
生态恢复对石漠化地区土壤有机碳累积特征及其机制的影响
2022-10-12 13:50龙启霞蓝家程姜勇祥
生态学报 2022年18期
龙启霞,蓝家程,*,姜勇祥
1 贵州师范大学喀斯特研究院, 贵阳 550001 2 国家喀斯特石漠化防治工程技术研究中心, 贵阳 550001 3 福建师范大学地理科学学院, 福州 350007
石漠化是指由于人类活动导致植被破坏、土壤退化,并转为裸露的岩石景观的过程[1],被认为是中国西南喀斯特地区最为严重的生态环境问题之一[2]。为控制西南喀斯特地区水土流失、恢复和改善退化的喀斯特生态系统,国家实施了一系列的退耕还林生态工程治理措施。植被恢复期间土壤生态功能和质量可能会得到改善[3]。土壤有机碳(SOC) 参与了土壤的多种物理化学和生物过程[4],是土壤质量的重要组成部分[5]。同时,全球土壤有机碳库为1500—2300 Pg,约为大气碳库的2倍,其微小变化都会对大气CO2浓度的平衡和调节气候变化有重要作用[6]。土壤有机质参与土壤团聚体的形成和破环,是团聚体稳定的重要因素[7]。土壤团聚体稳定性被认为是反映土壤质量的一个重要指标[8],其团聚过程反过来促进了SOC的物理保护[9],并在维持土壤肥力和质量方面起着至关重要的作用[10]。因此,深入理解喀斯特地区生态恢复下SOC及组分对团聚体及其结合有机碳的影响,可为石漠化地区土壤生态功能恢复、控制水土流失和土地退化提供理论支撑。
植被恢复被认为是增加SOC固存的有效措施[11]。然而,在植被恢复过程中,SOC积累仍存在很大的不确定性。植被恢复可以显著增加[12]、减少[13]或没有显著改变[14]SOC含量。这可能归因于退耕造林以后的植被恢复类型、土壤深度和退耕时间等[15—16],尤其是SOC复杂的化学或物理组分的变化[17]及其与团聚体的相互关系。由耕地到原始森林的植被自然演替,喀斯特地区SOC含量逐渐增加[18],但耕地转变为人工林SOC含量略有下降[19]。因此,深刻认识生态恢复后的土壤有机碳库变化与团聚体关系对制定喀斯特地区合理的生态恢复策略具有重要意义。土壤SOC包含不同稳定性和周转率的组分。土壤易氧化有机碳(Easily oxidizable organic carbon, EOC) 是土壤有机碳库中易被氧化且活性较高的组分[20],SOC与EOC之差可以用来表征非活性有机碳(Non-liable organic carbon, NLOC) 或惰性有机碳。土壤SOC按颗粒分组可分为颗粒有机碳(Particulate organic carbon, POC) 和矿物结合有机碳(Mineral associated organic carbon, MOC),POC是新鲜动植物残体和腐殖化有机碳过渡的碳库,也可以表征土壤活性有机碳[21],MOC则属于惰性有机碳库,在一定程度上可以表征土壤碳库稳定性[22]。SOC和植物残体被认为是团聚体形成的重要胶结剂,显著影响团聚体稳定性[23],反之,团聚体在形成过程中伴随着新鲜有机物的不断输入,对活性碳组分的保护和固定发挥重要的作用[24—25]。一些研究主要聚焦于团聚体稳定性与不稳定和活性碳组分的关系。土壤活性有机碳组分与团聚体稳定性密切相关[26]。Wang等[27]认为SOC的活性组分(EOC) 和生物活性组分与>2 mm团聚体含量和团聚体稳定性呈正相关,同时活性有机碳通过改善土壤团聚体结构来增强团聚体对有机碳的物理保护[28]。POC构成大团聚体的核心,参与团聚体的周转和形成,并受到团聚体的物理保护[24]。然而,稳定有机碳组分与团聚体组成和稳定性的关系较少受到关注。Wu等[10]研究发现较不稳定有机碳组分,化学稳定性有机碳组分对团聚体稳定性的影响更显著,稳定性有机碳促进>2 mm团聚体的形成,>2 mm团聚体的物理保护降低了SOC的损失。可见,土壤有机碳组分对团聚体组成和稳定性的影响存有一些争议。此外,不同粒径团聚体有机碳对SOC贡献大小存在差异[29]。罗晓虹等[30]认为有机碳主要储存于<0.053 mm粒径中,以<0.053 mm粒径对SOC贡献率最大。相反,周方亮等[31]认为>2 mm团聚体有机碳含量最大,Shen等[32]研究表明>2 mm团聚体有机碳对SOC的贡献率最高。然而,已有研究缺乏从土壤碳组分对团聚体及其结合有机碳的影响的角度去解析生态恢复下SOC积累特征及其机制。
中国西南喀斯特地区是全球碳酸盐岩集中分布区面积最大,以石漠化为特征的土地退化严重。区域尺度上的生态恢复工程促进了植被恢复,显著提高了区域植被碳储量,表明中国西南喀斯特地区固碳潜力巨大[33]。已有的研究主要关注喀斯特地区植被恢复/土地利用对SOC及其活性组分、团聚体有机碳的分布特征的影响[18,34—37],一些研究发现,喀斯特地区植被自然恢复可以促进SOC积累[18,38],并促进团聚体形成和提高团聚体的稳定性[39—40]。鲜有将人工恢复和自然恢复进行对比,研究喀斯特石漠化地区生态恢复下土壤SOC组分变化(尤其是POC和MOC) 与团聚体关系的报道。贵州花江小流域为典型喀斯特高原峡谷干热河谷石漠化地区,自1999年以来,该地区开始实施退耕还林工程,使得大量坡耕地退耕为造林地。其中自然退耕的次生林和人工种植的花椒林为该地区典型的主要生态恢复模式。鉴于此,本文主要探究土壤SOC组分与团聚体及其结合有机碳的关系,以期更好地从团聚体尺度去理解喀斯特石漠化地区生态恢复后SOC积累特征及其机制,为石漠化地区植被恢复重建和改善土壤质量提供理论支撑。
1 材料与方法
1.1 研究区概况
研究区域位于中国贵州省安顺市关岭自治县和贞丰县交界的花江干热河谷小流域(25°39′13″—25°41′00″ N, 105°36′—105°46′107 30″ E, 图1)。研究区为亚热带河谷气候,年平均气温18.4℃,年降水量约1100 mm,降雨时空分布不均,集中在5—10月份。花江示范区主要发育在三叠系碳酸盐岩组为主的法郎向斜构造上,地势西高东低,海拔500—1200 m,为典型的喀斯特高原峡谷地貌。区域植被覆盖率低,岩石裸露率高达85%以上,为典型的中—强度石漠化地区。区域母岩以白云质石灰岩和泥质灰岩为主,土壤类型以钙质石灰土为主,土层薄且分布不均。研究区土地利用以林地和耕地为主。林地主要为自然退耕的次生林和人工种植的经济林。其中次生林为坡耕地退耕15—20 a,主要树种为栾树(Koelreuteriapaniculata)、香椿(Toonasinensis)、浆果楝(Cipadessabaccifera)、毛桐(Mallotusbarbatus)、八角枫(Alangiumchinense)、油桐(Verniciafordii)、粗糠柴(Mallotusphilippensis) 等,受人为干扰较少。花椒林是当地兼顾生态与经济效益的主要经济林,为坡耕地退耕后连续种植15—20年的纯林,已成为当地主要造林树种。花椒林的经营管理较为粗放,使用较多的除草剂、农药和复合肥[41],每年定期进行适当修剪管理,以保证花椒有效地采光与通风。耕地主要是种植历史50年以上的玉米,施用化肥、复合肥和农家肥混合,定期进行人为翻土、除草,采用传统耕作方式进行土壤管理。
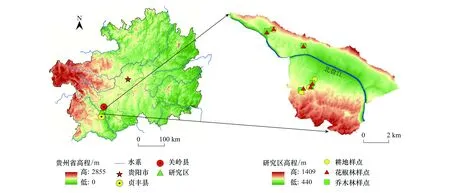
图1 研究区采样地理位置Fig.1 The geographic location of the sample in the study area
1.2 样地设置及采样
2018年11月在示范区内设置了6个样地,每个样地设置包括耕地、花椒林和次生林。每个样地中的次生林和花椒林与耕地相邻。每个样地每种土地利用分别设置3个样方,每个样方按照5点混合采样法,分别采集0—10 cm和10—20 cm土层样品,共108个土样。采用小铲子分土层采集大约2 kg土壤样品,装入密封袋混合成一个土样,带回实验室自然风干测定土壤理化性质。使用100 cm3不锈钢环刀在每层采集约1 kg原状土壤,为保证土壤原状结构,使用塑料盒带回实验室,取出样品沿自然结构面掰成约<10 mm的土壤颗粒,自然风干后去除土样中动植物残茬和砾石,进行颗粒分级及团聚体测定。土壤容重采用环刀法进行取样。
1.3 实验分析与测试
土壤团聚体测定[42]:称取100 g风干土样,置于2 mm、0.25 mm和0.053 mm组成的套筛上,进行2 min的上下均匀震荡(30次/min),残留在不同筛子上的土壤用蒸馏水洗到铝盒中,将铝盒中的土壤置于60℃的烘箱中烘干至恒重,最后获得4个团聚体粒径组分,分别称重并计算其占全土的百分比后,参考SOC的测定方法,将分离后的各粒径土壤团聚体有机碳含量进行测定。
土壤样品研磨过0.25 mm筛,并用稀HCl去除无机碳后,土壤SOC含量采用元素分析仪—稳定同位素质谱仪(Vario ISOPOTE Cube-Isoprime Elementar公司,德国) 测定。POC的测定如下[43]:称取过2 mm风干土样20 g,放入250 mL三角瓶中,加入浓度为5 g/L的六偏磷酸钠60 mL,用手摇约10—15 min后使用往复式震荡器振荡18 h(18℃, 90 rpm/min),震荡后的分散液倒在53 μm筛上,用纯水洗至筛下水为澄清,筛上的土壤颗粒即为颗粒有机质,分离后于铝盒中60℃下过夜烘干(24 h),并进行称重和计算其占全土的百分比。将>53 μm土壤颗粒磨碎过0.149 mm筛,取一定重量样品测定其有机碳含量,乘以其占全土的百分比计算出POC含量(g/kg)。土壤MOC含量为SOC与POC之差。土壤EOC采用333 mM KMnO4测定[44],土壤NLOC为SOC与EOC之差。
1.4 数据计算
土壤有机碳储量计算公式[45]如下:
OCs=BD×D×OC×10-2
式中,OCs为土壤SOC、POC、EOC、MOC、NLOC的储量(kg/m2),BD为土壤容重(g/cm3),D为土层厚度(cm),OC为土壤SOC、POC、EOC、MOC、NLOC的含量(g/kg)。
土壤有机碳组分分配比例计算公式[46]如下:
式中,Fc为土壤有机碳组分分配比例(%);OCi为土壤有机碳i组分含量(g/kg);SOC为土壤有机碳含量(g/kg)。
土壤团聚体有机碳贡献率、MWD和MGD计算公式[36]如下:
式中,Fa为土壤团聚体有机碳贡献率(%);ASOCi为i级粒经团聚体有机碳含量(g/kg);Ai为i级粒经团聚体含量(%);SOC为土壤有机碳含量(g/kg);MWD为平均重量直径(mm);MGD为几何平均直径(mm);xi为i级团聚体平均直径(mm);wi为i粒径团聚体重量百分比(%)。
1.5 统计分析
使用SPSS 26 软件进行统计分析,采用单因素方差分析(One-way ANOVA) 和多重比较LSD法分析不同土地利用土壤碳组分含量和储量、团聚体组成和稳定性、团聚体有机碳含量及有机碳贡献率之间是否显著差异(P<0.05)。采用皮尔逊相关分析团聚体组成、稳定性、团聚体有机碳和土壤碳组分之间的关系(P<0.05)。
2 结果与分析
2.1 土壤碳组分特征
生态恢复后,SOC及其组分含量和储量显著提高(MOC除外)(图2)。SOC及其组分含量和储量的变化模式在两个土层(0—10 cm和10—20 cm) 相似,基本呈现次生林>花椒林>耕地。在0—10 cm土层,花椒林和次生林的土壤SOC、POC、EOC、NLOC含量分别增加了16.6%、68.2%、26.8%、14.7%和56.5%、151.7%、98.2%、48.5%,储量分别增加了12.7%、62.0%、22.2%、11.0%和47.5%、134.5%、86.3%、40.2%。在10—20 cm土层中,土壤碳组分的变化趋势与0—10 cm土层相似,但相对于0—10 cm土层,10—20 cm土层碳组分的含量和储量增加幅度较小。次生林和花椒林的土壤MOC含量与耕地均无显著差异。花椒林和次生林下的POC和EOC等活性有机碳增加幅度大于SOC,而MOC和NLOC等稳定性有机碳增加的幅度小于SOC。
生态恢复增加了活性有机碳组分的比例,减小了非活性或惰性有机碳组分的比例(图2)。在0—10 cm土层,花椒林和次生林的POC分配比(POC/SOC)、EOC分配比(EOC/SOC) 分别显著增加了38.3%、7.8%和58.3%、26.2%,MOC、NLOC分配比(MOC/SOC、NLOC/SOC) 的变化趋势与POC/SOC、EOC/SOC相反,分别下降了15.3%、1.5%和23.3%、5.1%。在10—20 cm土层,EOC/SOC、NLOC/SOC的趋势与0—10 cm土层一致,花椒林的POC/SOC最高,但其MOC/SOC最低。
2.2 土壤团聚体组成及稳定性
在0—10 cm土层,由耕地转为花椒林和次生林显著增加了>2 mm团聚体含量,显著减少了0.25—2 mm、0.053—0.25 mm和<0.053 mm团聚体含量(图3),花椒林和次生林的>2 mm团聚体含量比耕地分别增加了162.2%和379.3%,0.25—2 mm、0.053—0.25 mm和<0.053 mm团聚体含量比耕地分别下降了20.0%、36.7%、51.8%和52.8%、81.9%、83.0%。10—20 cm土层的团聚体分布的变化规律与0—10 cm土层相同,但相较于0—10 cm土层,各团聚体分布的变化幅度相对较小。生态修复后,>2 mm团聚体含量的变化最大,其次为<0.053 mm和0.053—0.25 mm团聚体,0.25—2 mm团聚体的变化最小。
生态恢复显著增加MWD和MGD,表现为次生林>花椒林>耕地(图3)。0—10 cm土层,花椒林和次生林的土壤MWD、MGD分别增加了75.2%、82.6%和172.0%、265.2%。10—20 cm土层,花椒林和次生林的土壤MWD、MGD分别增加了77.2%、94.5%和92.4%、125.6%。
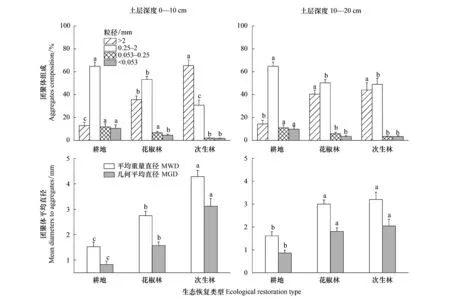
图3 生态恢复后土壤团聚体组成和稳定性的变化Fig.3 Changes of soil aggregates composition and stability after ecological restorationMWD:平均重量直径;MGD:几何平均直径
2.3 团聚体有机碳含量及团聚体有机碳贡献率
在0—20 cm土层,耕地转为花椒林和次生林不同程度增加了>2 mm、0.25—2 mm、0.053—0.25 mm和<0.053 mm团聚体有机碳含量(图4)。在0—10 cm土层,花椒林和次生林的>2 mm、0.25—2 mm、0.053—0.25 mm和<0.053 mm团聚体有机碳含量依次增加了12.9%、12.2%、19.0%、16.9%和51.4%、52.5%、84.2%、62.2%,其中次生林的土壤团聚体有机碳含量显著高于花椒林和耕地。在10—20 cm土层,不同土地利用的各粒径团聚体有机碳含量差异不显著(P>0.05)。生态恢复后,0.053—0.25 mm和<0.053 mm团聚体有机碳含量的增加幅度较大,>2 mm团聚体有机碳含量的增加幅度最小。在0—20 cm土层,生态恢复后各粒径团聚体有机碳贡献率的变化趋势与土壤团聚体组成一致(图4)。生态恢复显著增加了>2 mm团聚体有机碳贡献率,显著减少了0.25—2 mm、0.053—0.25 mm和<0.053 mm团聚体有机碳贡献率。在0—10 cm土层,花椒林和次生林的>2 mm团聚体有机碳贡献率分别增加157.9%和363.7%,0.25—2 mm、0.053—0.25 mm和<0.053 mm团聚体有机碳贡献率分别下降了22.5%、37.3%、51.0%和53.7%、78.7%、82.1%。>2 mm团聚体有机碳贡献率增加的幅度大于<2 mm团聚体有机碳贡献率减小的幅度。耕地中以0.25—2 mm团聚体有机碳贡献为主体,花椒林以0.25—2 mm和>2 mm团聚体有机碳贡献为主,次生林以>2 mm团聚体有机碳贡献为主导。
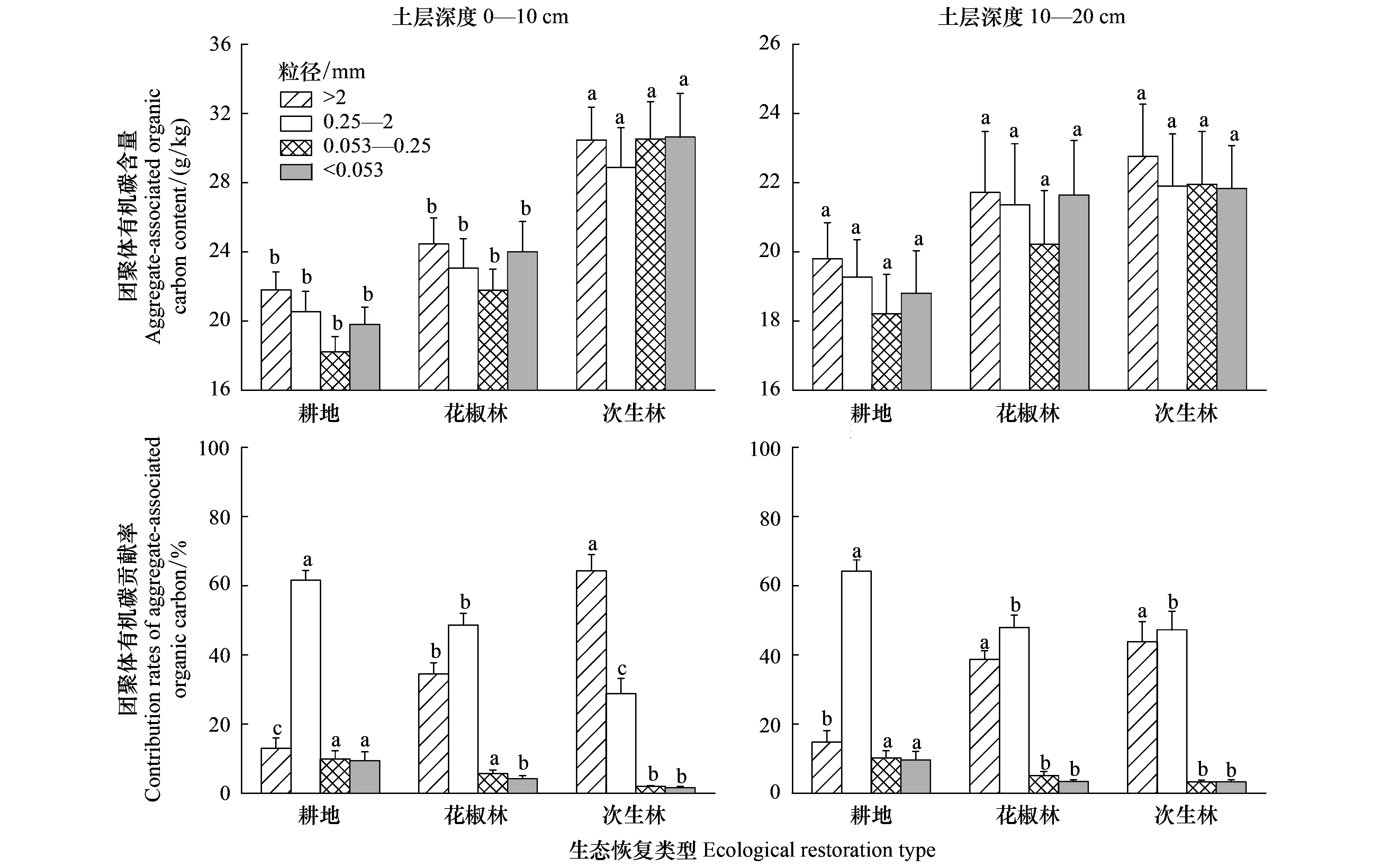
图4 生态恢复后土壤团聚体有机碳含量及贡献率的变化Fig.4 Variation of content and contribution rates of soil aggregate-associated organic carbon after ecological restoration
2.4 土壤碳组分间关系及碳组分与团聚体的关系
由表1可知,土壤SOC分别与POC、EOC、MOC和NLOC显著正相关(P<0.01);与MOC(r=0.515) 相比,POC、EOC与SOC的相关系数较高(r=0.876,r=0.871);除了MOC外,土壤碳组分之间均显著正相关(P<0.01)。
由表2可知,>2 mm团聚体有机碳贡献率与土壤SOC、POC、EOC显著正相关(P<0.01),与土壤NLOC显著正相关(P<0.05),0.25—2 mm和0.053—0.25 mm团聚体有机碳贡献率与土壤SOC、POC、EOC、NLOC显著负相关(P<0.05),< 0.053 mm团聚体有机碳贡献率与土壤碳组分无显著相关(P>0.05);各粒径团聚体有机碳贡献率与土壤MOC均无显著相关(P>0.05)。
由表2可知,除MOC外,土壤SOC、POC、EOC、NLOC均与>2 mm团聚体组成、MWD、MGD显著正相关(P<0.05),与0.25—2 mm、0.053—0.25 mm团聚体组成显著负相关(P<0.05),且POC和EOC对团聚体组成和稳定性的影响更为显著(P<0.01);土壤MOC与团聚体组成和稳定性无显著相关(P>0.05),土壤各碳组分均与<0.053 mm团聚体组成无显著相关(P>0.05)。土壤碳组分与团聚体有机碳含量显著正相关(P<0.01),相对于MOC,土壤POC、EOC与团聚体有机碳含量的相关性较强(r=0.884,P<0.01),土壤NLOC与团聚体有机碳含量相关系数也较高。土壤碳组分与团聚体有机碳贡献率的关系与团聚体组成一致,土壤POC、EOC对团聚体有机碳贡献率的影响更为显著(P<0.01)。
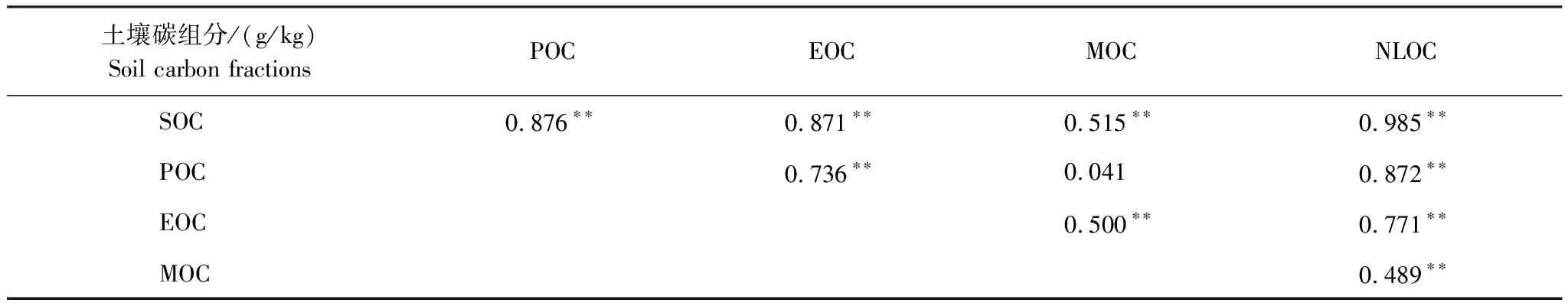
表1 土壤碳组分之间的相关性分析
2.5 土壤团聚体组成、有机碳含量与团聚体有机碳贡献率关系
由表3可知,各粒径团聚体有机碳含量与>2 mm团聚体有机碳贡献率显著正相关(P<0.05),与0.053—0.25 mm团聚体有机碳贡献率显著负相关(P<0.05),与<0.053 mm团聚体有机碳贡献率无显著相关(P>0.05);0.053—0.25 mm和<0.053 mm团聚体有机碳含量与有机碳贡献率相关性较强。土壤各粒径团聚体含量与其有机碳贡献率均显著正相关(P<0.01)。
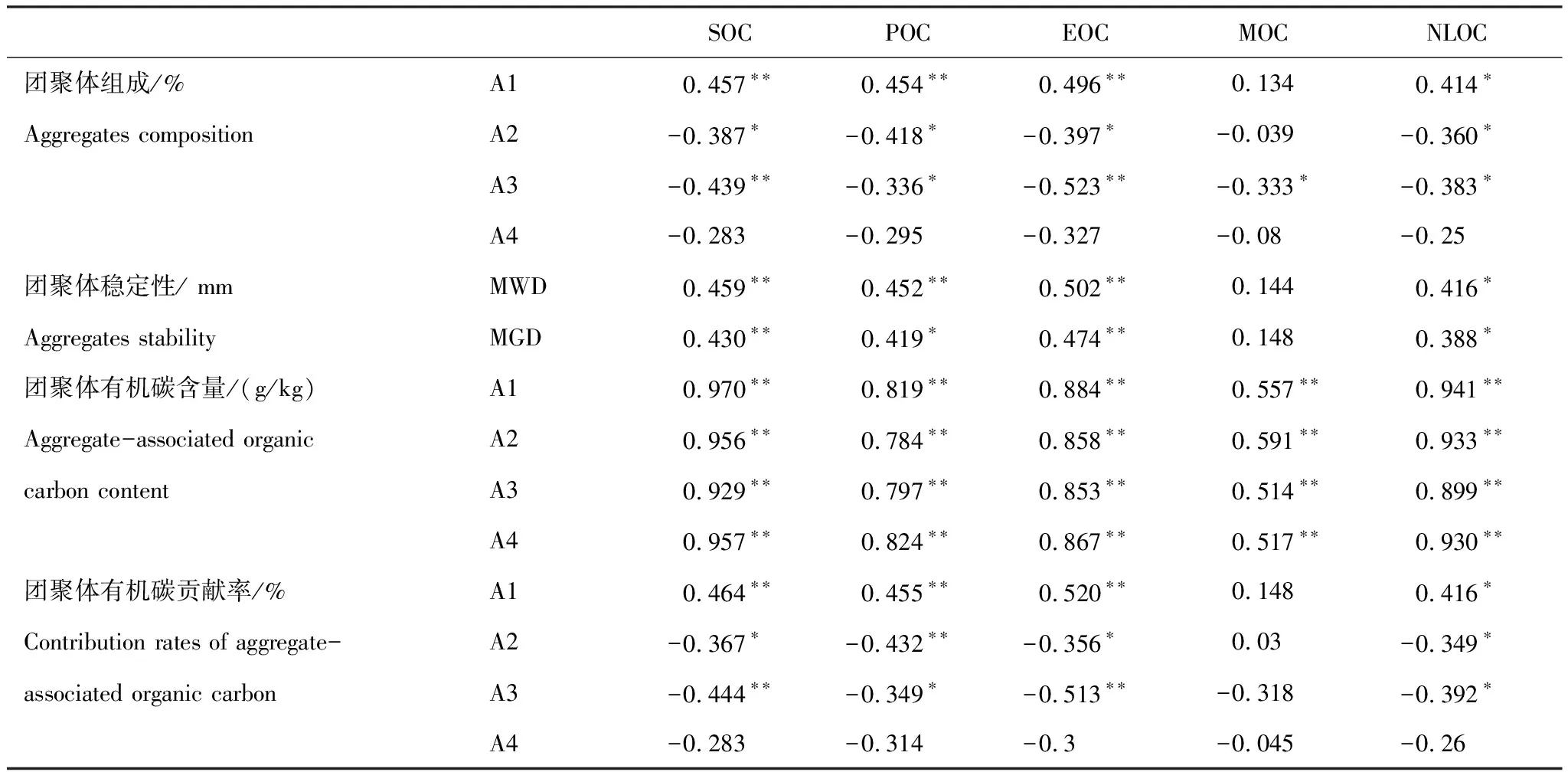
表2 土壤碳组分与团聚体组成、稳定性及团聚体有机碳的相关系数
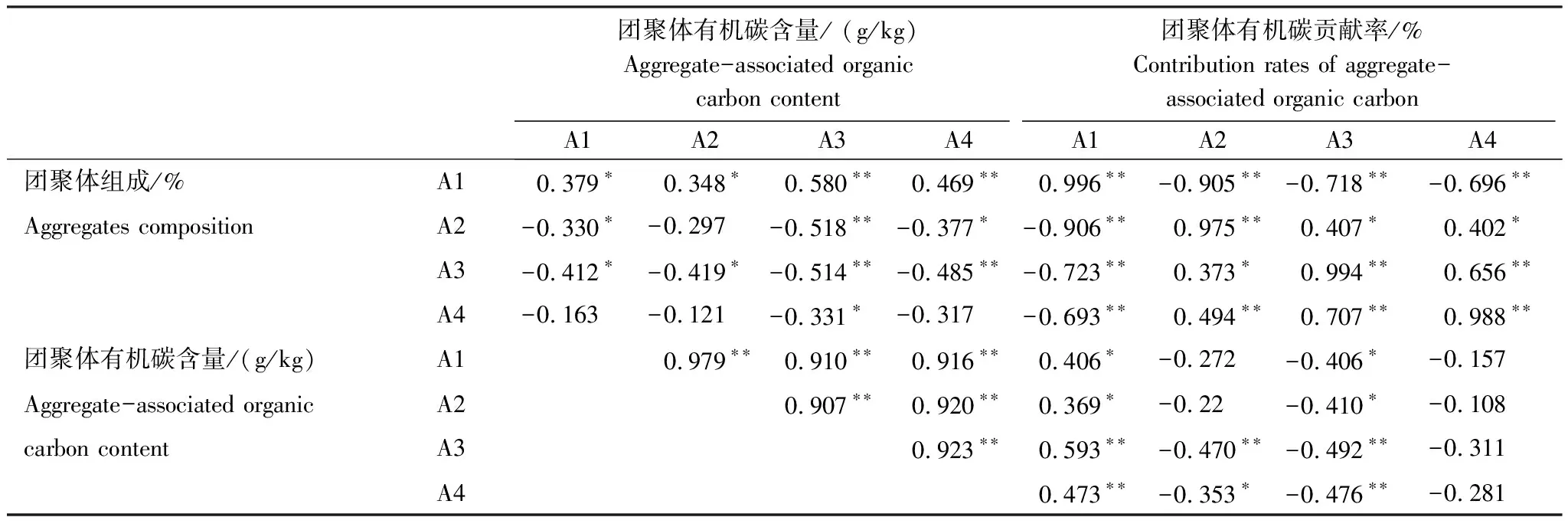
表3 土壤团聚体组成、有机碳含量与贡献率的相关系数
3 讨论
3.1 生态恢复对土壤有机碳及组分影响
土壤SOC储量取决于凋落物、根系分泌物和生物量等碳输入和土壤呼吸、淋失等碳输出的平衡[47]。本研究中生态恢复后,0—20 cm土壤碳组分的含量与储量显著增加(MOC除外)。许多退化喀斯特生态系统的研究也证实了植被恢复促进了SOC的积累是由于退耕后较高的植物残体和根系生物量返回到土壤中[18,38,48]。地表凋落物量和根系分泌物量为土壤有机质积累提供丰富的来源[8]。在同一研究区发现,不同种植年限(5年、10年和20年) 花椒林SOC含量高于耕地[49]。次生林和花椒林的POC、EOC含量和储量、POC/SOC、EOC/SOC显著高于耕地(图2) 表明次生林和花椒林有更多的新鲜植物残体来源碳回归土壤。这是因为土壤活性有机碳主要来源于植物生物量和根系分泌物[43]。次生林地表生物量(2278.32 g/cm2) 大于花椒林地表生物量(681.39 g/cm2) 使得次生林SOC及组分含量较高。杨龙[50]在本研究区的研究也发现花椒林、灌木林和乔木林的生物量为721.39 g/cm2、1219.38 g/cm2和2716.83 g/cm2,SOC含量高低由地表生物量决定。相关分析(表1) 发现SOC与活性有机碳相关性较强(r=0.876,P<0.01),表明土壤SOC的增加主要依赖于土壤POC、EOC等活性有机碳的增加。另外,喀斯特地区植被恢复促使土壤有机质的积累进而促进了土壤团聚体的形成和稳定,从而增强团聚体内有机碳的物理保护[15,39,48]。植被恢复增加的植物残体的输入、根系生物量和分泌物也能够促进土壤颗粒的团聚[51]。相反,长期的耕作或人为扰动一方面破坏了土壤大团聚体,降低了团聚体的稳定性[52],另一方面较高的分解速率、土壤侵蚀和有机质输入减少导致SOC及组分含量和储量[12]。与花椒林和次生林相比,耕地土壤中EOC、POC的含量较低(图2) 表明耕地SOC可能具有较高的分解速率,使得活性有机碳大量损失。大量研究表明植被恢复或造林能够显著增加土壤活性有机碳组分[35,53—54],这与我们的研究结果一致。一方面植被下层凋落物输入可以直接提供活性有机碳组分[55];另一方面,植被恢复提升了微生物活性,微生物迅速分解促进了植物凋落物来源碳转化为活性有机碳组分[56]。然而,生态恢复并未显著提升MOC含量和储量(图2),这与其它喀斯特地区的研究结果一致[15]。这可能是因为MOC是与粘粒和矿物质结合的有机碳组分,相对稳定且很难分解,对土地利用变化不敏感[43]。也可能与恢复时间有关,因为只有长期的植被恢复才能使有机物的输入和输出之间达到相对平衡,非保护性的POC才能逐步向稳定性MOC转化,增加MOC含量[57]。次生林0—10 cm土层土壤NLOC含量和储量显著高于耕地(图2)。总之,生态恢复不仅增加了土壤总有机碳含量,产生了更多的活性有机碳组分,而且也增加了非活性碳组分的含量,但SOC的增加主要归因于土壤活性有机碳的增加。此外,自然恢复措施(次生林) 比人工恢复(种植花椒) 可以更好地促进0—20 cm土层SOC的积累。
3.2 生态恢复下团聚体对土壤有机碳变化的作用
生态恢复后,各粒径团聚体有机碳含量均不同程度增加,其中0.053—0.25 mm和<0.053 mm团聚体有机碳含量的增加幅度较大,>2 mm团聚体有机碳含量的增加幅度最小(图4)。这符合有机碳输入优先向小粒径团聚体积累而后再胶结组成较大团聚体的层级理论[26],并与其它喀斯特地区研究结果一致[37]。从团聚体对有机碳贡献率的变化来看,也证实了较大团聚体是由较小团聚体与有机胶结物组成的。耕地土壤以0.25—2 mm团聚体有机碳贡献为主体,而生态恢复后转为以>2 mm团聚体对有机碳的贡献为主体(图4),>2 mm团聚体有机碳贡献率的显著增加抵消了<2 mm团聚体有机碳贡献率的减小,最终增加了SOC含量。这表明>2 mm团聚体有机碳增加成为生态恢复后SOC增加的主要形式。土壤SOC与>2 mm团聚体含量显著正相关(表2) 也验证了这一点。在其它喀斯特地区的研究也发现植被自然演替恢复过程中SOC的增加主要归因于>2 mm团聚体有机碳的积累[48]。退耕地植被恢复过程中土壤团聚体有机碳贡献率主要受团聚体数量和团聚体有机碳含量的影响[58]。王兴等[29]的研究也发现撂荒地植被恢复过程中团聚体对SOC的贡献率80%来自>2 mm团聚体,>2 mm团聚体数量的显著增加是该团聚体有机碳贡献率高的主要原因,其次才是该团聚体有机碳含量的增加。本研究中,>2 mm团聚体数量、>2 mm团聚体有机碳含量分别与>2 mm团聚体有机碳贡献率显著正相关(表3),且团聚体数量与团聚体有机碳贡献率关系更强(r=0.996,P<0.01),进一步证实了>2 mm团聚体数量是影响团聚体有机碳贡献率的关键因子。此外,0.25—2 mm、0.053—0.25 mm和<0.053 mm团聚体有机碳贡献率显著减小也主要是因为团聚体数量的显著减小(图4,表3),这潜在说明生态恢复使团聚体有机碳含量发生了再分配过程,即生态恢复促进有机碳由<2 mm团聚体向>2 mm团聚体转化,且这种转化主要是由于植被恢复促进了较小团聚体胶结形成较大团聚体,从而保护了有机碳不易被微生物利用。
3.3 生态恢复下土壤碳组分对团聚体的影响
生态恢复后,>2 mm团聚体数量、MWD、MGD显著增加,而0.25—2 mm、0.053—0.25 mm和< 0.053 mm团聚体数量显著减少(P<0.01,图3),这表明生态恢复能够有效改善土壤团聚体结构和团聚体稳定性。这与Hu等[48]在重庆南川岩溶石漠化地区关于植被自然演替恢复对团聚体结构的影响的结果一致。生态恢复后植物残体和根系生物量不断输入土壤,促进SOC的积累,有机碳作为团聚体主要的胶结物质促进团聚体的形成和稳定[59]。SOC及组分与>2 mm团聚体数量、MWD、MGD显著正相关(表2) 验证了这一点。土壤团聚体的有机胶结物质可分为瞬变性(微生物和植物来源得多糖类物质)、临时性(根系和真菌菌丝及分泌的多糖) 和持久性胶结剂三大类[26]。有机碳组分中EOC和POC对团聚体形成和稳定性影响更大,主要是因为EOC主要来源于植物凋落物和根系分泌物的分解[43],主要包含复合多糖等有机化合物[60],POC主要来源于新鲜动植物残体以及腐殖有机质中的临时或过渡有机碳[61],因此EOC和POC很可能由各种临时或暂时的胶结剂组成,它们促进了大团聚体的形成和稳定。廖林超等[62]的研究也验证了土壤团聚体的分布和稳定性与活性有机碳关系更密切,活性有机碳是土壤团聚化的重要有机质胶结物。然而,SOC及组分含量与0.25—2 mm、0.053—0.25 mm和<0.053 mm团聚体数量显著负相关,可能是由于初始有机碳最先在微团聚体或细颗粒中积累,随后整合进入大团聚体[26]。NLOC和MOC含量与团聚体数量和稳定性的相关性较弱(表2),可能与其对生态恢复的响应较不敏感有关。MOC是有机物的最终分解产物,与粘粒、粉粒和矿物质结合的有机碳,相对稳定且很难分解[43],因此其可能很少参与团聚体的形成。然而,Wu等[10]研究发现较不稳定有机碳组分,化学稳定性有机碳组分促进>2 mm团聚体的形成,对团聚体稳定性的影响更显著。NLOC作为非活性有机碳,也与显著团聚体数量和稳定性显著相关,但其相关性小于活性有机碳与团聚体数量和稳定性的关系(表2)。因此,生态恢复使得新鲜有机质不断输入,显著提升了活性有机碳,活性有机碳作为临时或暂时的有机胶结剂主导了>2 mm团聚体形成和稳定。生态恢复增加的有机碳及组分作为胶结物质促进了团聚体周转、形成和稳定,反过来有机碳及组分受到团聚体的物理保护,使得SOC在不同粒径团聚体进行再分配。因此,SOC及组分含量与团聚体有机碳含量显著相关(表2)。生态恢复导致的地上生物量增加了有机残体的输入,可能通过促进大团聚体形成中作为主导胶结剂的EOC和POC的供应,加速大团聚体形成,从而导致大团聚体有机碳储量的显著增加。>2 mm团聚体有机碳贡献率与土壤EOC和POC呈显著正相关,且其相关系数高与NLOC和MOC验证了这一点。
4 结论
(1)生态恢复显著增加土壤有机碳组分含量与储量(MOC除外),尤其是产生了更多的POC和EOC,土壤SOC含量的增加主要依赖于土壤POC、EOC含量的增加。
(2)生态恢复加速了有机碳从较小团聚体向较大团聚体的重新分配过程,>2 mm团聚体有机碳储量增加是生态恢复后SOC储量增加的主要形式,而>2 mm团聚体有机碳储量增加依赖于其团聚体数量的大幅提升,其次才是团聚体有机碳含量的增加。
(3)生态恢复导致更多的新鲜植物残体来源碳的输入,可能通过促进>2 mm团聚体形成中作为主导胶结剂的EOC和POC的供应,从而改善土壤团聚体结构和促进土壤SOC的积累。
猜你喜欢
建材发展导向(2022年10期)2022-07-28
纺织标准与质量(2022年2期)2022-07-12
作物学报(2022年5期)2022-03-16
当代陕西(2020年24期)2020-02-01
当代陕西(2020年24期)2020-02-01
当代陕西(2020年24期)2020-02-01
当代化工(2019年3期)2019-12-12
领导文萃(2017年11期)2017-06-12
高教探索(2015年10期)2015-10-29
哈尔滨理工大学学报(2014年1期)2014-06-23
