
尿酸对猪精液液态保存效果的影响
2022-11-11 12:53林敬轶金小虎石学颖张天羽李丹彤岳顺利周佳勃
中国兽医学报 2022年9期
林敬轶,刘 雪,金小虎,石学颖,张天羽,李丹彤,岳顺利,周佳勃
(东北农业大学 生命科学学院 黑龙江省动物细胞与遗传工程重点实验室,黑龙江 哈尔滨 150030)
近20年全球猪肉消费呈现几何式增长,迫切要求猪生产企业不断提高生产效率来增加猪肉产量和提高猪肉品质[1]。人工授精是提高猪群质量和降低生产成本的关键技术。由于猪冷冻精液应用效果不佳,所以生产中主要使用液态精液进行人工授精[2]。虽然人们对液态保存技术已进行过大量研究,但是仍存在保存时间短和受精能力低等问题。有研究表明,猪精液在液态保存过程中,会随着保存天数的延长,精子内的活性氧(ROS)会逐渐积累。过量的ROS会对精子的结构及功能造成氧化损伤,进而影响其精液的品质和受精能力[3-4]。因此,人们一直在探索和研究用于猪精液液态保存的抗氧化剂[5-6]。
尿酸(UA)是嘌呤代谢的终产物,也是人类精液中的主要抗氧化物质。有研究报道,猪血清UA对烷氧基自由基的清除能力与人血清相同[7]。其在精液中的抗氧化活性接近维生素C的一半[8]。UA可以有效清除超氧阴离子自由基(·O2-)和羟自由基(·OH)等强氧化物,防止细胞受到氧化损伤[9]。有研究证明,人血液中UA含量与精子活力及形态呈正相关的趋势。低尿酸血症患者的精子畸形和精子不动症高发[10]。此外,UA还是鱼精液中主要的抗氧化剂。在冷冻保存过程中添加一定浓度的UA可显著提高鱼精子的活力[11]。
虽然有报道UA对人和一些动物精子有重要的抗氧化作用[11-13],但UA在猪精液保存中的作用尚未见报道。本研究旨在精液保存液中添加UA,探究不同质量浓度的UA对常温保存下猪精子活力、质膜完整性以及顶体完整性的影响。同时,还检测了UA对猪精子抗氧化能力(总抗氧化能力(T-AOC)、ROS含量和脂质过氧化水平)、线粒体膜电位以及体外受精能力的影响。为探究UA延长猪精液液态保存时间、UA的生物学功能和开发新型精液稀释液提供理论依据。
1 材料与方法
1.1 主要试剂除特殊说明外,本研究所用试剂均购自Sigma-Aldrich公司。Modena稀释液作为其猪精子保存液(pH6.9),渗透压为282 mOsm,成分包含25 g/LD-葡萄糖,2.25 g/L乙二胺四乙酸二钠,6.9 g/L柠檬酸钠,2 g/L柠檬酸,5.65 g/L三羟甲基氨基甲烷,0.05 g/L半胱氨酸,1 g/L碳酸氢钠,3 g/L牛血清白蛋白,0.05 g/L青霉素钠,0.05 g/L硫酸链霉素。
1.2 精液采集及保存猪精液采自6头健康可育、年龄均为1.5~2.5岁的长白公猪。通过手握法进行采精,每次采精至少间隔3 d。采集到的鲜精温度需保持在35~37℃之间,且在45 min内运至实验室。其鲜精精子活力经镜检在70%以上,且畸形率在15%以下可用于试验。采集的新鲜精液置于室内避光处理2 h,使其温度缓慢降至室温。将鲜精置于15 mL离心管中,于室温下1 000 r/min离心去除精浆,随后用等温的含有UA的Modena液稀释,稀释后UA的终质量浓度为2.5,5.0,10.0,20.0 mg/L(其对照组不含UA,精子密度为3×107~5×107个/mL)。用1.8 mL冻存管分装精液并置于17℃恒温箱中避光保存,分别检测0,1,3,5 d的精子质量参数[14]。
1.3 精子活力检测使用清华同方精子分析仪(MX 7.5)分析精子活力。检测前取500 μL精液样本混匀,于37℃恒温箱中孵育20 min,取10 μL精子样本于37℃预热的精子计数池上,每次检测6个视野,精子数量不少于200个。
1.4 精子质膜完整性检测采用低渗肿胀法对其精子质膜完整性进行检测[15]。检测前取50 μL样本,加入到500 μL预热低渗液中,37℃孵育50 min。取10 μL混合液于载玻片上,盖片。在光镜显微镜(×400)下观察并统计头部肿胀和弯尾精子数以及总精子数,每次计数不少于200个精子。
1.5 精子顶体完整性检测采用异硫氰酸花生凝集素(FITC-PNA)对顶体进行特异性染色[16]。检测时,取30 μL精液均匀涂抹在载玻片上自然晾干,用无水甲醇固定10 min。随后用30 μL FITC-PNA染液均匀涂片,于37℃避光孵育30 min。用PBS缓冲液轻轻冲洗载玻片2次,并置于黑暗中自然晾干。在倒置荧光显微镜(×400)下观察精子顶体完整性,每次镜检精子数不少于200个精子,且全程避光。
1.6 精子T-AOC检测采用T-AOC测定试剂盒对精子样本的T-AOC进行检测。检测前,取1 mL精子浓度为1×107个/mL的待测样本进行超声破碎处理,4℃、3 000 r/min离心10 min,吸取上清液待用。根据试剂盒说明书加入相应试剂。使用分光光度计在波长为520 nm处测定各管的D值。
1.7 精子ROS含量检测采用2’,7’-二氯荧光素二乙酸酯(2’,7’-dichlorodihydrofluorescein diacetate,DCFH-DA)染色检测精子中ROS含量。检测时,分别取对照组和试验组中1 mL的样本(精子浓度为1×106个/mL),室温下2 000 r/min离心5 min,并用37℃预热PBS缓冲液重悬精子样本。加入DCFH-DA(终浓度为10 μmol/L),于37℃培养箱中避光孵育25 min,期间每隔5 min轻轻颠倒混匀。随后用37℃预热PBS缓冲液洗涤3次,以充分去除未进入细胞内的DCFH-DA。通过流式细胞仪对其进行检测分析,检测细胞数为10 000个,设置其激发波长为488 nm,而发射波长为525 nm。
1.8 精子脂质过氧化检测过氧化脂质降解产物中的丙二醛(MDA)可与硫代巴比妥酸(TBA)缩合形成红色产物,在532 nm处有最大吸收峰。因此,可以通过检测精子MDA含量进而评估其脂质过氧化水平。检测前,取适量精子样本进行超声破碎处理,4℃、3 000 r/min离心10 min,并吸取其上清液用于试验。随后根据试剂盒说明书加入相应试剂,使用分光光度计在波长为532 nm处测定其D值。
1.9 精子线粒体膜电位检测采用线粒体膜电位检测试剂盒对其精子进行检测。取1 mL精液样本,室温环境下2 000 r/min离心5 min。弃上清,用500 μL 37℃预热PBS缓冲液重悬精子。取500 μL JC-1染色工作液与精液混合,反复颠倒并置于37℃恒温箱中避光孵育25 min。4℃、600×g离心5 min,弃上清。用JC-1染色缓冲液(×1)清洗2次。在30 min内用流式细胞仪进行检测分析,计算其线粒体膜电位高的精子百分比。
1.10 卵母细胞体外成熟将收集的卵巢置于37℃ 0.9%生理盐水中尽快运回实验室。随后用含双抗的37℃生理盐水对卵巢缓慢清洗3次,抽取直径为4~6 mm卵泡中的卵丘-卵母细胞复合体用于试验,置于预先37℃平衡4 h成熟液的培养孔中培养,37℃、5% CO2培养箱中培养44 h。
1.11 体外受精检测吸取1 mL精子,在室温环境下,1 000 r/min离心5 min,并用PBS缓冲液洗涤2次后,将精子以106个/mL的密度在mTBM获能液中,37℃、5% CO2培养箱内获能1 h。每50 μL受精滴中放有30个成熟卵母细胞,并加入适量获能精子,使其精子终质量浓度为2.5×105个/mL。在受精6 h后,取部分卵母细胞于胚胎培养液(PZM-3)中培养12 h,用乙酸/乙醇(1∶3)将其固定24 h,经DPBS洗涤3次后,用Hoechst 33342染料染色10 min,在荧光显微镜镜下观察其受精情况(单精受精比例)。其余卵母细胞在受精6 h后,用预先平衡好的PZM-3洗涤3次,随后转入PZM-3滴中,于37℃、5% CO2培养箱中继续培养。48 h后检测并计算卵裂率,6 d后检测并计算囊胚率。

2 结果
2.1 UA对液态保存中猪精子活力的影响在保存第3天时,2.5,5.0,10.0 mg/L UA组的精子活力分别为(74.42±0.16)%,(78.69±0.73)%,(74.48±0.49)%,其活力均高于其对照组(70.73±0.48)%,且差异显著(P<0.05)。而20.0 mg/L UA组的精子活力则显著低于对照组(P<0.05)。保存第5天时,5.0 mg/L UA组的精子活力仍显著高于其他组,比对照组提高了14.61%(表1)。

表1 不同质量浓度的UA对液态保存中猪精子活力的影响 %
2.2 UA对精子质膜完整性的影响如表2所示,在保存第1天时,各组之间无显著性差异(P>0.05)。从保存第3天开始,UA组与对照组之间出现差异。其中,5.0 mg/L UA组的精子质膜完整性(68.45±0.29)%要高于其他组质膜完整性。在保存第5天时,5.0 mg/L UA组的精子质膜完整性(54.71±0.55)%要显著高于对照组(47.23±0.31)%(P<0.05)。
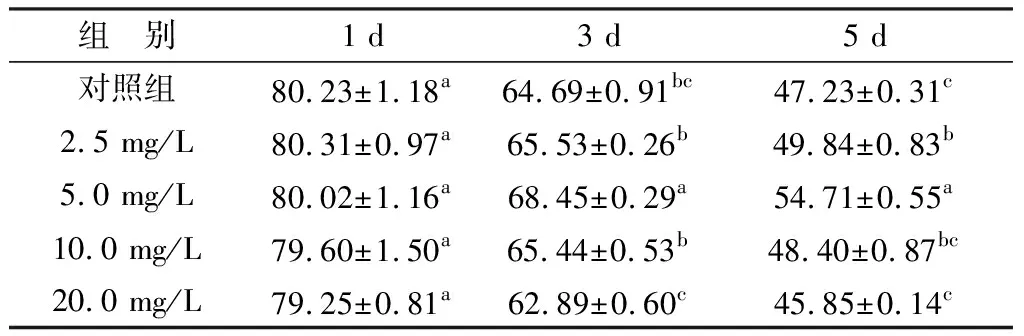
表2 UA对液态保存中猪精子质膜完整性的影响 %
2.3 UA对精子顶体完整性的影响如表3所示,在保存第1天时,各组精子的顶体完整性并无显著性差异(P>0.05)。在保存第3天时,5.0 mg/L UA组要显著高于对照组(P<0.05)。在保存第5天时,2.5,5.0 mg/L UA组均显著高于对照组(P<0.05),且5.0 mg/L UA组的效果最佳,而20.0 mg/L UA组的顶体完整性要显著低于其他各组(P<0.05)。结果表明,在Modena保存液中添加5.0 mg/L的UA对猪精液液态保存效果最佳。因此,后续试验均采用5.0 mg/L UA试验。
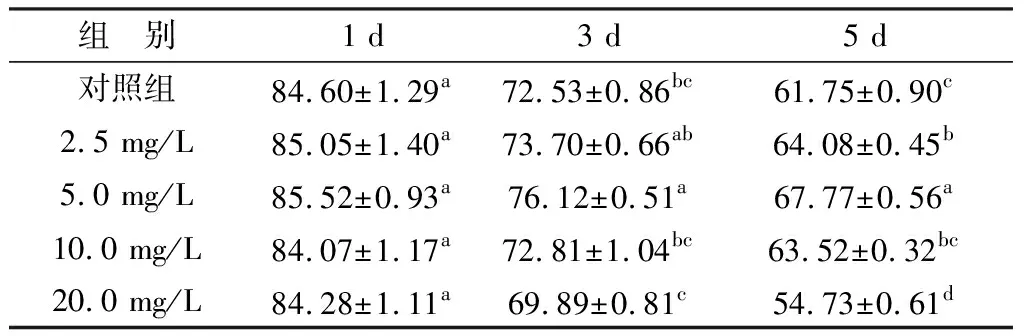
表3 UA对液态保存中猪精子顶体完整性的影响 %
2.4 UA对精子抗氧化能力的影响为探究UA对常温保存过程中精子的抗氧化能力的影响,对其精子的T-AOC、ROS、MDA含量进行检测。结果如表4所示,在保存第1天时,加入5.0 mg/L UA对其精子的抗氧化能力的影响并不显著(P>0.05)。随着保存时间的延长,各组的T-AOC呈下降趋势,而ROS和MDA含量呈升高趋势。但在保存第3天后,对照组与试验组之间的抗氧化能力开始出现显著性差异。其UA组的ROS和MDA含量要显著低于对照组(P<0.05),而其T-AOC要显著高于对照组(P<0.05)。
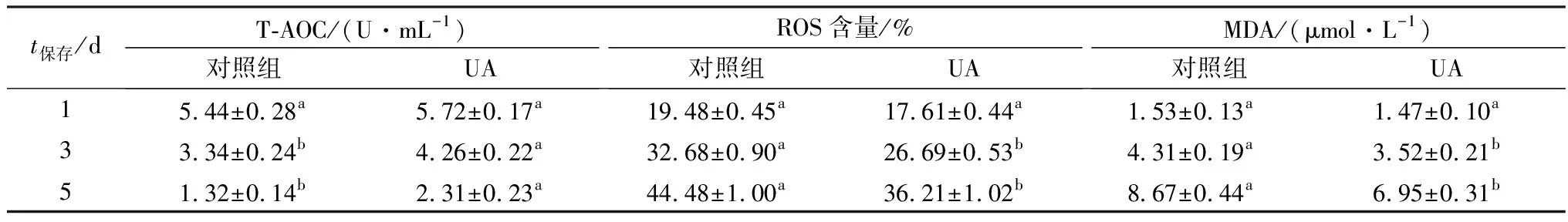
表4 UA对液态保存中猪精子抗氧化能力的影响
2.5 UA对精子线粒体膜电位的影响随着常温保存时间的延长,其线粒体膜电位高的精子比例呈逐渐下降趋势。如表5所示,在第1天时,对照组与5.0 mg/L UA组之间差异不显著(P>0.05)。从第3天开始,两者之间出现差异,UA组(68.55±0.60)%要显著高于对照组(63.41±0.56)%。在第5天时,两者差异扩大,UA组的高膜电位精子比例要高于对照组约6.06%。
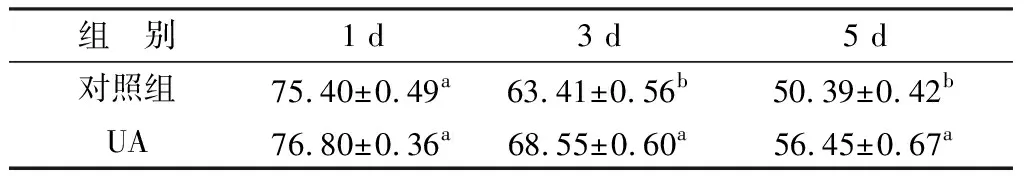
表5 UA对液态保存中猪精子线粒体膜电位的影响
2.6 UA对猪精子体外受精能力的影响根据上述试验结果,选用液态保存第5天的精子样品进行体外受精,检测其体外受精能力及胚胎发育率。如表6所示,5.0 mg/L UA组的穿透率、受精率、卵裂率和囊胚率均显著高于对照组(P<0.05)。

表6 UA对液态保存中猪精子体外受精及胚胎发育的影响
3 讨论
随着液态保存时间的延长,猪精液的质量会逐渐降低,具体表现为猪精子的活力及运动能力降低,质膜与顶体损伤,进而导致其受精能力下降。本研究结果显示,在稀释液中添加一定质量浓度的UA可有效抑制精子活力、质膜及顶体完整性的下降。而其中添加5.0 mg/L的UA对其精子的活力和质膜及顶体完整性要高于其他组。LAHNSTEINER等[17]研究表明,一定浓度的UA可以显著提高其鱼精子活力和精子膜完整性,与本研究结果相一致。此外,有研究表明,适量的UA可提高细胞抗氧化能力并通过调控B淋巴细胞瘤-2(Bcl-2)与Bcl-2相关X蛋白(Bax)的蛋白表达水平比值和半胱氨酸天冬氨酸蛋白酶-3(Caspase-3)蛋白表达水平进而减少其细胞凋亡[18],说明UA可能会抑制保存过程中精子的凋亡,延长精子寿命。
本研究还发现,高质量浓度的UA(20.0 mg/L)会显著降低其精子质量。这与ZHANG等[19]报道的结果类似,较高质量浓度的UA会导致细胞活率显著降低。有研究表明,高质量浓度UA会导致男性精液质量显著降低,其机制可能是高质量浓度UA通过减弱内皮一氧化氮合酶活性,导致一氧化氮水平减少,进而影响其精子活力。也可能是高质量浓度UA通过抑制肌酸激酶的活性,减少ATP的产生,从而降低精子功能[13,20-21]。
众所周知,细胞内ROS的形成主要是由细胞线粒体所产生的。在体外保存过程中,精子会由于自身代谢而产生ROS。生理范围的ROS在精子获能、过度活化、顶体反应、受精能力的维持和精子-卵母细胞相互作用中起着至关重要的作用[22]。但大量的ROS堆积会引起精子质膜脂质过氧化,使其通透性发生改变,严重时发生破损,进而导致精子结构完整性被破坏和运动性能降低。而猪精子中多不饱和脂肪酸的浓度高于其他物种,因此猪精子更容易受到氧化损伤[23],发生脂质过氧化。而MDA则是氧化应激脂质过氧化过程中最普遍的脂质过氧化副产物,常作为氧化应激的生物标志物[24]。本研究结果表明,添加5.0 mg/L UA能够有效降低其ROS水平,改善了精子的脂质过氧化水平和T-AOC。YA等[25]的研究结果表明,适宜浓度的UA可有效减轻氧化损伤,降低MDA含量。也有大量报道显示,一定质量浓度的UA可有效提高其细胞的抗氧化能力,改善脂质过氧化水平,降低ROS水平,进而提高细胞的存活率[17,19,26-27]。因此,我们认为UA通过这种提高其抗氧化能力以及减少脂质过氧化水平,进而改善在液态保存过程中维持其质膜完整性,有利于精子活力的维持,延长保存时间。
线粒体是精子氧化磷酸化形成ATP的主要场所,在为精子运动提供ATP方面发挥着重要作用[28]。当受到高浓度的ROS影响时,氧化应激会导致线粒体功能受到抑制,线粒体膜及DNA受到损伤,使线粒体膜发生去极化,从而导致线粒体活性、线粒体膜电位和精子活力的下降[29-32]。而低膜电位的动物精子不易发生顶体反应[33]。通过完整的线粒体膜电位来实现的线粒体功能似乎是维持精子中顶体酶活性、顶体反应和染色质完整性的先决条件。因此,线粒体膜电位是评价线粒体功能及精子质量的重要指标之一[34]。本研究结果表明,在保存过程中,添加UA可提高猪精子的线粒体膜电位。刘宣等[35]报道,UA能有效清除ROS,提高大鼠肾上腺嗜铬细胞瘤PC12细胞线粒体膜电位,与本研究结果完全一致,其机制可能是UA通过清除过量ROS来维持线粒体的正常功能。
为了进一步评价UA是否能够提高保存精子的受精能力和胚胎发育能力,本试验通过体外受精检测其液态保存精子的穿透率、受精率、卵裂率和囊胚率,结果表明,与对照组相比,在保存液中加入5.0 mg/L UA能够有效提高体外受精的精子穿透率、受精率、卵裂率及囊胚率。精子质膜和顶体完整是精子存活和完成受精的重要条件。结合UA可提高精子质量等结果,我们推测UA通过提高精子活力、质膜和顶体完整性,从而提高其精子穿透率及体外受精能力。精子因含有高含量多不饱和脂肪酸,在受到氧化应激损伤时,脂质过氧化的副产物会导致DNA损伤,诱导DNA片段化,进而致使胚胎发育停滞[36-37]。有研究表明,UA能有效保护DNA免受损伤[38]。此外,HUGHES等[39]的研究结果显示UA能有效减轻人精子在X-射线照射下所诱导的DNA损伤程度。因此,我们推测UA可能是通过清除ROS,减少精子DNA损伤,进而提高胚胎发育能力。
综上所述,在Modena保存液中添加一定质量浓度的UA,可显著提高精子的活力、质膜完整性和顶体完整性。UA能够有效清除活性氧,显著提高其精子抗氧化能力,维持精子的线粒体活性,从而延长猪精子体外保存时间。此外,UA还可以有效提高保存精子的受精能力和胚胎发育能力。目前,UA对猪精子的影响仍处于初步研究阶段,还可能存在其他调控机制需要深入研究。相关研究结果可为探究UA对雄性动物生殖细胞的影响及机制研究和新型猪精液稀释液的研发提供理论参考。
猜你喜欢
重庆理工大学学报(自然科学)(2022年9期)2022-10-26
中华实用诊断与治疗杂志(2022年1期)2022-08-31
土壤学报(2022年3期)2022-08-26
国外畜牧学·猪与禽(2022年1期)2022-04-23
焊接学报(2022年1期)2022-03-28
文萃报·周二版(2020年42期)2020-11-28
中国医药导报(2019年13期)2019-06-20
文理导航(2016年32期)2016-12-19
飞碟探索(2016年4期)2016-04-07
科学启蒙(2013年7期)2013-09-22
