动。DIN∶DIP值的长期变化趋势也不显著(p>0.05),如果以雷德菲尔德化学计量比(16∶1)作对比,可以看到1999−2017年间只有少数年份(2000年、2007年、2009年和2014年)的DIN∶DIP值在16以下,其他年份的DIN∶DIP值均在16以上,其中在2011年和2015年大亚湾DIN∶DIP值出现较大幅度上升,分别达到 58.32±30.08 和 85.46±42.58。
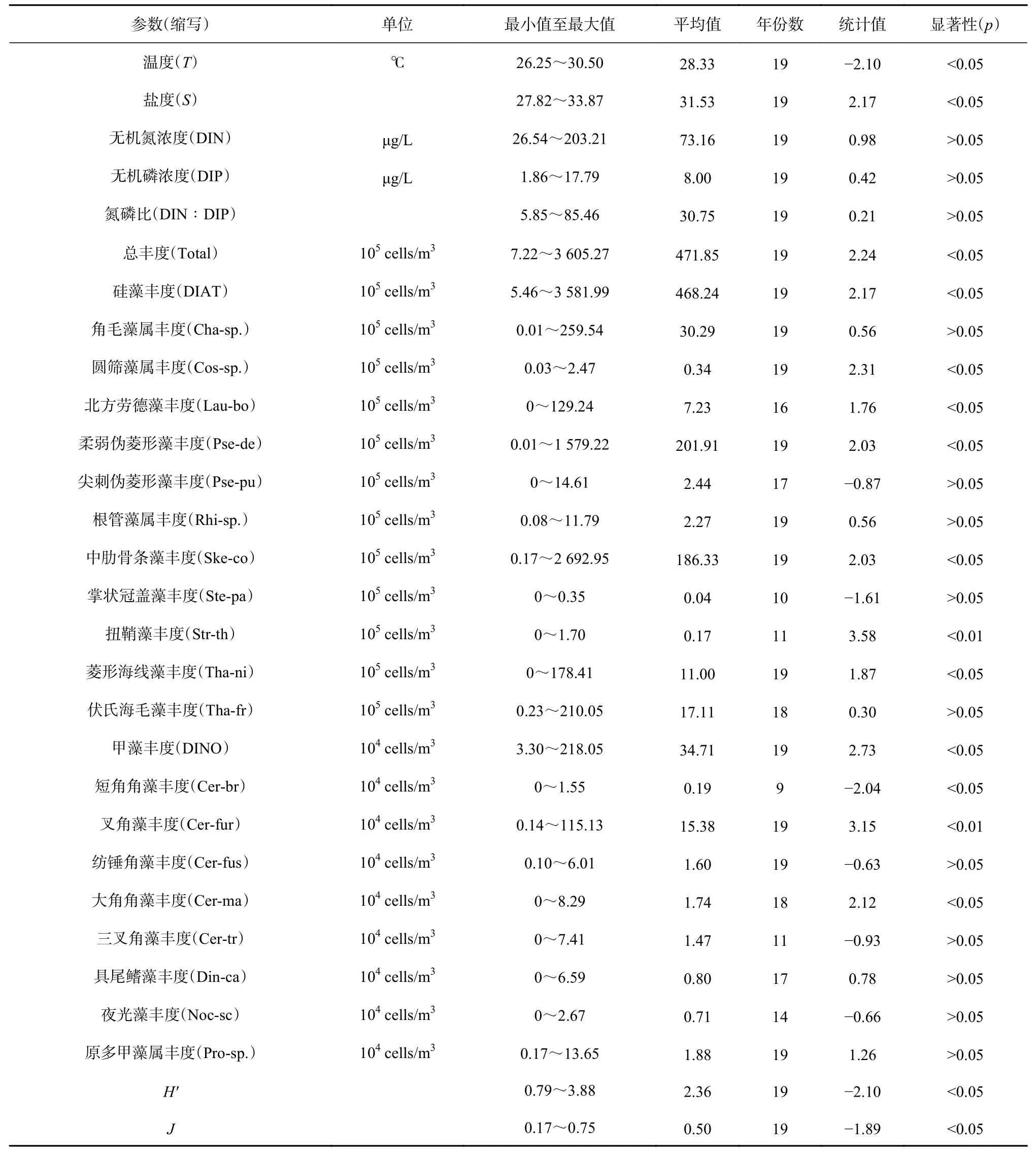
表2 环境因子和浮游植物群落结构长期变化趋势的Mann-Kendall检验结果Table 2 Results of the Mann-Kendall test for detection of long-term changes in the environmental factors and the phytoplankton community structure
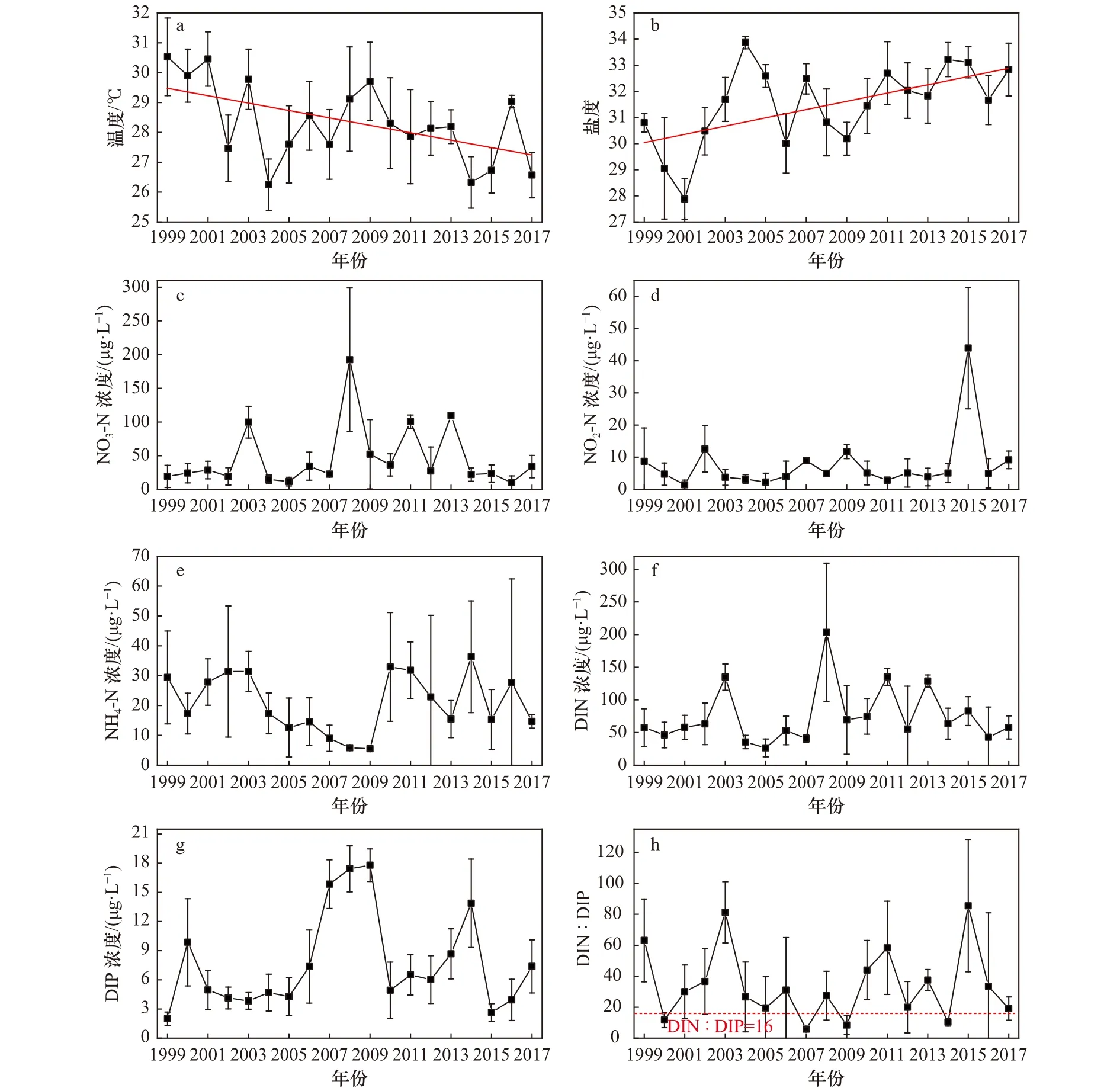
图2 1999−2017年大亚湾夏季环境因子的长期变化Fig.2 Long-term changes of environmental variables in the Daya Bay in summer between 1999 and 2017
3.2 浮游植物群落结构的长期变化
3.2.1 种类组成
1999−2017 年间大亚湾夏季共鉴定出浮游植物6门81属332种,其中种类数最多的为硅藻(55属226种),其次为甲藻(19属91种),其他门类种类数较少。浮游植物种类数的年际变化情况见图3,可以看到,种类数的年际波动较大,其中总种类数的变化范围为49~110种,硅藻种类数的变化范围为37~85种,甲藻种类数的变化范围为8~27种。每年均以硅藻种类数最多,其次为甲藻,其他种类数较少。从长期变化来看,总种类数年际波动较大,变化趋势不明显,硅藻和甲藻种类数的变化趋势也不明显。
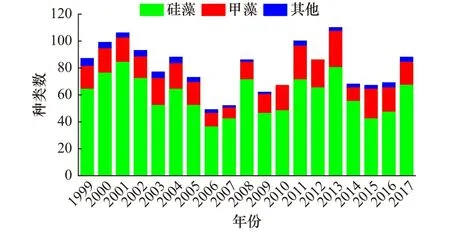
图3 1999−2017年大亚湾夏季浮游植物种类数长期变化Fig.3 Long-term changes of phytoplankton species number in the Daya Bay in summer between 1999 and 2017
3.2.2 优势种
1999−2017 年大亚湾夏季浮游植物优势种的年际变化情况见表3,大亚湾浮游植物优势种种类数较多,均属于硅藻和甲藻两门类,其中硅藻优势种数量最多,以耐高温和盐度适应能力较强的种类占优势,如近岸种柔弱伪菱形藻(Pseudonitzschia delicatissima)、中肋骨条藻和菱形海线藻等为优势种。柔弱伪菱形藻为大亚湾海域第一优势种,出现频率最高,其次为中肋骨条藻和菱形海线藻,出现过的甲藻优势种有叉角藻、纺锤角藻(Ceratium fusus)、三叉角藻(Ceratium trichoceors)、大角角藻(Ceratium macroceros)和锥状斯克里普藻(Scrippsiella trochoidea),其中叉角藻出现频率最高,在2014年其优势度最高,并成为海区第一优势种。优势种存在较大的年际变化,但从长期变化来看,主要优势种类的长期变化不大。
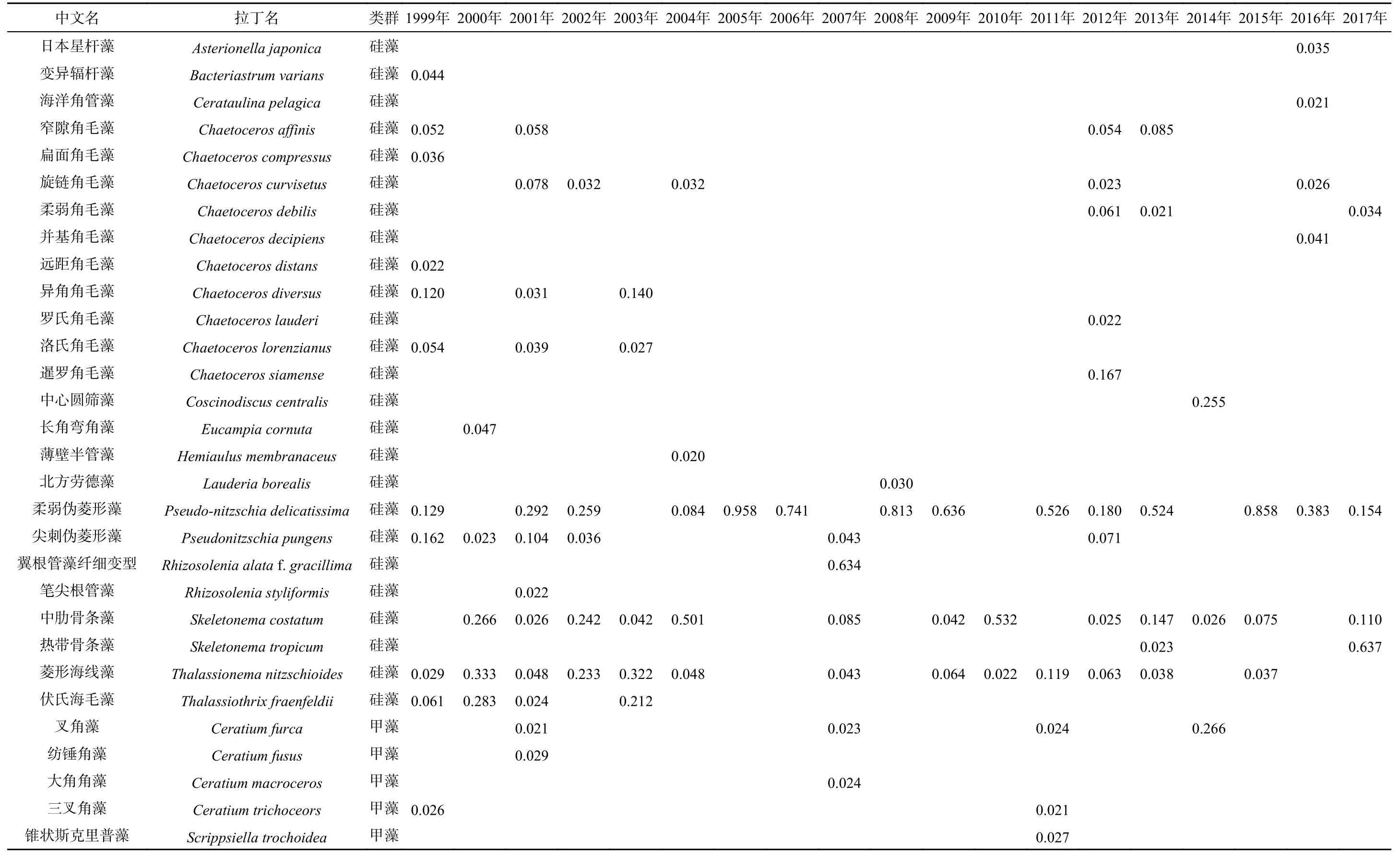
表3 1999−2017年大亚湾夏季浮游植物优势种优势度的长期变化Table 3 Dominance changes of phytoplanktondominant species in the Daya Bay in summer between1999 and2017
3.2.3 浮游植物丰度
Mann-Kendall检验结果显示,浮游植物总丰度、硅藻丰度和甲藻丰度均呈现显著上升趋势(p<0.05)(表2)。浮游植物总丰度和硅藻丰度的年际变化情况非常相似,这是由于大亚湾浮游植物中硅藻占据绝对优势所致,它们在1999−2011年间丰度虽有较大波动,如在2000年、2005年和2008年丰度出现过较大幅度增加,其中在2008年硅藻丰度达到了(1.78±2.22)×108cells/m3,但总体来看变化趋势不明显,丰度处于较低水平。在2011−2017年间总丰度和硅藻丰度呈现明显的上升趋势,硅藻丰度由2011年的(1.44±3.48) ×106cells/m3上 升 至 2017年 的 (3.58±2.39) ×108cells/m3。甲藻丰度在1999−2007年间变化趋势不明显,丰度处于较低水平,在2007−2017年丰度呈现出明显的上升趋势,由2007 年的(4.33±5.19)×104cells/m3上升至2017 年的(2.18±3.84)×106cells/m3(图4a 至图4c)。1999−2017年间浮游植物类群丰度占比年际变化显示,硅藻丰度占比处于69.92%~99.82%之间,平均值为95.20%;甲藻丰度占比一般处于10%以下;其他门类丰度占比均低于1%。由于近年来甲藻丰度的增加,其在某些年份出现了丰度大幅增加的现象,如2014年其丰度占比达30%(图4d)。
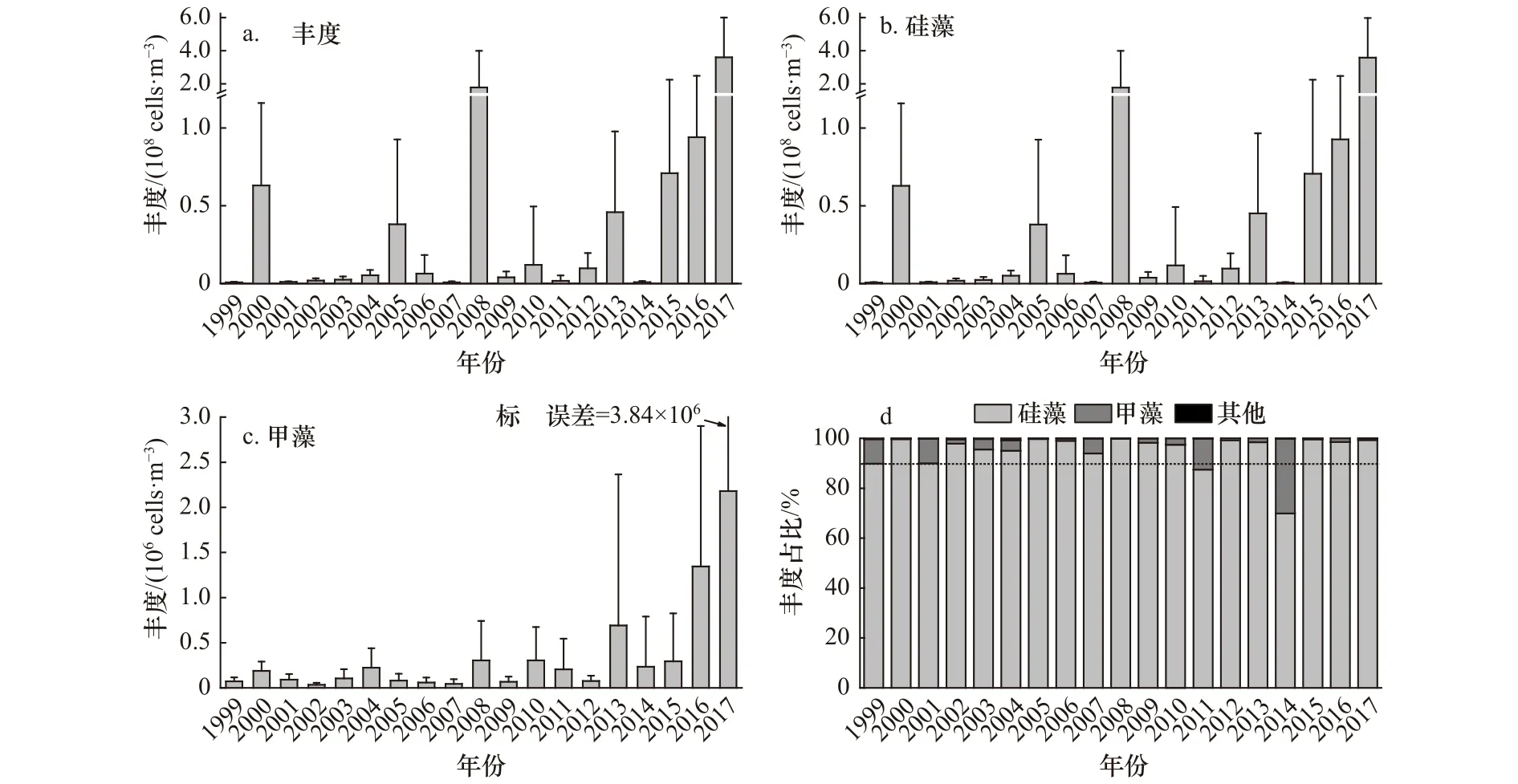
图4 1999−2017年大亚湾夏季浮游植物生物量的长期变化Fig.4 Long-term changes of phytoplankton groups abundance in the Daya Bay in summer between 1999 and 2017
本研究列举了5种大亚湾浮游植物主要优势种类的丰度及其与总丰度占比年际变化情况(图5)。柔弱伪菱形藻丰度呈现显著上升趋势(p<0.05),其丰度占比平均值达37.11%,为大亚湾第一优势种。中肋骨条藻丰度也呈现显著上升趋势(p<0.05),其丰度占比平均值为19.36%,为大亚湾第二优势种,其丰度占比在2011−2017年大体呈现显著升高趋势,而此期间柔弱伪菱形藻丰度占比呈下降趋势,这说明近些年来中肋骨条藻有取代柔弱伪菱形藻成为海区第一优势种的迹象。叉角藻丰度呈现显著上升趋势(p<0.05),其在甲藻中的丰度占比呈现显著升高的趋势(p<0.05),由 1999年的 11.71% 升至 2016年的 85.59%,在甲藻中占据绝对优势。
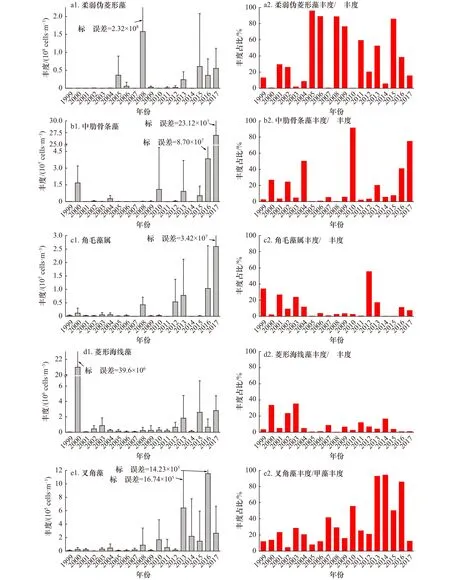
图5 1999−2017年大亚湾夏季几种主要浮游植物种类丰度和占比的长期变化Fig.5 Long-term changes of phytoplankton dominant species abundance and their abundance percentages in the Daya Bay in summer between 1999 and 2017
3.2.4 多样性指数
Mann-Kendall检验结果显示,浮游植物多样性指数(H′)呈现显著下降趋势(p<0.05),均匀度指数(J)也呈下降趋势,但趋势并不显著(p>0.05)(表2)。由年际变化趋势图可以看出,多样性指数(H′)和均匀度指数(J)的变化走势较为相似,其中多样性指数(H′)由1999年的 3.88±0.35降至 2017年的 1.66±0.53,均匀度指数(J)由1999年的0.75±0.05降至2017年的0.35±0.11(图6)。
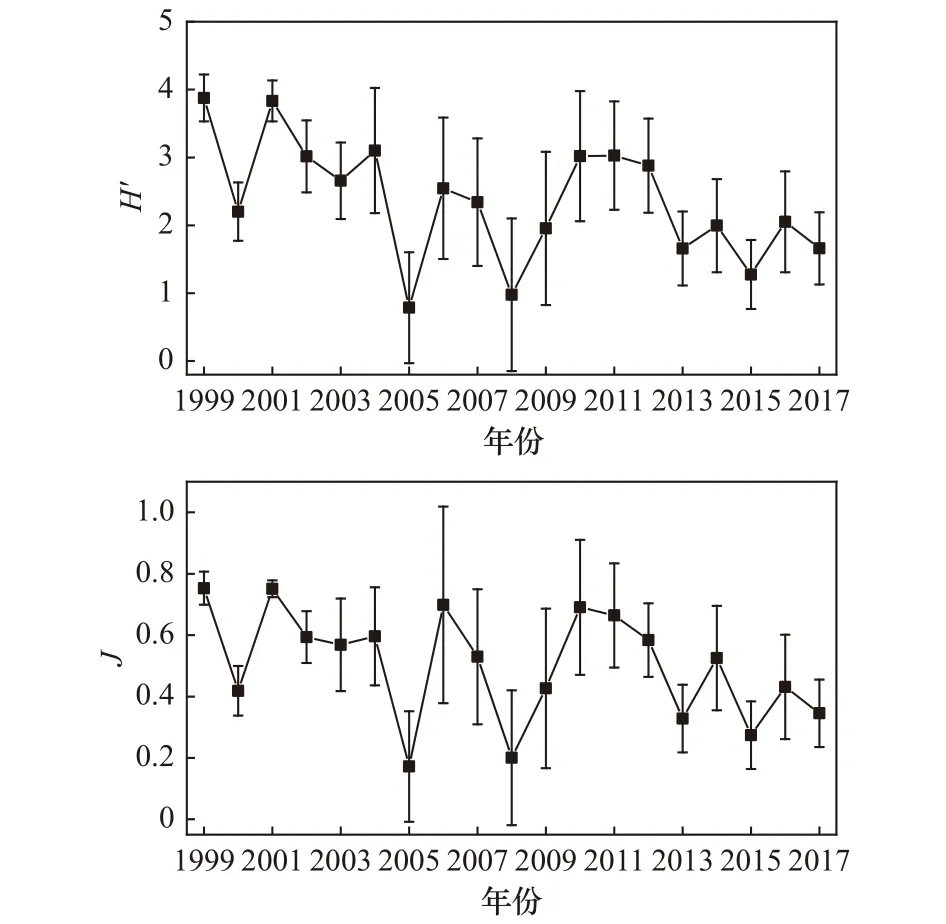
图6 1999−2017年大亚湾夏季浮游植物生物多样性指数(H′)和均匀度指数(J)的长期变化Fig.6 Long-term changes of species diversity indes (H′) and evenness indes (J) in the Daya Bay in summer between 1999 and 2017
3.3 相关性分析
图7a显示,可以将不同年份进行一个较为松散的聚类,较早的年份(1999−2007年)大多数可以聚为一类,其所对应的环境状况为高温低盐、DIN和DIP浓度适中;较近的年份(2008−2017年)大多数可聚为一类,其所对应的环境状况为低温高盐、高DIN浓度和高DIN∶DIP比值,DIP浓度较低的状态。这说明1999−2017年夏季大亚湾海水环境特征出现了由高温低盐向低温高盐、高DIN浓度和高DIN∶DIP比值的转变,这与环境因子的长期变化趋势一致。
图7b显示,硅藻中的柔弱伪菱形藻、中肋骨条藻、菱形海线藻、扭鞘藻(Streptothece thamesis)、北方劳德藻(Lauderia borealis),甲藻中的叉角藻、原多甲藻属(Protoperidiniumsp.)等与盐度、DIN、DIN∶DIP比值呈正相关,与温度和DIP呈负相关。而这些种类中柔弱伪菱形藻、中肋骨条藻、菱形海线藻、叉角藻等为大亚湾主要浮游植物优势种,这也就导致了硅藻和甲藻丰度与环境因子也呈现相似的关系。此外,硅藻中的角毛藻属、尖刺伪菱形藻(Pseudonitzschia punges)、伏氏海毛藻(Thalassiothrix frauenfeldii)、掌状冠盖藻(Stephanopyxis palmeriana),和甲藻中的短角角藻(Ceratium breve)、纺锤角藻、三叉角藻、夜光藻(Noctiluca scintillans)与温度、DIN浓度、DIN∶DIP比值呈正相关,而与盐度和DIP浓度呈负相关。根管藻属、圆筛藻属和大角角藻等与之相反,它们与盐度和DIP浓度呈正相关,与温度、DIN浓度、DIN∶DIP比值呈负相关。而具尾鳍藻(Dinophysis caudata)与DIN浓度、DIN∶DIP比值呈正相关,与DIP浓度呈现负相关,而与温度和盐度的相关性不明显。
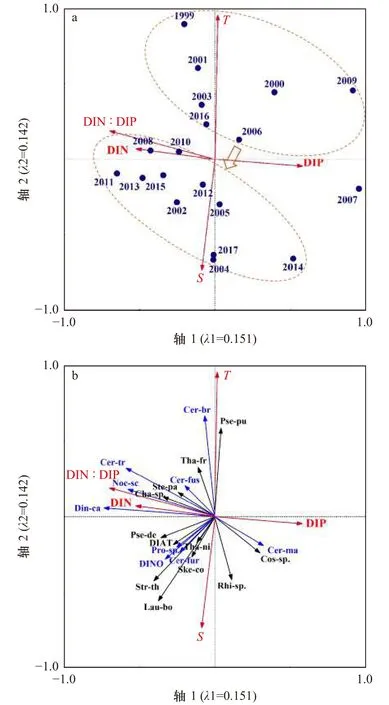
图7 冗余分析图Fig.7 Correlation plots of the redundancy analysis
4 讨论
4.1 环境因子长期变化原因分析
由于大亚湾夏季存在外海水入侵的现象,这股入侵海水源自夏季粤东上升流水系,其主要来自于南海北部陆架区50~100 m深处的海水,具有低温高盐特征,其入侵强度存在明显的年际差异,入侵强度大的时候高盐冷水能够深入大亚湾内区至湾顶部[19−20],近20年来其入侵强度有增加的趋势[21],这导致了夏季大亚湾海水温度呈下降趋势,盐度呈上升趋势。
大亚湾DIN浓度在2007年后出现大幅度提升。引起大亚湾DIN浓度升高的原因主要有以下两个方面:一是地表径流输入量的增加,统计显示1992−2017年大亚湾常住人口由4.38万增加至21.20万,地区工业生产总值由0.20亿美元增加至240.70亿美元,其中地区工业生产总值的增长主要集中在2007年之后[22],人口的急剧增加和经济的快速发展会导致生活污水和工农业废水的产生量大幅增加,这些含有大量DIN的污水通过河流和排污口等注入大亚湾。大亚湾存在10多条小型季节性河流,其中径流量较大的有淡澳河、霞涌河、岩前河和柏岗河等,研究显示,大亚湾夏季DIN的补充绝大部分来源于河流注入,通过河流输送进入大亚湾的氮占氮总输入量的90%以上[23]。降雨量的增加也会导致营养盐输入量的增加,根据距离大亚湾较近的香港天文台降雨量统计数据显示,2008−2017年间7月平均降雨量要明显高于1991−2007年。二是海域点源污染的增加,包括海水养殖污染和船舶污染,其中以海水养殖污染为主,湾内网箱养殖面积已由90年代初的几十公顷发展到目前的将近800 hm2,网箱养殖过程中多余的养殖饲料和养殖生物的排泄物会导致水体营养盐浓度,尤其是DIN浓度的升高[24]。上述因素所引起的营养盐输入导致了大亚湾在2007年后DIN浓度出现大幅度升高。大亚湾PO4-P浓度虽在大多数年份均高于Redfield比率(16∶1),但从 PO4-P的绝对浓度来看,大亚湾整体 PO4-P浓度平均值为(7.69±4.98)μg/L,高于浮游植物对其生长临界值(3.10 μg/L)[25],加上陆源输入中含有大量溶解有机磷[23],因此磷对大亚湾浮游植物生长并不构成限制。
4.2 浮游植物群落结构长期变化原因分析
本研究显示,大亚湾浮游植物种类历年均是以硅藻为主,甲藻次之。总种类数在49~110种之间波动,平均80种,1999−2017年间总体变化趋势不明显,生物多样性指数呈显著下降趋势。此前研究显示,20世纪80年代大亚湾浮游植物种类数在150种以上,90年代大多处于120种以上[11],与20世纪80年代至90年代相比,1999−2017年大亚湾浮游植物种类数出现了较大幅度下降,这与早期变化趋势相一致[12,14]。优势种变化方面,本研究显示,1999−2017年大亚湾主要优势种类无明显变化,硅藻中的柔弱伪菱形藻、中肋骨条藻、菱形海线藻和角毛藻一直为大亚湾夏季的主要优势种类,其中柔弱伪菱形藻为大亚湾第一优势种,甲藻中的主要优势种为叉角藻。Wu等[13]对大亚湾1985−2013年间浮游植物群落的变化进行了分析,发现浮游植物主要硅藻优势种类为角毛藻、伪菱形藻、根管藻(Rhizosoleniasp.)、细柱藻(Leptocylindrussp.)和骨条藻(Skeletonemasp.),甲藻主要优势种为角藻属(Ceratiumsp.),长期变化来看主要优势种类以伪菱形藻、角毛藻和中肋骨条藻为主,该结果与本研究基本一致。
大亚湾浮游植物丰度的长期变化与此前研究存在一些差异。早期一些研究结果显示,20世纪80年代至2000年初大亚湾网采浮游植物总丰度呈现下降趋势,通过与同样表征浮游植物现存量的叶绿素a含量相比较发现,此期间叶绿素a含量呈现上升趋势,对于这种矛盾现象,其解释是由于是网采样品在采样过程中可能漏掉了一些个体较小的浮游植物种类,而大亚湾这一部分小个体浮游植物种类的丰度在增加,这表明大亚湾浮游植物群落组成有小型化趋向[9,12−13,26]。本研究显示,1999年以来大亚湾网采浮游植物总丰度、硅藻丰度、甲藻丰度以及一些主要优势种类丰度均呈现上升趋势,且上升趋势在2008年之后最为显著。本研究中浮游植物主要优势种类变化不明显,且丰度均呈现上升趋势,没有出现此前研究中的浮游植物小型化现象,是否出现小型化现象还需要不同粒径浮游植物长期调查数据加以佐证。
本研究相关性分析显示,大亚湾浮游植物群落结构在1999−2007年间主要受水体高温低盐的影响,而2008−2017年大亚湾环境发生了转变,期间主要受水体低温高盐、高DIN浓度和高DIN∶DIP的影响。人类活动导致的营养盐输入引起了大亚湾DIN浓度的升高,这使得一些耐污种类生长占据优势,如大亚湾主要种类中的伪菱形藻属和中肋骨条藻,它们均为耐污种类,在高DIN和高DIN∶DIP的条件下它们种间竞争优势要明显高于角毛藻属和菱形海线藻等其他种类[27],而且它们属于广温广盐种类,能适应由外海水入侵所引起的水体温盐的转变,因此其丰度和丰度占比均呈现显著的上升趋势。叉角藻具有较低的营养盐吸收半饱和常数(Ks)以及特殊的营养盐吸收特征,如“奢侈消费”[28],此外,它属于广温广盐种,能适应温盐的变化[29],这些特征使得它相对于其他甲藻具有更强的竞争优势,导致其丰度在1999−2017年间呈现显著上升趋势,并成为大亚湾甲藻中的绝对优势种。优势种的集中化、单一化也导致了大亚湾浮游植物生物多样性的下降。大亚湾浮游植物群落结构的长期变化情况与胶州湾较为相似,它们均属于半封闭型海湾,且都受到较大的人为活动影响。研究显示,胶州湾在2000年后其水体中NO3-N、NH4-N浓度分别比1962年提高了14.5倍和9.5倍,20世纪50年代至2000年初,其浮游植物总丰度呈现增加的趋势,一些喜肥的种类如中肋骨条藻丰度呈现显著增加趋势而成为最主要的优势种,浮游植物多样性指数呈下降趋势[30−31]。
5 结论
(1)近20年来大亚湾夏季浮游植物主要优势种类没有发生明显变化,优势种以硅藻种类为主,主要为柔弱伪菱形藻、中肋骨条藻、菱形海线藻和角毛藻,叉角藻为甲藻中的第一优势种。
(2)浮游植物总丰度、硅藻丰度、甲藻丰度以及主要优势种类中的柔弱伪菱形藻、中肋骨条藻和叉角藻的丰度均呈显著上升趋势。
(3)柔弱伪菱形藻和中肋骨条藻已成为大亚湾夏季的绝对优势种,叉角藻也发展成为甲藻中的绝对优势种,优势种的单一化导致了大亚湾浮游植物生物多样性指数的下降。
(4)2007年以来人类活动所引起的DIN浓度大幅升高以及外海水入侵加强所引起的海水温度降低和盐度上升是导致大亚湾夏季浮游植物群落结构变化的主要原因。
致谢:向国家海洋局南海环境监测中心和南海海洋研究所所有参与过相关航次样品采集和分析的工作人员致以衷心的感谢!
猜你喜欢
法医学杂志(2022年1期)2022-06-21
法医学杂志(2022年1期)2022-06-21
法医学杂志(2022年1期)2022-06-21
小天使·三年级语数英综合(2022年3期)2022-04-17
成都信息工程大学学报(2021年3期)2021-11-22
天津诗人(2018年3期)2018-11-13
农业与技术(2017年16期)2017-09-16
科技创新导报(2016年24期)2017-03-13
意林(2008年12期)2008-05-14
意林(2008年14期)2008-05-14
