
内耳毛细胞静纤毛高度调控分子机制
2022-08-18 03:00李娜娜徐志刚
自然杂志 2022年3期
李娜娜,徐志刚
山东大学 生命科学学院,山东 青岛 266237
毛细胞(hair cell)是动物内耳中负责感知听觉和平衡信息的感受器细胞,将负载声音信息或平衡信息的机械能信号转换为电信号,这个过程被称为机械-电转换(mechano-electrical transduction,MET)。每个毛细胞的上表面有大量指状细胞突起,统称为纤毛束(hair bundle),由一根以微管蛋白为骨架的动纤毛(kinocilium)和多根以肌动蛋白为骨架的静纤毛(stereocilia)组成(图1(a))[1]。动纤毛在纤毛束的发育过程中发挥着重要作用,但对于机械-电转换不是必需的,甚至在哺乳动物听觉毛细胞发育后期退化掉[2-3]。静纤毛组织成多排高度不同的阶梯状结构,对于机械-电转换至关重要,机械-电转换离子通道就位于较低几排静纤毛的顶部(图1(b)和(c))[4-6]。
在静纤毛之间以及静纤毛与动纤毛之间有大量的细胞外连接把它们联系起来[7-8]。在这些细胞外连接中,顶连接(tip link)联系了较低排静纤毛的顶端与其相邻较高静纤毛的侧部(图1(a)和(b))[7]。顶连接由两个单次跨膜的钙黏蛋白构成,位于上端的是CDH23蛋白,位于下端的是PCDH15蛋白,二者的胞外部分相互结合,共同构成了顶连接(图1(c))[9-10]。在机械能信号的作用下,当静纤毛向较高一排的方向偏转时,顶连接的张力增加,开启位于顶连接下端附近的机械-电转换离子通道,阳离子内流,毛细胞去极化,最终产生感受器电位,完成机械-电转换[11]。已经发现多个蛋白定位于较低排静纤毛顶部、顶连接下方,形成所谓的“MET复合物”,参与执行机械-电转换[12-13](图1(c))。目前已知的MET复合物成分包括TMC1/2、LHFPL5、TMIE以及CIB2,它们与顶连接下端蛋白PCDH15的跨膜区或胞内段结合[10,14-22]。

图1 毛细胞纤毛束示意图:(a)侧面观;(b)顶面观;(c)不同蛋白复合物在静纤毛中的相对位置
静纤毛的阶梯状结构对于毛细胞执行其机械-电转换功能非常重要。静纤毛的高度受到严格调控,部分调控这一过程的蛋白已经陆续被发现[23-26]。本文将对静纤毛高度调控的分子机制进行简要介绍。
1 静纤毛发育分为不同阶段
静纤毛的核心部分由纤维状肌动蛋白(F-actin)束组成,这些纤维状肌动蛋白的正末端(plus end)位于静纤毛的顶部,负末端(minus end)位于静纤毛的根部[1-4]。在发育过程中,毛细胞通过肌动蛋白在正端的聚合和负端的解聚调控静纤毛的高度。静纤毛发育过程最初是在鸡耳蜗中利用扫描电镜得到了详细的观察[27-30]。根据形态上的变化,静纤毛发育可分为四个阶段。第一阶段(胚胎期第8天左右),所有的毛细胞和支持细胞上表面都覆盖着多根短的以肌动蛋白为骨架的微绒毛以及一根以微管蛋白为骨架的纤毛。第二阶段(胚胎期8~12天),位于毛细胞上表面中心位置的动纤毛在胚胎期第9天随机移动到周边,紧接着在胚胎期9~11天所有毛细胞的动纤毛重新定位到毛细胞的同一侧。毛细胞上表面的微绒毛在这一时期发育成为静纤毛并覆盖整个上表面,随后在胚胎期10~12天靠近动纤毛的静纤毛渐次增加其高度,形成静纤毛的阶梯状模式。第三阶段(胚胎期13~16天),通过增加肌动蛋白丝的数量,静纤毛直径显著增加。静纤毛高度保持不变,但其负端的肌动蛋白丝向下延伸进入胞体形成小根(rootlet)。因远离动纤毛而没有在上一阶段延长并形成阶梯状结构的静纤毛会被重吸收,而动纤毛也在这一阶段退化不见。第四阶段(胚胎期17天至孵化),静纤毛再次延长并达到最终的高度。
哺乳动物毛细胞静纤毛发育大体上遵循类似的规律[31-32]。对于小鼠内毛细胞而言,静纤毛初始延长(第二阶段)发生于胚胎期18.5 天之前,这一阶段顶回毛细胞最高一排(Row 1)静纤毛的长度可达到3~4 μm。胚胎期18.5天到出生后4.5天对应于静纤毛发育的第三阶段,在这一时期静纤毛停止延长,直径增加。之后进入第四阶段,最高一排静纤毛继续延长,第二排(Row 2)静纤毛反而变短,直到在出生后19.5天达到最终的高度,分别为6 μm和2 μm左右。
2 成熟静纤毛仅在顶部发生肌动蛋白的更新
肌动蛋白丝在其两个末端可以同时发生聚合和解聚,在正末端一般聚合速度大于解聚速度,负末端解聚速度大于聚合速度。当正末端的净聚合等于负末端的净解聚时,肌动蛋白丝的长度保持稳定,这样一种模式被称为“踏车(treadmill)模型”[33]。研究人员利用基因枪技术在毛细胞中过表达肌动蛋白-GFP,发现成熟静纤毛核心肌动蛋白丝的更新也符合踏车模型,单根静纤毛的纤维状肌动蛋白骨架每2~3天就全部更新一次[34-35]。
随后多个实验室利用多同位素质谱成像、活细胞成像、转基因小鼠等工具发现成熟静纤毛的肌动蛋白骨架相当稳定,肌动蛋白的更新仅在静纤毛顶部发生[36-38]。这些实验结果表明静纤毛是一个相当稳定的亚细胞结构。最近又有报道指出,较低几排静纤毛顶部肌动蛋白的更新较最高一排静纤毛更快,可能与在较低排静纤毛顶部附近发生的机械-电转换有关[39]。
3 Row 1和Row 2蛋白复合物调控静纤毛高度
研究显示特定蛋白在静纤毛高度调控过程中发挥了重要作用。在胚胎期毛细胞中,MYO15A-S和EPS8定位于所有静纤毛的顶部[40]。在出生后及成年毛细胞中,MYO15A-S和EPS8主要定位在最高一排静纤毛顶部,并将GPSM2/GNAI/WHRN三元复合体运输到同样位置,共同形成所谓的“Row 1蛋白复合物”[32,40](图1(c))。在小鼠中敲除不同Row 1蛋白编码基因会引起相似的表型,主要表现为静纤毛排数增多,高度变短且不同排之间高度差异减小(图2(a)和(b))[41-45]。
在较低两排(亦即有机械-电转换功能)的静纤毛顶端定位有另外一个蛋白复合物,称作Row 2蛋白复合物,成员包括MYO15A-L、EPS8L2、TWF2、CAPZB2及BAIAP2L2[46-52](图1(c))。在这一复合物中,EPS8L2、TWF2和CAPZB2有加帽蛋白(capping protein)的活性,被认为发挥了抑制肌动蛋白聚合,从而限制静纤毛高度的作用[46-48,50]。在小鼠中敲除Row 2蛋白编码基因会引起较低两排静纤毛的退化,对最高排静纤毛影响较小(图2(a)和(c))[48-49,51-52]。此外,有研究显示ESPNL也在较低排静纤毛顶部高表达,其编码基因敲除后引起最低一排静纤毛退化[53],而且体外实验表明ESPNL能够结合BAIAP2L2[51],提示ESPNL可能也是Row 2蛋白复合物的成员。
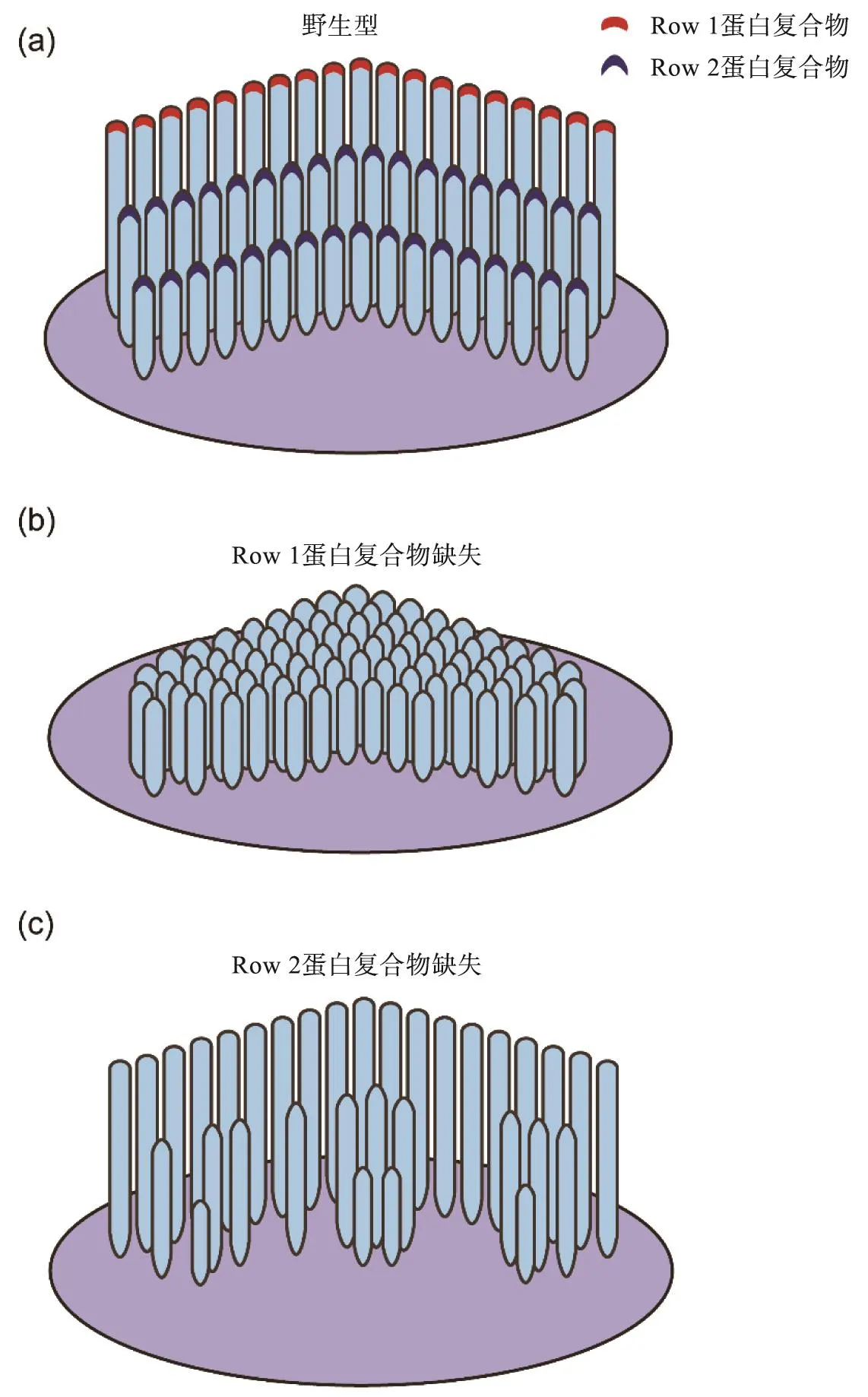
图2 Row 1和Row 2蛋白复合物缺失时对静纤毛发育及维持的影响
4 机械-电转换影响静纤毛高度
前文已经讲过MET蛋白复合物定位于较低几排静纤毛的顶部,与Row 2蛋白复合物的位置接近(图1(c))。与此相应,研究显示机械-电转换会影响静纤毛的高度[32,39,54]。用阻断MET通道或破坏顶连接的药物处理体外培养的听觉感觉上皮,会导致毛细胞较低两排静纤毛高度减小;去除药物后这一表型得到恢复[54]。用胞内钙离子螯合剂处理得到类似的结果,提示这一过程可能与钙离子通过MET通道的内流有关[54]。
基因敲除小鼠为MET蛋白调控静纤毛高度提供了进一步的证据。Tmc1/Tmc2双敲和Tmie敲除小鼠静纤毛的表型类似,主要表现为不同排静纤毛高度及直径差异减小[32]。但二者之间也有一些细微的差异,如Tmc1/Tmc2双敲小鼠毛细胞最高一排静纤毛的高度不变,但Tmie敲除小鼠毛细胞最高一排静纤毛高度减小[32]。体外实验显示成熟毛细胞中肌动蛋白在不同排静纤毛顶部聚合速度不同:在最高一排聚合较慢,在较低两排聚合较快[39]。然而在药物阻断MET或Tmie敲除小鼠毛细胞中,不同排静纤毛顶部聚合速度趋向一致[39]。
LHFPL5和CIB2对静纤毛高度的影响还没有被详细报道过,但根据已发表的扫描电镜结果来看,Lhfpl5敲除小鼠和Cib2敲除小鼠呈现出较低排静纤毛退化的趋势[14,16,19],与Tmc1/Tmc2双敲及Tmie敲除小鼠静纤毛的表型并不一致。这一结果提示我们,除了影响钙离子内流外,不同的MET蛋白可能通过不同的方式影响静纤毛高度。与这一设想一致,最近我们发现MET复合物蛋白CIB2可以结合Row 2复合物蛋白BAIAP2L2,而且BAIAP2L2在Cib2敲除小鼠中不再定位于较低两排静纤毛顶端,表明CIB2可能通过BAIAP2L2直接调控较低排静纤毛的高度[52]。这一模式能否扩展到其他MET蛋白和Row 2蛋白,还有待进一步的研究。
5 展望
成熟毛细胞的静纤毛组织成多排高度不同的阶梯状结构,这对于机械-电转换至关重要,静纤毛形态、结构的异常往往会引起耳聋和前庭功能受损[23]。目前我们对静纤毛高度调控的分子机制已经有了一定的认识,如:成熟毛细胞的静纤毛相当稳定,其核心肌动蛋白丝的更新往往只在顶部发生;特定蛋白定位于静纤毛的顶部,调控不同排静纤毛的高度;机械-电转换可以影响静纤毛的高度;等等。然而,Row 1、Row 2蛋白复合物以及机械-电转换调控静纤毛高度的具体机制目前还不是十分清楚,在这一过程中可能还有其他发挥重要作用的蛋白有待发现。
本文主要关注的是毛细胞静纤毛高度调控的分子机制。除了高度调控之外,关于静纤毛发育和维持还有很多重要问题亟待回答。例如,静纤毛直径调控的分子机制是什么?毛细胞中静纤毛的排数是如何控制的?此外,毛细胞平面细胞极性(planar cell polarity, PCP)的确立对听觉和平衡功能至关重要,在亚细胞水平体现为静纤毛和动纤毛的不对称分布。这一领域已经取得了很多重要的进展,发现了多个调控PCP的重要蛋白,但仍有很多细节和关键问题有待深入探讨[55-57]。
猜你喜欢
生物学杂志(2022年5期)2022-10-20
医学研究生学报(2021年4期)2021-12-02
辐射研究与辐射工艺学报(2021年2期)2021-05-06
临床与实验病理学杂志(2021年3期)2021-04-25
听力学及言语疾病杂志(2020年4期)2020-12-20
中国医科大学学报(2020年8期)2020-08-11
复旦学报(医学版)(2020年2期)2020-04-17
中国畜牧杂志(2020年1期)2020-01-16
中华耳科学杂志(2020年6期)2020-01-08
中华皮肤科杂志(2019年5期)2019-06-24
