
凡纳滨对虾含酪氨酸酶结构域血蓝蛋白的免疫功能
2022-09-28 01:40王子昂汤俊宇杨林蔚
广东海洋大学学报 2022年5期
张 锐,王子昂,汤俊宇,杨林蔚
(1.广东海洋大学食品科技学院/ 现代生化中心,广东 湛江 524088;2.中山大学生命科学学院有害生物控制与资源利用国家重点实验室,广东 广州 510275;3.南方海洋科学与工程广东省实验室(珠海),广东 珠海 519000;4.广东海洋大学深圳研究院,广东 深圳 518108)
血蓝蛋白是节肢动物和软体动物血淋巴中的一种含铜呼吸蛋白,在氧运输、能量贮存、渗透压维持、蜕皮和表皮固化调节等方面发挥重要功能。研究发现,甲壳动物的血蓝蛋白也有免疫活性,参与机体对多种病原的免疫防御[1-3]。在NCBI Genbank 数据库中,每个甲壳类物种往往有多个序列有明显差异的血蓝蛋白编码基因,表明其体内的血蓝蛋白可能以多个基因表达产物混合物的形式出现,从而拥有结构和功能上的多样性。因此,有必要从单一基因的角度去探讨血蓝蛋白的免疫功能。
凡纳滨对虾(Litopenaeus vannamei)是全球主要养殖虾种之一,也是研究无脊椎动物免疫的重要对象。从NCBI 上公布的凡纳滨对虾基因组中共发现16个血蓝蛋白基因,其蛋白结构域均拥有典型氨基端的Hemocyanin_N结构域和羧基端的Hemocyanin_C结构域,其中的一种血蓝蛋白还含有酪氨酸酶结构域(Tyrosinase-domain),与其他血蓝蛋白家族成员存在明显差异,故命名为含酪氨酸酶结构域血蓝蛋白(Tyrosinase-domain containing hemocyanin,TDCH)。本研究克隆该蛋白的基因tdch,分析其在对虾抗副溶血弧菌免疫中的作用,为深入探索虾类血蓝蛋白功能以及病害防治提供基础。
1 材料和方法
1.1 虾和病原体
凡纳滨对虾(约10 g)购自深圳某对虾养殖场,在约28 ℃富含氧气的地下水(盐度2.0) 中适应一周。正式实验前,对5%的对虾进行随机抽样,采用文献[4-5]方法,通过PCR 检测,确保没有副溶血弧菌(Vibrio parahaemolyticus)和白斑综合征病毒(White Spot Syndrome Virus,WSSV)污染。副溶血弧菌和白斑综合征病毒按照文献[6-7]方法制备。
1.2 凡纳滨对虾tdch基因片段的扩增及生物信息学
从凡纳滨对虾基因组和转录组数据库获得tdch基因序列[8],以凡纳滨对虾鳃组织cDNA 为模板,利用引物对TDCH-ORF-F/R(表1)扩增获得tdch编码区序列。PCR 反应体系(20 μL):Premix Taq 10 μL,ddH2O 7.5 μL,cDNA 模板0.5 μL,上下游引物各1 μL。PCR 产物经凝胶电泳检测、目的条带切胶回收,连接至pMD-19T,转化至DH-5α 感受态细胞。挑取单菌落进行菌落PCR 检测,阳性菌送至天一辉远有限公司广州测序部测序。获得tdch序列后,运用SMART 网站(http://smart.embl-heidelberg.de/)分析tdch的结构域,运用AlphaFold 预测tdch的三维结 构[9]。用Clustal W 1.8 将tdch序列与Genbank 数据库中凡纳滨对虾其他血蓝蛋白序列进行比对。采用MEGA 5.0 软件通过邻接法(Neighbor-joining)进行进化树分析,具体参数如文献[10-11]。

表1 引物名称及序列Table 1 Primers and their sequences
1.3 实时荧光定量PCR
取健康凡纳滨对虾,分别取肌肉、肝胰腺、鳃、胃、表皮、触角、心脏、肠、幽门盲囊、眼柄和神经索,样品剪碎至约2 mm,置于RNAlater(Invitrogen)中,血淋巴离心后用RNAlater 重悬。置4 ℃下12 h,放于-80 ℃冰箱保存,用于分析tdch组织分布。每组样品中为15 尾虾的组织混合物。每个组织至少取3组。
取凡纳滨对虾700 尾,随机分为7 组,每尾虾分别于第二腹肢基部位置用1 mL 的胰岛注射器分别注射0.1 μg/μL 脂多糖(LPS)、0.1 μg/μL Poly (I:C)、2.0×103cfu/μL 金黄色葡萄球菌(Staphhylococcu saureus)、2.0×103cfu/μL 副溶血弧菌、2.0×103cfu/μL黑曲霉菌(Aspergillus niger)、2.0×104拷贝/μL WSSV 各50 μL,对照组注射同剂量磷酸盐缓冲液(PBS,pH=7.2)[12]。在注射0、4、12、24、48、72、96 h时,分别随机选取虾6 尾,采集其鳃和血细胞,置于RNALater 存 储。RNA 提 取、cDNA 合成和实时荧光定量PCR 安照文献[13]的方法进行。根据tdch基因开放阅读框的保守区,用Primer Premier 5.0 设计引物qRT-TDCH-F、qRT-TDCH-R,对照组EF-1α基因(Genbank 登录号:GU136229)为管家基因,引物如表1。qRT-PCR 反应体系(10 μL):引物(10 μmol/L)各0.5 μL,Mix 5 μL,ddH2O 2.5 μL,cDNA 0.5 μL。反应条件:95 ℃4 min;95 ℃20 s、60 ℃30 s、72 ℃2 min,35 个循环;72 ℃10 min,95 ℃15 s。用2-ΔΔCt法计算相对表达量,用GraphPad Prism 7 软件进行单因素方差分析。
1.4 RNA干扰
用T7 RiboMAX™Express RNAi System 试剂盒(Promega)依照文献[14]及试剂盒说明书合成tdch基因特异的dsRNA,阴性对照为绿色荧光蛋白基因(GFP)的dsRNA,所用引物见表1。取体质健康、大小均一的凡纳滨对虾(约5 g)暂养后,将dsRNA用PBS 稀释后,向对虾的第二腹肢基部注射50 μL 1 μg/μL dsRNA。注射后48 h 随机取虾6 尾于RNAlater 中,-80 ℃保存,用于检测dsRNA 对对虾tdch的沉默效果。另外,在注射dsRNA 48 h 后,用1×106cfu/mL 的副溶血弧菌对凡纳滨对虾进行人工感染,每间隔2 h 统计对虾死亡数量。
1.5 统计分析
采用SPSS 16.0 软件进行统计学分析。计算3次检测的平均值和标准差。采用t检验分析两组数值间差异的显著性。实验组与对照组间存活率差异的显著性则用Mantel-Cox log-rankχ2计算。
2 结果
2.1 凡纳滨对虾tdch结构
克隆得凡纳滨对虾tdch(GenBank 登录号:ROT60816.1)的编码区长1 656 bp,编码551 个氨基酸,分子质量为63.37 ku,理论等电点为7.74。在1~17 氨基酸处含有一个信号肽,21~147 氨基酸处含有一个Hemocyanin_N 结构域,308~410 氨基酸处含有一个酪氨酸酶(Tyrosinase)结构域,413~522 氨基酸处含有一个Hemocyanin_C结构域(图1、2)。

图1 凡纳滨对虾tdch及其推导的氨基酸序列Fig.1 Tdch of Litopenaeus vannamei and its deduced amino acid sequence
2.2 凡纳滨对虾tdch与其他血蓝蛋白的差异分析
将凡纳滨对虾tdch的mRNA 与其对应的DNA进行比对,发现tdch编码区序列含有两个外显子及一个内含子。tdch氨基端含有一个Hemocyanin_N结构域,羧基端含有一个Hemocyanin_C 结构域,符合血蓝蛋白家族的结构特征,与NCBI 数据库中凡纳滨对虾其他血蓝蛋白不同的是,其结构域中间含有典型的酪氨酸酶结构域(图3)。构建的系统进化树(1 000 倍bootstrap)表明,凡纳滨对虾tdch单独聚为一支,在进化关系上远离凡纳滨对虾其他血蓝蛋白,提示tdch可能与以往研究的血蓝蛋白不同,为新的血蓝蛋白家族成员(图4(A))。凡纳滨对虾tdch与其他物种血蓝蛋白的系统进化树显示,凡纳滨对虾tdch与斑节对虾(Penaeus monodon)、日本囊对虾(Marsupenaeus japonicus)的血蓝蛋白聚为一个分支,亲缘关系较为接近,提示凡纳滨对虾tdch在进化较为保守(图4(B))。

图2 凡纳滨对虾tdch的结构Fig.2 Schematic diagram of tdch of Litopenaeus vannamei

图3 凡纳滨对虾不同血蓝蛋白的mRNA序列与对应的DNA序列比对结果及相应的结构Fig.3 Schematic diagram of different L.vannamei hemocyanins and the comparison results of their mRNA sequences and corresponding DNA sequences

图4 凡纳滨对虾不同血蓝蛋白(A)及甲壳动物血蓝蛋白(B)的系统进化树Fig.4 Phylogenetic trees of different hemocyanins in Litopenaeus vannamei(A)and TDCH in crustacean(B)
2.3 凡纳滨对虾TDCH的三维结构预测
经预测凡纳滨对虾TDCH 蛋白的三维结构简单,主要以α螺旋结构为主,包含4处β折叠(图5)。
2.4 凡纳滨对虾tdch基因组织表达分析
图6 表明,tdch在凡纳滨对虾各组织中均有表达,在眼柄、肌肉、触角和肠道中表达量较高,在肝胰腺、表皮、心脏和幽门盲囊中表达量较低。

图6 凡纳滨对虾tdch的组织表达Fig.6 Expression of tdch in different tissues of Litopenaeus vannamei
2.5 凡纳滨对虾tdch时序表达
2.5.1 凡纳滨对虾免疫后tdch在血细胞中的表达变化 经6 种免疫原刺激后,凡纳滨对虾tdch在血细胞中表达量均显著上升。其中,副溶血弧菌、金黄色葡萄球菌、白斑综合征病毒和Poly (I:C) 刺激后,tdch在血细胞中4 h被极显著激活(P<0.01),与对照组相比分别上调151.8、194.8、621.8 和1 254.9倍,并且在4~96 h 一直保持在高表达状态。黑曲霉菌刺激后,tdch在血细胞中24 h 被显著激活(P<0.01),与对照组相比上调2.8倍,并在48 h表达量骤升,与对照组相比上调322.6倍,并且在48~96 h一直保持在高表达状态。LPS 刺激下,tdch在血细胞中24 及72 h 被显著激活(P<0.01),与对照组相比分别上调3.5倍、1.8倍(图7)。

图7 不同免疫原感染后凡纳滨对虾tdch在血细胞中的表达Fig.7 Expression of tdch in hemocyte in Litopenaeus vannamei challenged by different immunogens
2.5.2 凡纳滨对虾免疫后tdch在鳃组织中的表达变化 图8可见,经副溶血弧菌、金黄色葡萄球菌、黑曲霉菌、白斑综合征病毒、LPS、Poly (I:C)或PBS 刺激后,凡纳滨对虾tdch在鳃组织中表达变化较小,在副溶血弧菌和黑曲霉菌刺激24 h时被显著激活(P<0.01),与对照组相比分别上调2.1 倍和2 倍;在金黄色葡萄球菌刺激12 h时被显著激活(P<0.01),与对照组相比上调1.7 倍,刺激4 h 及24~96 h 时表达量没有影响;在白斑综合征病毒刺激48 h 及96 h 时被显著激活(P<0.01),与对照组相比分别上调2.4倍和1.9倍;在LPS刺激24 h及72 h时被显著激活(P<0.01),与对照组相比分别上调1.9倍和1.9倍;Poly(I:C)刺激对tdch在鳃组织中的表达没有影响。

图8 不同免疫原感染后凡纳滨对虾tdch在鳃组织的表达Fig.8 Expression of tdch in gill of Litopenaeus vannamei challenged by different immunogens
2.6 凡纳滨对虾tdch在副溶血弧菌感染中的作用
图9 表明,经RNAi 后,在血细胞中tdch被成功敲降,与对照组相比敲降效率达到70%。图10 表明,注射dsRNA 48 h 后的对虾经副溶血弧菌攻毒后,与GFP dsRNA 对照组相比,TDCH dsRNA 组的存活率显著下降(Mantel-Cox log-rankχ2=8.159,P=0.043 <0.05),表明tdch的表达增强凡纳滨对虾对副溶血弧菌的抗性。
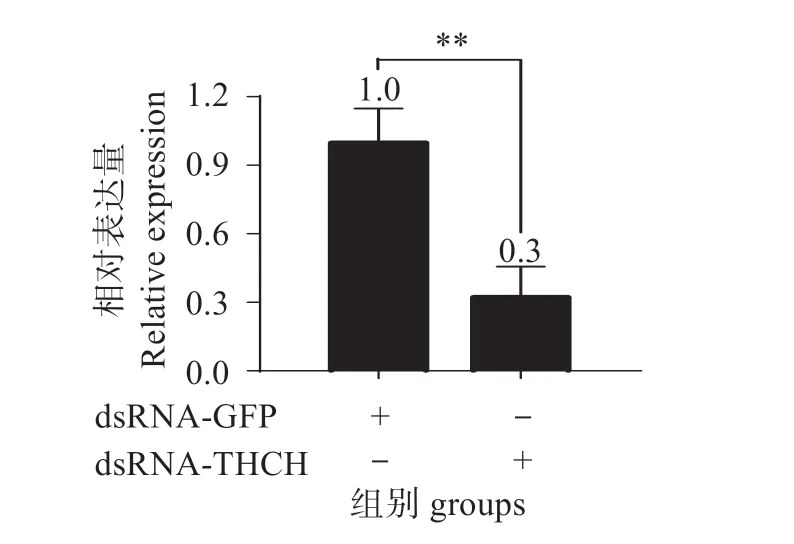
图9 Tdch在血细胞中的敲降效率Fig.9 Knockdown efficiency of tdch in hemocyte

图10 Tdch抑制副溶血弧菌感染Fig.10 Tdch inhibits Vibrio parahaemolyticus infection
3 讨论
在凡纳滨对虾中,TDCH 蛋白氨基端的Hemocyanin_N 结构域和羧基端Hemocyanin_C 结构域与其他血蓝蛋白类似[15],TDCH 还含有一个典型的酪氨酸酶结构域。酪氨酸酶是一种黑色素通路中的结合铜离子的酶,具有酚氧化酶活性,参与体液免疫反应、损伤修复等重要生理活动[16-17]。有研究表明,血蓝蛋白可能是在大约55亿年前由酚氧化物酶进化而来[18]。目前有研究证实血蓝蛋白可能保留有酚氧化酶的活性,例如在软体动物中,散盖大蜗牛(Helix aspersa)的血蓝蛋白在SDS 作用下能够表现出酚氧化酶活性[19];也有报道称凡纳滨对虾的血蓝蛋白也具有酚氧化酶活性,并且还具有细胞凝集和细菌凝集活性[20-21],在外界病原菌刺激下能够发挥类似酚氧化酶的活性参与免疫防御。血淋巴酚氧化酶活力包括血蓝蛋白酚氧化酶活力和血细胞酚氧化酶原释放的酚氧化酶活力,当机体受到外界病原刺激下,在一定时间内血细胞释放的酚氧化酶不足以支撑机体的需要,此时血蓝蛋白会表现出酚氧化酶活性参与免疫反应增强甲壳动物免疫防御能力[22]。而本研究中发现TDCH含有一个典型的酪氨酸酶结构域,提示TDCH 可能是对虾血蓝蛋白混合物酚氧化酶活性的一个来源,这还需要进一步实验证实。
利用AlphaFold 预测凡纳滨对虾TDCH 的三维结构,结果显示该蛋白主要以α螺旋结构为主,与已知节肢动物血蓝蛋白的结构组成一致[23]。在对虾中,眼柄与一系列生理功能高度相关,包括卵巢成熟、蜕皮、葡萄糖稳态,这些都与对虾的生长高度相关[24]。Tdch在对虾的眼柄中含量最丰富(图4),提示tdch可能是内分泌调节相关基因之一。此外,血细胞[25]、鳃[26]和肝胰腺[27]是对虾中重要的免疫相关组织。本研究中,这些组织中含有丰富的tdch,提示tdch可能与凡纳滨对虾的先天免疫有关。在副溶血弧菌、金黄色葡萄球菌、黑曲霉菌、白斑综合征病毒、LPS 和病毒模拟物Poly(I:C)的刺激下,tdch在对虾血细胞中的转录水平有显著和持久的升高(图5)。考虑到合成的dsRNA 类似物Poly(I:C)是病毒感染过程中发生的一种分子模式,tdch在Poly(I:C)和白斑综合征病毒刺激过程中上调,表明tdch可能对包括DNA 病毒和RNA 病毒在内的病毒感染做出反应。值得注意的是,在副溶血弧菌、金黄色葡萄球菌和白斑综合征病毒感染后4 h,tdch在血细胞中均显著增加,提示tdch可能在感染早期就对细菌和病毒感染产生了反应。而黑曲霉菌刺激24 h 后tdch在血细胞中的表达才显著增加,提示tdch可能在真菌感染的早期反应不明显,主要在中晚期起作用。tdch在鳃组织中的反应弱于血细胞中,这可能是由于tdch需要在特定的组织环境中起作用。存活率实验表明,敲降tdch能提高副溶血弧菌的敏感性,提示tdch在机体对抗副溶血弧菌感染中发挥重要作用。有研究表明,凡纳滨对虾血蓝蛋白HMC可被副溶血弧菌、金黄色葡萄球菌及白斑综合征病毒激活,促进p38及c-Jun磷酸化,调控下游抗菌肽的表达,参与抗菌与抗病毒感染[28]。然而凡纳滨对虾tdch是通过何种途径发挥抗弧菌作用还有待进一步研究。
4 结论
本研究表明,tdch在各组织中均有表达,在眼柄中表达量最高;副溶血弧菌、金黄色葡萄球菌、黑曲霉菌、白斑综合征病毒、LPS及Poly(I:C)刺激均可引起凡纳滨对虾tdch在血细胞中表达量上调,敲降tdch可提高副溶血弧菌的敏感性,表明tdch在机体对抗副溶血弧菌感染中发挥重要作用。本研究可为甲壳动物血蓝蛋白类群的免疫功能研究提供新思路。
猜你喜欢
当代水产(2022年8期)2022-09-20
食品安全导刊(2021年20期)2021-08-30
昆明医科大学学报(2020年12期)2021-01-26
当代水产(2019年5期)2019-07-25
当代水产(2019年2期)2019-05-16
当代水产(2019年1期)2019-05-16
中成药(2018年10期)2018-10-26
上海农业学报(2017年4期)2017-04-10
中国粮油学报(2016年5期)2016-01-23
广东海洋大学学报(2015年4期)2016-01-13
