
北部湾防城港—钦州近岸海域鱼类群落格局及其与环境因子的关系
2022-08-23 07:36张公俊杨长平周文礼单斌斌王良明王思涵孙典荣贾春斌
南方水产科学 2022年4期
张公俊,杨长平,刘 岩,周文礼,单斌斌,王良明,王思涵,孙典荣,贾春斌
1. 中国水产科学研究院南海水产研究所,广东 广州 510300
2. 天津农学院,天津 330384
3. 中国水产科学研究院南海水产研究所热带水产研究开发中心,海南 三亚 572018
4. 深圳市渔业发展研究中心,广东 深圳 518067
海湾是鱼类的天然繁育场,其特殊的地理位置不仅可提高幼鱼的成活率[1],还能为鱼类提供丰富的食物保障[2]。鱼类根据自身需求的不同,多样化地利用海湾的特殊生境,有些鱼类终生生活于湾内,有些在季节迁移期间或幼鱼期间于湾内觅食活动,另有些仅把海湾作为其在较深海域与河口间的临时过渡区[3]。Blaber等[4]推断至少存在11种鱼类利用卡奔塔利亚湾度过幼鱼期继而游向深海。鱼类作为渔业资源的重要组成部分,其群落分布特征与水域生态环境密切相关,对鱼类群落与环境因子间关系进行研究,一方面可了解鱼类的生活习性,另一方面可探究人类活动对鱼类群落的影响[5]。例如,瑞典西海岸鱼类为躲避低温游向深海[2];巴西北部海域鱼类在降水较多的季节游离近岸以寻求最适的生存盐度[6];Franco和 Santos[7]发现常栖于礁石底质的鱼类更易受盐度影响,而洄游性鱼类分布受温度和盐度的共同作用;欧盟水框架指标(European Water Framework Directive) 通过比较不同 (干扰与未干扰) 水域内的鱼类群落特征,评估了人类活动对水域的干扰程度[8]。综合以往研究,温度、深度、盐度、溶解氧、底质类型、海域地理特征和人为因素是对鱼群分布产生明显作用的环境因子[9-12]。
北部湾位于南海西北部 (105°50'E—110°10'E,17°00'N—21°45'N),东邻广东省的雷州半岛和海南省,西接越南,北靠广西壮族自治区,是一个半封闭海湾[13]。该海湾沿岸河流众多,有中国的南流江、防城河、大风江、北仑河、九州江和越南的先安河、红河、马江等,数量达200多条,其中湾北部有120多条,年径流量为3.0×1011m3,仅4—10月 (雨季)占年径流量的80%以上[14]。北部湾作为中越两国共有的优质渔场,水质肥沃、鱼类资源丰富[13]。目前对北部湾海域的研究集中在鱼类生态位测定[15]、分类多样性分析[16]、单物种时空分布[17]、经济鱼类资源变动[18]和鱼类群落组成研究[19]等方面,然而针对其鱼类群落与环境因子间关系的研究鲜有报道,仅王雪辉等[12]、何思璇和何斌源[20]对北部湾鱼类群落和环境因子间的相互关系开展了一定的研究,结果表明鱼类群落时空异质性较明显,鱼类呈小型、低值化特征,深度、温度和盐度对鱼群分布作用较明显。同时,调查区域多分布在北部湾中部[12]、湾口[16]、西北部[19]和东北部海域[21],鲜见针对北部湾防城港—钦州近岸海域鱼类群落结构年际动态变化的研究。北部湾防城港—钦州近海由于防城江和钦江的注入,其径流携带了大量营养盐和有机碎屑,成为北部湾多种鱼类主要的索饵、产卵和育幼场,对北部湾鱼类资源的补充起到关键作用,是北部湾渔场的重要组成部分[19]。本文以北部湾防城港—钦州近岸海域为研究区域,依据2016年4个航次鱼类资源调查数据,对鱼类资源现状开展科学评估,阐明鱼类群落与环境因子间的关系,以期为北部湾防城—钦州近岸海域渔业资源开发利用和保护提供基础理论支持。
1 材料与方法
1.1 研究区域
2016年2、5、8和11月在北部湾防城港—钦州近岸海域 (108°7.5'E—108°42'E, 21°02'N—21°32'N) 设置29个渔业调查站位,进行4个航次的底拖网调查 (图1)。此海域底部坡度平缓,底质类型主要以粉砂为主[22]。经琼州海峡带来的混合水是产生湾内环流的主要因素,混合水进入湾内使湾北部出现气旋式环流 (春夏) 及东北—西南流向海流 (秋冬)[23]。此海域冬季存在风力较强的东北风,夏季盛行西南风,但西南风相较冬季季风风力弱、不稳定[24]。防城江和钦江为注入此调查区域的两条主要河流,月平均径流量均从4月开始增加,7—8月到达峰值,之后持续下降,至10月趋于稳定,11月至翌年3月维持在较低水平。依据月平均径流量变化特征,将4—10月划分为丰水期,11月至翌年3月为枯水期[25]。
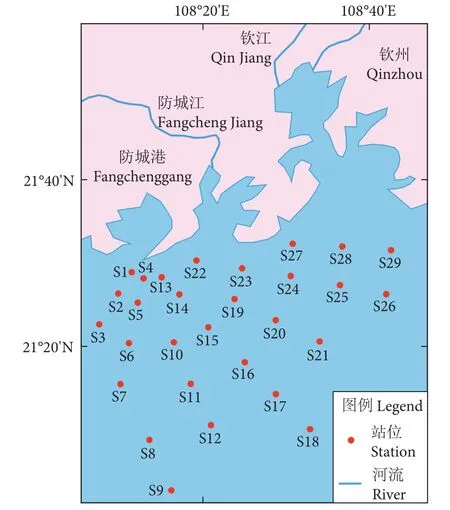
图1 北部湾防城港—钦州近岸海域鱼类资源调查站位图Fig. 1 Sampling station of fishery resources in Fangchenggang(Qinzhou coastal area of Beibu Gulf)
1.2 采样方式与环境数据
本研究对北部湾防城港—钦州近岸海域进行4 次底拖网 (网口宽度 10 m,网口高度 5 m,网囊网目 40 mm) 渔业资源调查,每次拖网时长 0.5 h,平均拖速3节,均在白天作业。调查船为:“桂北渔12075”,船长23.08 m,型宽5.4 m,型深2.7 m,总吨位 90 t,主机功率 162 kW。为更全面地了解环境因子对鱼类分布的影响,选取涵盖该海域各位置的29个调查站位,确保每航次均对大致相同位置的站点实施采样。
在每次拖网结束后,采用YSI 5908 MEMBRANE KIT-12.5 MILPE 对各站点的盐度 (±0.1)、温度 (±0.1 ℃)、溶解氧 (±0.1)、pH (±0.1) 进行测量,深度由船体自带的深度仪勘测。除深度外,以上环境因子数据由每个调查站位采集的表层水样(深度 0.5 m) 及底层水样 (高于底部 0.5 m) 测量得出。每次拖网结束后,对渔获物鉴定到种,记录每个鱼种的数量和体质量,体质量精确到1 g。
1.3 群落分析
1.3.1 丰度和生物量
为使各时空范围内的渔获物信息易于相互比较,评估丰度 (尾·km−2)、生物量 (kg·km−2),计算方法为Y=d/(a×q),d、a、q依次为鱼类渔获率(kg·h−1或尾·h−1)、每小时拖网扫海面积 (km2·h−1)、网具的捕获率 (本研究取0.5)[26]。
1.3.2 鱼类优势种计算
采用相对重要性指数 (Importance relative index, IRI) 计算各季节、各组群鱼类优势种,计算方法为 IRI =(RN+RW)RFO,RN(%)、RW(%)、RFO(%)分别代表某种鱼类丰度、生物量在总丰度或总生物量中的占比、某种鱼类在总站位中出现的频率。采用传统划分标准定义IRI指数,IRI<10为少见种;10≤IRI<100 为常见种;100≤IRI<1 000 为重要种;IRI≥1 000 为优势种[27]。
1.3.3 聚类与排序
利用各站位鱼种重量信息,计算各季节群落多样性指数[18]。根据各站位鱼种数量信息 (去掉机会种和个别干扰站位),计算各站位间的相似度,形成Bray-Curtis相似矩阵并绘制聚类图,采用NMDS (Nonmetric multidimensional scaling) 将聚类站位绘制成图展示群落划分。以上群落多样性指数计算、聚类分析采用软件PRIMER 5.0[28]。为减少优势种的绝对优势度,免除机会种干扰,本研究对多于5尾的所有鱼种数量信息 (共105种,占总鱼种的69%) 及环境因子数据均进行log转换,使结果更准确。利用CANOCO软件对各站位转换后的鱼类及环境因子数据进行排序分析,得出由1、2轴解释的关系图[29]。
调查站位图、鱼类生物量季节变化、平面分布图、聚类图等分别采用 ArcGIS 10.4、Origin 9.1和R软件进行绘制。鱼类及环境因子数据季节差异性利用SPSS 19.0软件统计分析,P<0.05表示差异显著,P<0.01表示差异极显著。
2 结果
2.1 北部湾防城港—钦州近岸鱼类资源状况
北部湾防城港—钦州近岸海域4个航次共捕获鱼类152种,隶属于15目55科96属,其中软骨鱼类2目3科3属3种,占总捕获种类的1.97%,硬骨鱼类13目52科93属149种,占98.03%,如龙头鱼 (Harpadonnehereus)、竹䇲鱼 (Trachurus ja-ponicas) 和卵鳎 (Soleaovata) 等 (表1,附录 A,详见http://dx.doi.org/10.12131/20210255的资源附件)。其中以鲈形目占绝对优势地位,隶属于26科52属,共91种,占总捕获种类的59.87%,如皮氏叫姑鱼 (Johnius belengeri)、多鳞(Sillagosihama) 和褐蓝子鱼 (Siganus fuscescens) 等。春季共捕获鱼类12目93种,夏季13目88种,秋季11目86种,冬季9目76种。四季共有鱼类有32种,包括鲈形目22种、鲽形目4种、鲀形目和鲉形目各2种、灯笼鱼目和鲱形目各1种。
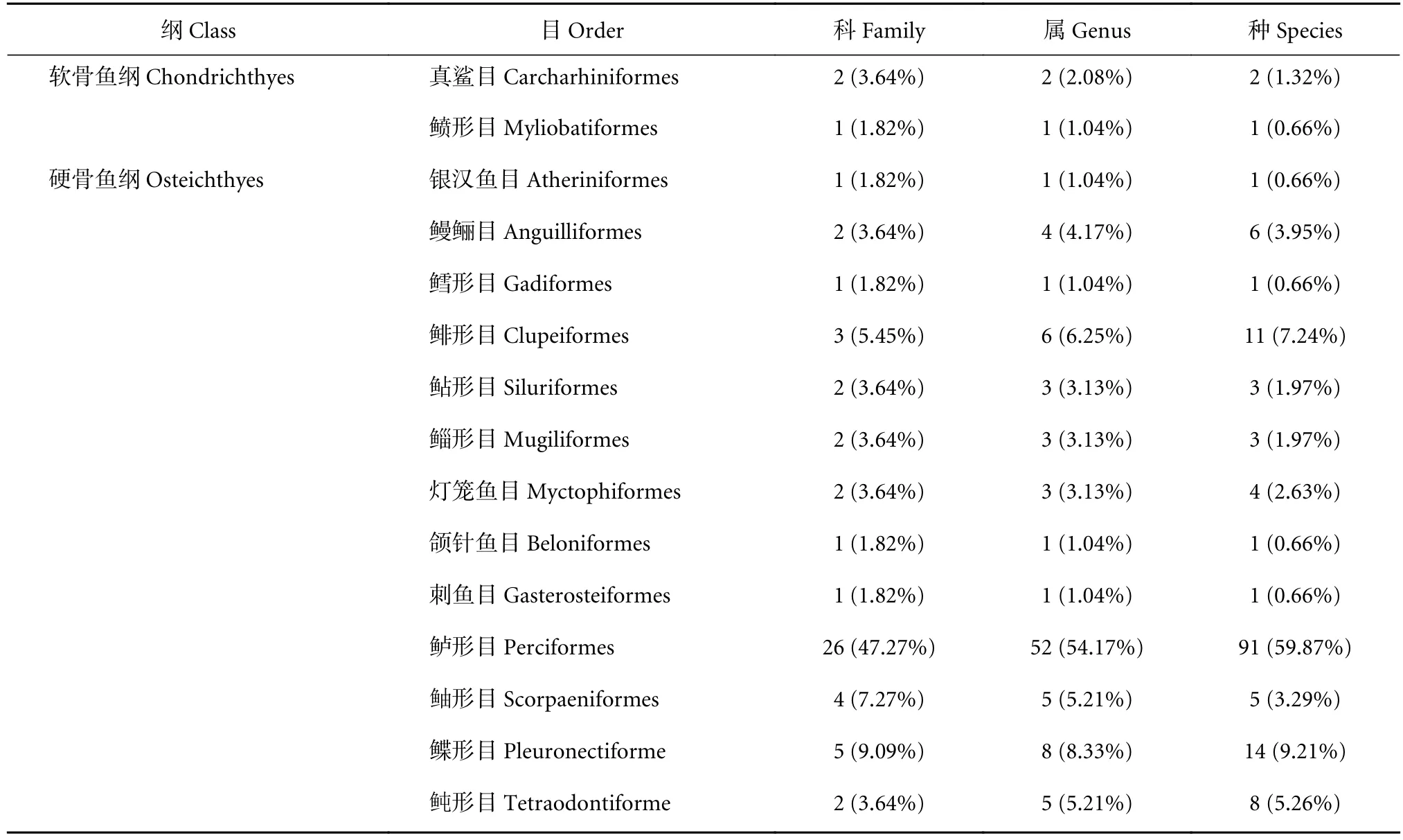
表1 北部湾防城港—钦州近岸渔获鱼类种类组成Table 1 Species composition of fish in Fangchenggang (Qinzhou coastal area of Beibu Gulf)
该海域鱼类资源量季节分布呈一定规律性(图2)。2016 年平均生物量是 473.61 kg·km−2,浮动介于 29.76~884.62 kg·km−2,春、夏季明显高于秋、冬季,春、夏、秋和冬季平均生物量依次为884.62、722.04、258.03 和 29.76 kg·km−2;整年度平均丰度为 104.5×103尾·km−2,变化介于 8.1×103~263.2×103尾·km−2,冬季最低,春季最高,春、夏、秋和冬季平均丰度依次为263.2×103、124.8×103、21.8×103和 8.1×103尾·km−2。
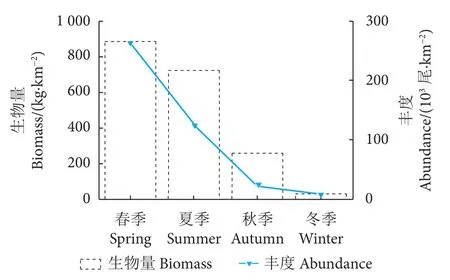
图2 四季鱼类平均生物量和丰度Fig. 2 Mean biomass and abundance of fish in four seasons
各季节鱼类丰度和生物量平面分布差异较明显(图3、图4),春季鱼类资源量集中在近岸,最高生物量和丰度分别出现在S3和S2站位,为3 647.97 kg·km−2和 1 331.1×103尾·km−2,S16 站位生物量和丰度均最低,为 34.11 kg·km−2和 1.0×103尾·km−2;夏季资源量主要分布于南部,S9站位的生物量和丰度均最高,为 2 548.88 kg·km−2和 454.85×103尾·km−2;秋季资源量呈东高、西低势态,最高生物量和丰度分别为 1 113.78 kg·km−2(S19) 和57.5×103尾·km−2(S18),S1 站位生物量和丰度均最低,分别为 99.00 kg·km−2和 6.1×103尾·km−2;冬季资源量较高的站位集中在西部,S2站位生物量最高,为 102.20 kg·km−2,占该季总生物量的 11.84%,S20 站位最低,为 3.75 kg·km−2;丰度最高出现在S12 站位,为 77.6×103尾·km−2,最低为 S17 站位的 0.2×103尾·km−2。
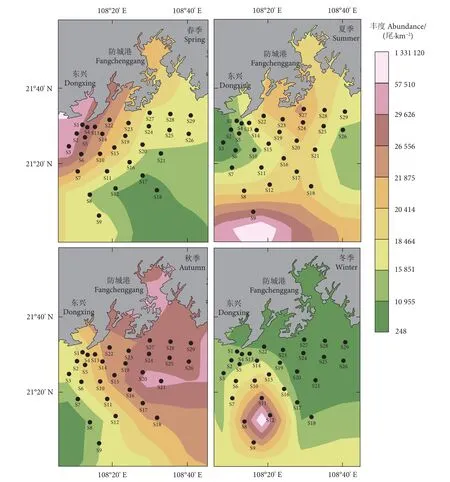
图3 四季鱼类丰度的时空变化Fig. 3 Spatio-temporal changes of fish abundance in four seasons
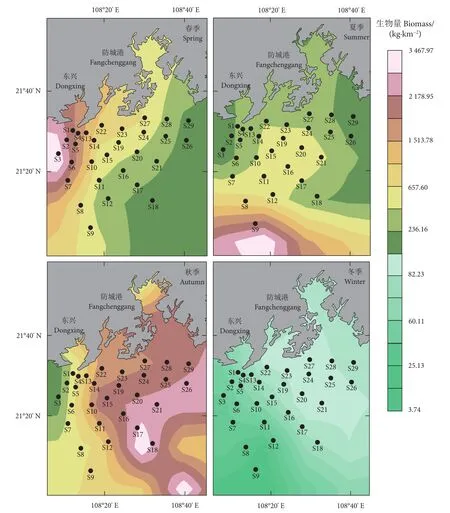
图4 四季鱼类生物量的时空变化Fig. 4 Spatio-temporal changes of fish biomass in four seasons
2.2 多样性指标及优势种的季节变动
北部湾防城港—钦州近岸海域鱼类多样性指标季节间变动幅度较大 (图5)。鱼类多样性指数变化介于 (1.952±0.697)~(2.590±0.567),最高值出现在冬季的S27站位 (3.624),最低是秋季的S25站位(0.637)。均匀度指数介于 [(0.481±0.175),冬季 ]和[(0.699±0.198),春季],秋季的S25站位为谷值(0.172),春季的S16站位为峰值 (0.973)。丰富度指数方面,变化介于 (2.303±0.638)~(3.114±0.543),最高值出现在冬季S3站位 (4.393),冬季的S12站位最低 (1.062)。总体来看,该海域鱼类多样性指标在春、夏季较高,冬季最低。
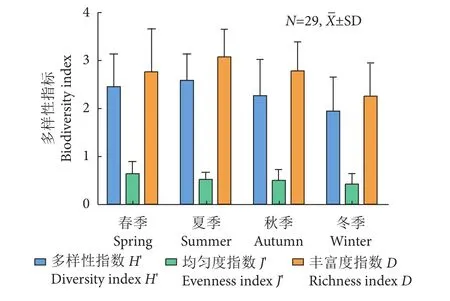
图5 北部湾防城港—钦州近岸各季节鱼类群落多样性指数Fig. 5 Biodiversity index of fish community in Fangchenggang (Qinzhou coastal area of Beibu Gulf) in four seasons
调查结果显示,4季共发现9种优势鱼类,夏季优势种数最高 (5种),冬季最低 (2种) (表2)。二长棘鲷 (Paerargyrops edita) 为 4 季共有优势种,鹿斑鲾 (Secutor ruconius) 为 3 季共有优势种,多齿蛇鲻 (Saurida tumbil) 和褐菖鲉 (Sebastiscus marmoratus) 为2季共有优势种。矛尾虾虎鱼 (Chaeturichthys stigmatias) 为春季第一优势种,平均丰度为145.3×103尾·km−2;夏季第一优势种鹿斑鲾的出现频率为96.55%;多齿蛇鲻在秋季平均渔获率为207尾·h−1,是该季的第一优势种;冬季共捕获第一优势种褐菖鲉1 765尾,占该季总丰度的31.01%,但其生物量仅为 0.13 kg·km−2。经济价值最大的为二长棘鲷 ,是北部湾典型的传统经济种类,蓝圆鲹(Decapterus maruadsi) 和多齿蛇鲻也具有一定的经济价值,其余大部分种类经济价值较低。
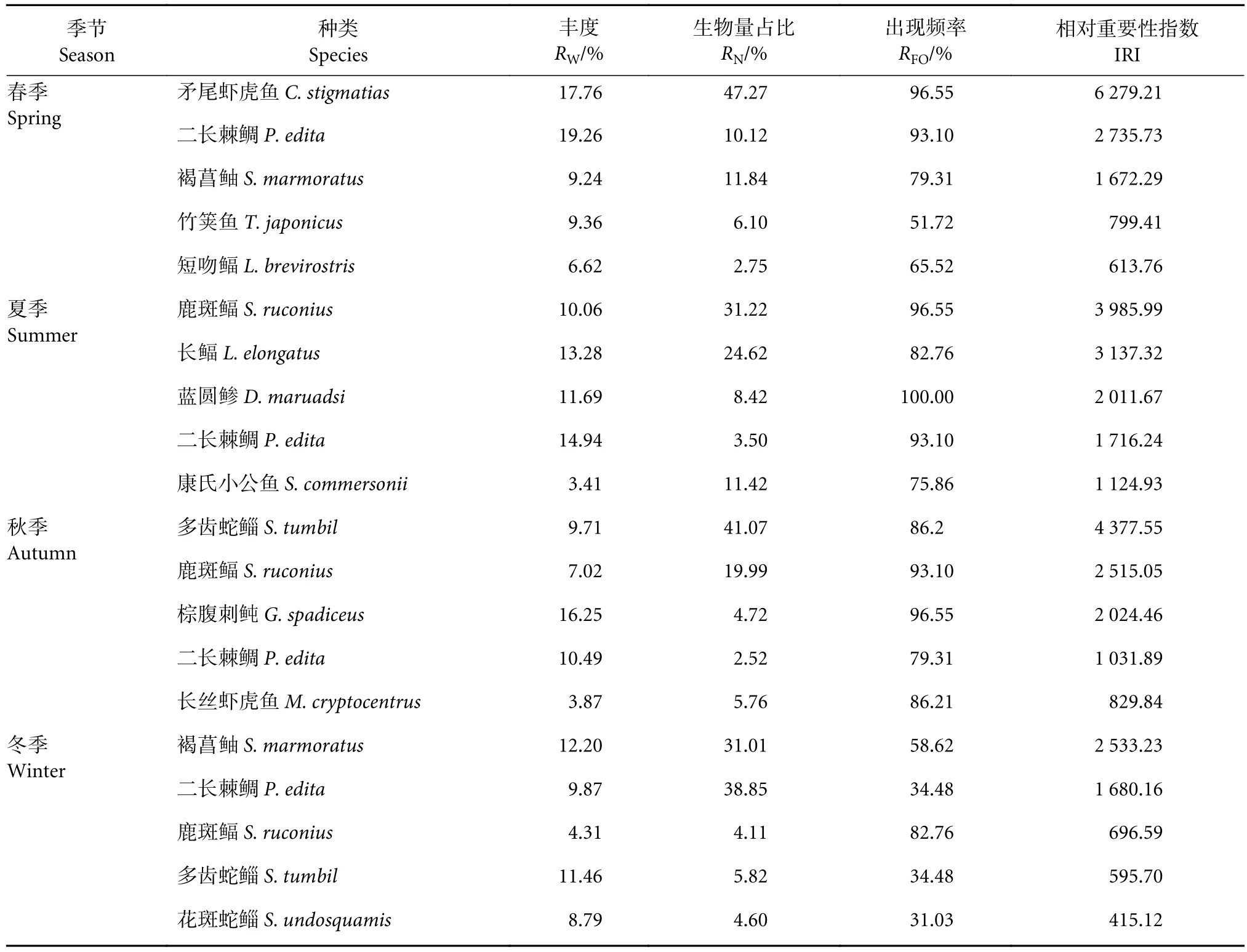
表2 各季节相对重要性指数前5的鱼类优势种Table 2 Importance relative index of five dominated fish species in four seasons
2.3 鱼类组群的划分
本研究中NMDS胁迫系数为0.2,表明组群划分具有可信度,北部湾防城港—钦州近岸鱼类群落可分为4个组群 (图6、图7)。组群I由33个站位构成,其中27个春季站位,6个冬季站位,共发现92种鱼类,平均生物量和丰度为799.09 kg·km−2和 239.7×103尾·km−2,以矛尾虾虎鱼、二长棘鲷和褐菖鲉为优势鱼种,共占该季总丰度的72.93%。组群Ⅱ主要出现于夏季,由30个站位构成,共发现88种鱼类,平均生物量和丰度为704.18 kg·km−2和 121.6×103尾·km−2,短体银鲈 (Gerres abbreviates)、冠鲽 (Samaris cristatus)、棘头梅童鱼 (Collichthys lucidus)、麦氏犀鳕 (Bregmaceros macclellandi)、白氏银汉鱼 (Allanetta bleekeri)、大海马(Hippocampus kelloggi)、大鳞鳞鲬 (Onigocia macrolepis) 和丝鳍美尾䲗 (Calliurichthys dorysu) 为该组所特有。组Ⅲ出现在秋、冬季,由35个站位组成,秋季站位占82.86%,共发现108种鱼类,平均生物量和丰度为 231.51 kg·km−2和 20.5×103尾·km−2,优势种为多齿蛇鲻和鹿斑鲾,占总丰度的58.29%。组群Ⅳ仅现于冬季,由16个站位构成,共捕获63种鱼类,平均生物量和丰度为19.41 kg·km−2和 1.5×103尾·km−2,圆腹鲱 (Dussumieria elopsoides)、峨眉条鳎 (Zebrias quagga)、斑尾复虾虎鱼 (Synechogobius ommaturus)、暗带双线䲗 (Diplogrammus xenicus) 和短尾大眼鲷 (Priacanthus macracanthus) 为该组所特有。
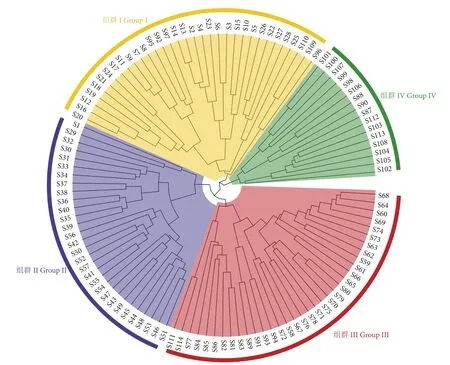
图6 2016年北部湾防城港—钦州近岸鱼类聚类图Fig. 6 Cluster analysis of fish species collected from Fangchenggang (Qinzhou coastal area of Beibu Gulf) in 2016
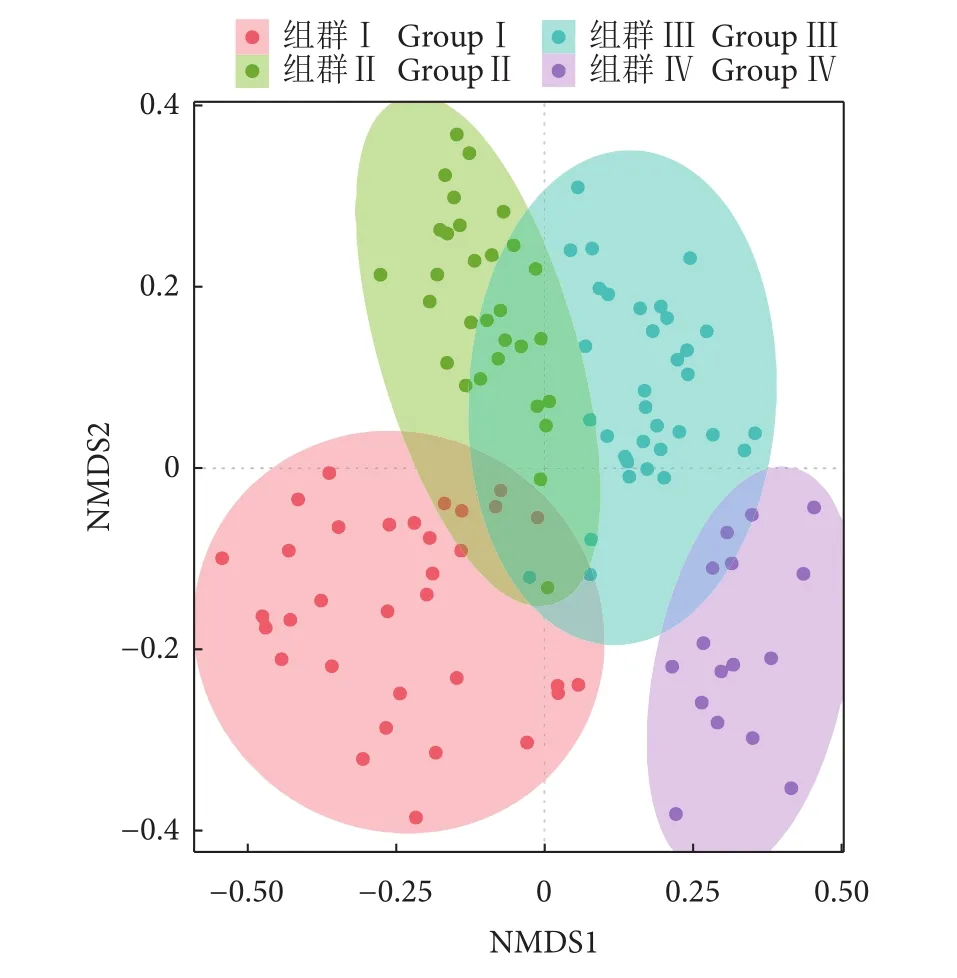
图7 鱼类群落结构NMDS分析Fig. 7 Non-metric multidimensional scaling analysis of fish community
ANOSIM分析结果表明,鱼类组成在各组群间差异极显著 (R=0.703~0.982,P<0.01)。组Ⅱ平均相似度最高 (32.49%),组Ⅰ最低 (18.27%)。二长棘鲷、鹿斑鲾、多齿蛇鲻和圆额金线鱼 (Nemipterus metopias) 依次为4个鱼类组群的典型种,其贡献率分别为28.42%、36.46%、39.23%和12.58%。组群间相异性介于 89.58% (组Ⅱ、Ⅲ) 和 98.02% (组Ⅰ、Ⅳ),长鲾 (Leiognathus elongates) 是促使组Ⅰ与各组群差异的主要分歧种,组Ⅱ与各组间差异的主要分歧种为矛尾虾虎鱼,组Ⅲ、Ⅳ与各组主要分歧种分别为多齿蛇鲻和长丝虾虎鱼 (Myersina cryptocentrus)。
2.4 鱼类群落与环境因子的关系
表、底层温度在季节间差异极显著 (P<0.01),表层温度介于 12.92 ℃ (冬季 S28) 和 31.69 ℃ (夏季S17) ;底层平均温度在冬季最低 [(14.22±0.59) ℃,图8],夏季最高 [(30.45±0.42) ℃];表层平均盐度介于 (28.72±0.88) (夏季) 至 (30.63±0.37) (秋季)间,除春、冬季间差异不显著 (P>0.05) 外,其他季节间差异均极显著 (P<0.01);底层盐度的谷、峰值分别现于夏季的S25和S14站位,依次为28.52、32.02,除夏、冬季间差异不显著 (P>0.05) 外,其余两两季节间差异均极显著 (P<0.01)。深度季节间差异不显著 (P>0.05),但站位间差异极显著 (F=39.842,P<0.01)。pH、溶解氧均在春、夏季间差异不显著 (P>0.05),在其他季节间差异显著 (P<0.05),其中 pH 波动介于 (8.06±0.08) (夏季) 至(8.31±0.06) (冬季) 间,溶解氧的峰、谷值分别出现在冬季的 S18 站位 (10.44) 和春季的 S16 站位 (5.68)。
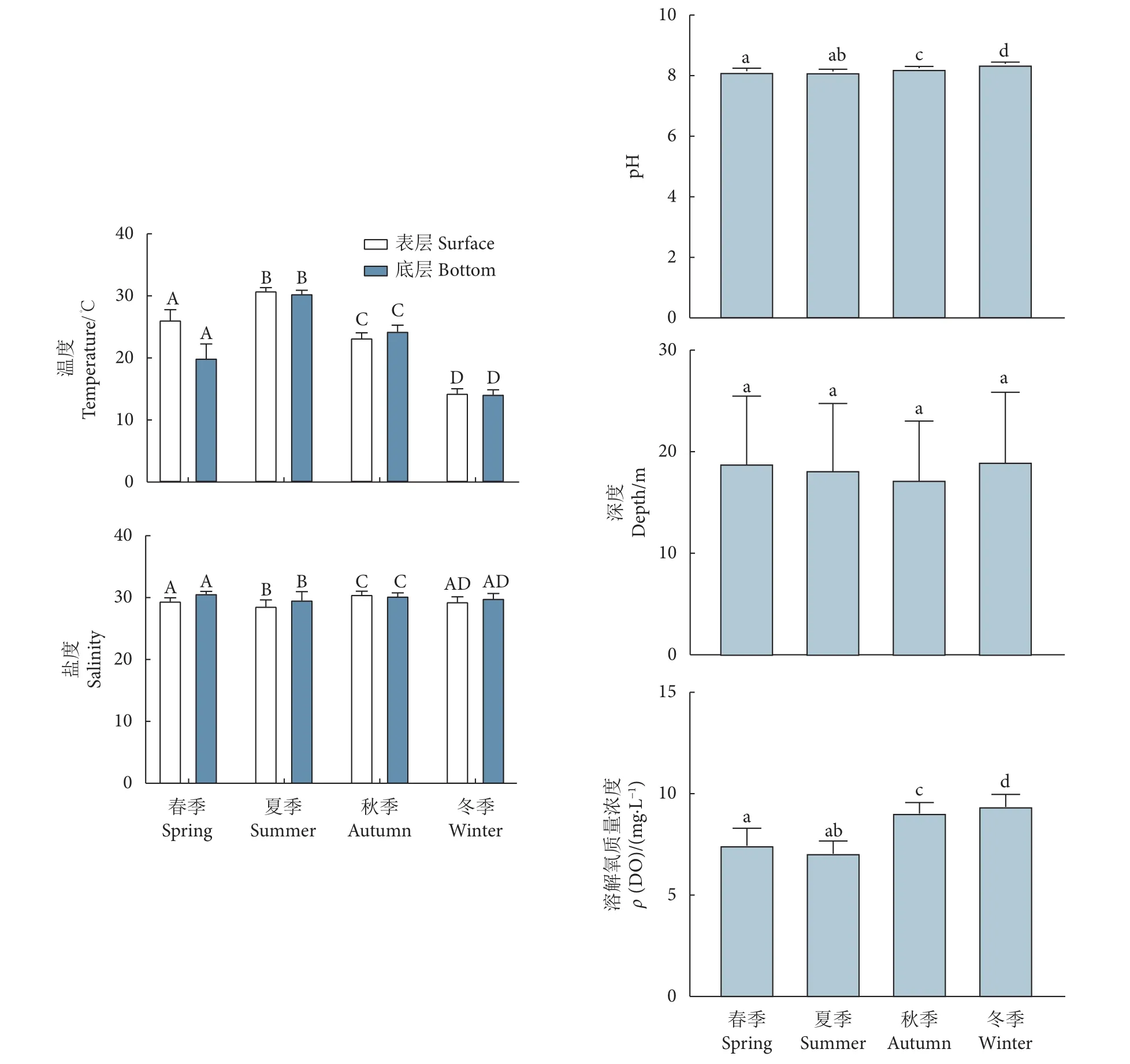
图8 北部湾防城港—钦州近岸四季环境因子注:不同的大写字母表示差异极显著 (P<0.01);不同的小写字母表示差异显著 (P<0.05)。Fig. 8 Environmental factors in Fangchenggang (Qinzhou coastal area of Beibu Gulf) in four seasons Note:Different uppercase and lowercase letters on the columns indicate extremely significant difference (P<0.01) and significant difference (P<0.05).
各季节环境因子相关分析结果见图9。春季,溶解氧与pH间正相关性最显著 (R=0.842,P<0.01),表层温度与表层盐度的负相关关系最显著(R=−0.634,P<0.01);夏季,底层盐度与深度间正相关性最显著 (R=0.935,P<0.01),底层温度与深度间负相关关系最显著(R=−0.892,P<0.01);秋、冬季环境因子间均不存在显著负相关性。秋季,表层盐度与深度间正相关性最显著 (R=0.828,P<0.01);冬季,底层温度与底层盐度间正相关性最显著(R=0.952,P<0.01)。
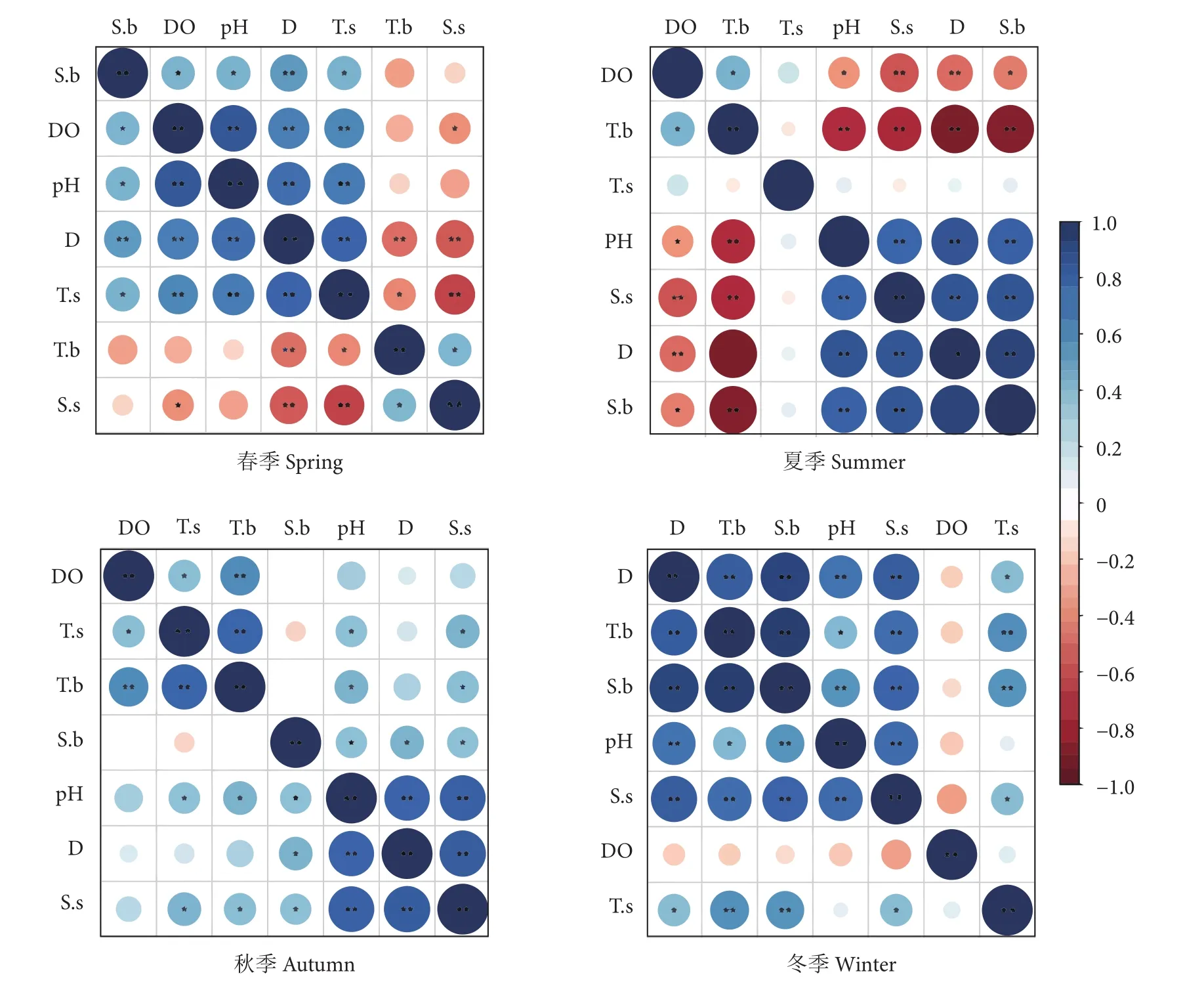
图9 各季节环境因子相关性热图Fig. 9 Correlation heat maps of environmental factors in four seasons
根据图10中环境因子的矢量长度可知,底层温度 (与第1轴存在较强相关性,R=0.76) 和溶解氧 (与第2轴存在较强相关性,R=−0.63) 对鱼类分布具有较大影响。第1、2轴分别解释了12.4%和7.5%的物种变化率,第1轴将鱼类划分为好高温 (轴右侧) 和耐低温 (轴左侧) 两种,组Ⅱ优势种康氏小公鱼 (Stolephorus commersonii)、鹿斑鲾、长鲾均位于轴右侧,与温度具有较强的正相关性,组Ⅲ优势种多齿蛇鲻和组Ⅳ优势种圆额金线鱼位于轴左侧,与温度存在较强的负相关性。第2轴将鱼类划分成好高氧 (轴下部分) 和喜低氧 (轴上部分) 两种,竹筴鱼、海鳗 (Muraenesox cinereus) 和半滑舌鳎 (Cynoglossus semilaevis) 等密度与溶解氧含量成正比,组I优势种矛尾虾虎鱼与溶解氧含量成反比。此外,深度和底层盐度 (与轴1的相关系数分别为0.58、0.42) 也影响着鱼类资源格局,且两者存在较强相关性 (R=0.94),油魣 (Sphyraena pinguis)、黄鳍马面鲀 (Thamnaconus hypargyreus) 和短带鱼 (Trichiurus brevis) 的资源密度与深度、底层盐度成正比,多鳞、短吻鲾 (L. brevirostris) 和鰕虎鱼科则相反。蒙特卡罗检验表明,7种环境因子变量与鱼类分布的相关性显著 (P<0.05)。
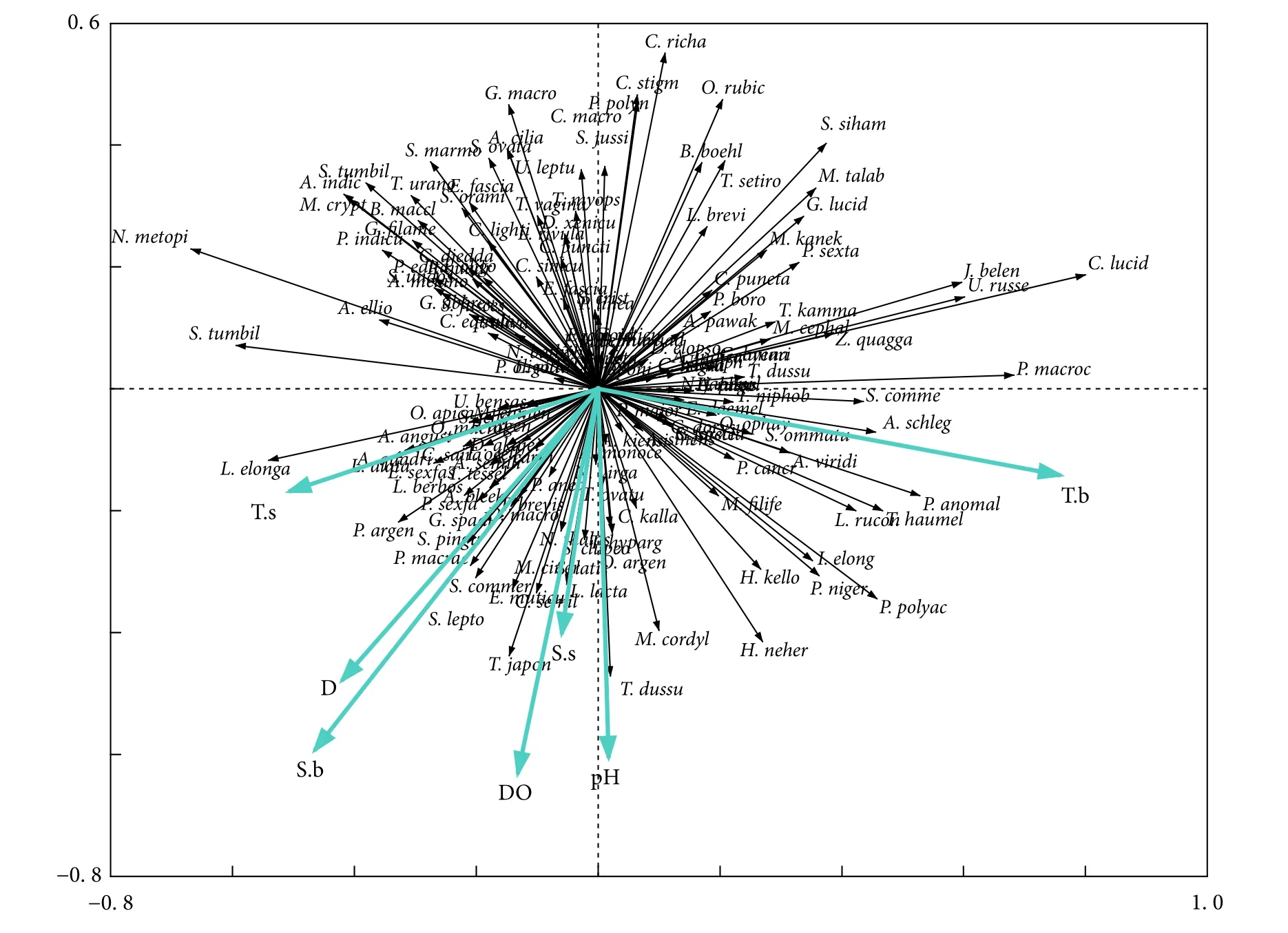
图10 基于冗余分析的鱼类群落和环境因子排序图Fig. 10 Ordination bi-plot of fish communities and environmental factors based on redundancy analysis
3 讨论
3.1 鱼类群落时空格局
鱼类种数的多寡受生境个数及规模的影响,优质的生存环境可提高鱼类的成活率,而且海域所处纬度也会影响鱼种数目[30-31]。此次调查鱼种类数少于较孙典荣[31]、王雪辉等[12]分别于2001、2002和2007年在北部湾海域捕获的种数 (209、244和323种),但比北部湾西北部海域和防城河口湾种类数 (125和95种) 多[19-20]。季节变化上,与防城河口湾[20]、北部湾东北部[21]及西北部[19]海域鱼类资源调查结果一致,该海域种类数春季最多,冬季最少。
北部湾防城港—钦州近岸海域鱼类资源季节分布不均,春季鱼类丰度较高,因为该季多数鱼类正处于繁殖期,以北部湾主要优势种二长棘鲷为例,成鱼于冬季洄游至近岸产卵,幼鱼于春季充分利用湾内富足的食物及天然的保护屏障度过幼鱼期[32],新增的幼鱼补充了资源密度。夏季鱼类生物量和丰度亦较高,休渔措施起了重要作用,休渔期间北部湾海域温度较高、饵料丰富,幼鱼得到了充分的生长机会。另外,拖网会破坏底栖生物的生存生境,休渔措施减少了人类活动对海域生态的扰动,以上因素使得夏季鱼类资源得以有效修复[20]。到了秋季,随着幼鱼成长,大部分鱼类开始游向深海生活,地方性鱼类成为优势鱼种,这与傅昕龙等[19]的鱼类资源量在秋季较低的结论一致。冬季鱼类生物量仅占春季的3.45%,季节变化并不会直接改变鱼类群落结构,但会结合某些鱼类的生活习性促使鱼类进行特定活动[33],使得近岸海域渔获物减少。例如,瑞典西部海域近岸鱼群越冬洄游移至深海[3],小口棘隆头鱼 (Centrolabrus exoletus) 和岩梳隆头鱼 (Ctenolabrus rupestris) 在冬季会躲避在岩石缝隙中减少游动[34-35],该海域多种鱼类越冬洄游至湾南部和中西部越冬[19],导致冬季鱼类资源量明显降低。
鱼类资源的空间分布与水系水团的消长、食物多寡及获取的难易程度密切相关[33,36]。春季沿岸河流水势由弱转强,为调查海域带来丰厚的陆源营养物,吸引多种鱼类前来索饵和繁殖[19],使得鱼类资源均集中在近岸,何思璇等[20]也指出春季防城河口湾鱼类资源集中分布在湾口。夏、秋季混合水进入湾内使北部湾北部出现气旋式环流,聚集在近岸的鱼类跟随逆时针趋势水团游向南部和东南部[23]。冬季强烈的东北季风,使温度急剧下降,大部分鱼类游至外海越冬,加之对北部沿岸水体扰动加强,导致地方性鱼类游向西部[37]。
3.2 鱼类多样性及优势种变化
此次调查鱼类平均多样性指数为2.32,与以往研究结果相比处于较低水平[38-40]。1992—1993年北部湾鱼类群落平均多样性指数为3.50,1998—1999年多样性指数冬季最高 (2.98),春季最低 (2.47),2007年平均多样性和均匀度指数分别为3.31和0.57。自20世纪80年代起,中国渔业捕捞能力显著增强,鱼类资源量持续下滑,到了1999年,由于休渔政策、燃油价格上涨等原因,海洋生物资源得到有效恢复,多样性指数在一定时期内呈先下降后上升的趋势[41]。进入21世纪以来,随着人类活动的不断加剧及环境变化,北部湾海域鱼类多样性又呈现下降趋势[39]。
北部湾海域在捕捞压力和环境改变等因素影响下,其优势鱼类结构特征和组成发生了较大变化。1957—1960年,金线鱼属、红笛鲷 (Lutjanus erythopterus) 和大眼鲷属占据优势生态位[41];红笛鲷、黄带绯鲤 (Upeneus sulphureus) 和摩鹿加绯鲤(U. moluccensis) 在 1961—1962 年丰度较高,占该年度总丰度的19.92%[39];1992—1993年调查航次以发光鲷 (Acropoma japonicum)、白姑鱼 (Argyrosomus argentatus) 和长尾大眼鲷 (P. tayenus) 为主要优势种[39];2001—2002年间,短尾大眼鲷、二长棘鲷和大头白姑鱼等占据优势生态位[39];竹荚鱼、大头白姑鱼、发光鲷和二长棘鲷在2007年为常年优势种[12]。较往年结果相比,个体较大的底层中型鱼类数量急剧减少,虾虎鱼科、鲾科等小型低质鱼类成为主要优势种群,此次调查IIR最高的优势种为矛尾虾虎鱼、多齿蛇鲻、鹿斑鲾和长鲾。传统优势经济种寿命长、生长缓慢,易受人为捕捞和生态环境改变的干扰,进而易被繁殖能力强、生长迅速、能在短时间内恢复种群数量和规模的小型种类占据[18],Prista等[41]发现某些个体较小的鱼类资源密度易受网目规格的影响,当网目较大时其资源量易被低估,此调查采用40 mm网目的底拖网,相比王雪辉等[12](39 mm 网目)、傅昕龙[19](20~50 mm网目) 较大,因此该海域鱼类小型化程度可能比调查结果更严重。
3.3 鱼类群落与环境因子的相关性
本研究表明,共有105种鱼类与此次群落划分有关,占总鱼种的69%,主要属于虾虎鱼科、鲾科和鲷科。而Molina等[42]仅利用14种鱼类 (占总种数20%,占总生物量90%) 研究布埃纳文图拉湾鱼类群落分布特征。不同海域的差异致使鱼类群落存在多样的优势度形态,这通常与水文地形环境、饵料分布以及鱼类对海湾的利用方式有关[43]。与Whitfield[44]对巴拉那瓜湾鱼类群落划分特征类似,北部湾防城港—钦州近岸海域鱼类群落的划分具有较明显的季节相关性,85%的组Ⅰ站位分布在春季,97%的组Ⅱ站位出现在夏季,83%的组Ⅲ站位出现在秋季,组Ⅳ站位均出现于冬季。该海域鱼类群落划分的典型种与分歧种存在一定交互性,并且对组内相似性及组间差异性贡献较大的典型种和分歧种多为调查海域优势鱼种,这与王小林[43]对海州湾及邻近海域鱼类群落划分结果一致。
入海河流的径流量以及时空分布格局对近岸海域动、植物资源量和生态过程起到关键作用[45]。学者们认为,入海河流可影响近岸海域初级生产力的水平,改变海洋生物栖息地的范围、海水浊度和盐度,从而间接影响鱼类群落结构与资源分布特征[46-48]。春、夏航次属于丰水期,河流淡水携带大量有机碎屑、无机盐等涌入近岸海域,其中有机碎屑由细菌和真菌分解,被海洋生物吸收利用,磷酸盐、硝酸盐和硅酸盐等无机盐促进浮游生物迅速生长,促进浮游生物食性鱼类 (如沙丁鱼) 资源量的增加[47]。春季,因河流注入的营养物质为幼鱼生长提供食物保障,同时由于强大的水流作用,河口中幼鱼被冲向沿岸海域,故春季鱼类丰度较高的站位多集中于近岸海域[48]。夏季,持续涌入的淡水使得河口区域的透明度和盐度显著下降,鱼类的优质生境变小或消失,多种鱼类以寻求适宜盐度及有利的生存和繁殖环境,选择从河口向近海扩散[47],因此夏季近海鱼类资源得到一定补充。与以往研究相似,此次调查秋季中部和东部近岸站位 (靠近两江入海口) 资源量较高。秋季处于枯水期,干旱引起河口水质恶化,食物供应减少,单位捕捞努力量(Catch per unit of effort, CPUE) 明显下滑,栖息于河口的鱼类被迫向近岸迁移[48]。
鱼类群落的划分除受其生活习性影响外,还与海水环境因子的作用密切相关。由于水域环境特征不同,主要的环境影响因子也不尽相同[12,49]。RDA分析结果显示,温度对北部湾防城港—钦州近岸海域鱼类分布具有较大作用,这与何思璇等[20]提出的温度是影响防城河口鱼类群落数量和结构的最主要环境因子一致。由于不同鱼类的温度耐受性存在差异,所以温度可通过不同鱼类对耐受程度的差别影响资源密度的分布格局[13],此研究结果表明,多齿蛇鲻和圆额金线鱼位于第1轴的左侧,其资源量随着温度的升高而降低,这与Poxton和Allouse[49]对鲑的研究结果一致,鲑的最大耐热温度为25 ℃,超过这个限度会出现死亡现象。另外,康氏小公鱼和鹿斑鲾位于轴右侧,其密度随着温度的降低而降低。类似地,大西洋白姑鱼通常会在邻近塔霍河河口区域过冬,由于该水域 2000—2001年冬季平均气温低于15 ℃,导致这期间并未发现该鱼种[34]。温度还可以和其他环境因子互作,综合影响鱼类分布;Pomfret等[50]发现在低氧条件下,英国北部入海河口的鱼群划分受到温度的明显界定。溶解氧含量也是重要的影响因素,海域内鱼类饵料丰富度与溶解氧含量存在一定关系[51-52],胡翠林等[52]发现当海域中浮游动物含量增多时溶解氧含量普遍降低,此时底层鱼类为获取充足饵料而提高对低溶解氧的耐受性。深度与轴1、2均具较好相关性,深度与生物体关系微妙,一般来说,深度只能表示水文、地形方面的异同,不能直接说明与生物间的相关性[53],但深度可通过影响其他因素间接影响鱼类分布,Prista 等[41]认为葡萄牙沿岸海域鱼类群落结构受季风、浪涌和底质类型 (礁石多寡) 的直接影响,而上述因素又与深度存在较强相关性,所以深度间接影响了鱼类分布。邱永松[54]指出温度、盐度等环境因子随着深度的改变而变化,不同深度可利用与之相应的多样化的环境因素限定鱼类的活动范围。此调查发现,油魣、黄鳍马面鲀、多鳞和短吻鲾的种群密度与底层盐度密切相关。海湾鱼类群落可看作广盐性鱼类、海水鱼类和淡水鱼类在适应一定程度的盐度动态变化和调整生活习性后的复杂组合,虽然大多数鱼种对盐度的适应范围较广,但是其仍习惯于在某一较窄盐度区间内生存[55],Renfro[56]调查阿蓝萨斯郡河口鱼类资源量时指出,某些鱼类因受盐度频繁波动干扰,大多选择向盐度含量较低的水域迁移,特殊情况下会因不适应高盐环境而死亡。
4 结论
本研究结果表明,2016年北部湾防城港—钦州近岸海域共捕获鱼类15目55科152种。与以往调查相比,鱼类多样性水平较低,鱼类小型化程度严重。鱼类资源量具明显的时空分布规律,二长棘鲷、鹿斑鲾、多齿蛇鲻和褐菖鲉为该海域主要优势种。鱼类群落结构存在较明显的季节分布特征,其中组Ⅰ (春季为主) 和组Ⅱ (夏季为主) 的平均生物量和丰度较高,构成组内相似及组间相异的典型种和分歧种,具有交互性,且多为调查海域优势鱼种。不同季节环境因子间的相关程度不同,其中表、底层温度在各季节间均出现极显著差异 (P<0.01)。此外,底层温度、溶解氧、深度和底层盐度是影响北部湾防城港—钦州近岸海域鱼类分布格局的主要环境因子。
猜你喜欢
水运工程(2022年10期)2022-11-02
广西电业(2022年7期)2022-10-13
水上消防(2021年4期)2021-11-05
食品安全导刊(2021年20期)2021-08-30
北方音乐(2019年10期)2019-07-10
学苑创造·B版(2019年2期)2019-02-19
中国-东盟博览(旅游版)(2018年7期)2018-05-14
军营文化天地(2017年12期)2017-03-01
艺术教育(2011年10期)2011-12-19
