
多年生黑麦草、雀麦根系形态和生长对土壤干旱的适应性*
2022-11-16 08:34张咏梅胡海英白小明MatthewCoryGarcFavreJavierOrdezIv
中国生态农业学报(中英文) 2022年11期
张咏梅,胡海英,白小明,Matthew Cory,García-Favre Javier,Ordóñez Iván P,5
(1.甘肃农业大学干旱生境作物学国家重点实验室 兰州 730070;2.宁夏大学农学院 银川 750021;3.甘肃农业大学草业学院 兰州 730070;4.梅西大学农业与环境学院 北帕 4442 新西兰;5.坎佩奈克农艺研究所 蓬塔 6212707 智利)
根系是连接植物地上部分茎叶与土壤进行物质循环和能量流动的重要桥梁与通道[1-2]。根系从土壤中汲取水分和养分,供给茎叶进行光合作用,茎叶光合作用产生的能量和光合产物又有部分运输至根系,满足根系生长的需要。根系作为植物汲取水分和养分的主要器官,也是对土壤水分变化最敏感的器官,不仅决定着植物对土壤资源的利用方式和利用效率,也反映了植物对环境的适应策略和生存策略[3]。研究表明,根系在适应干旱胁迫方面发挥重要的作用[4]。生长在砂壤土的小麦(Triticum aestivum)在干旱条件下,根系表面可以形成根鞘,有助于小麦从干燥的土壤中获取水分和养分[5]。对9 种禾本科草本植物进行耐旱能力评价,偃麦草(Elymus repens)‘京草1 号’和柳枝稷(Panicum virgatum)在干旱胁迫下,细根根长和根表面积呈增加趋势,表现出增大了根系与土壤的接触面积以获取水分的适应性策略[6]。在干旱条件(年降雨量150 mm)下,内蒙古锡林郭勒典型草原建群种羊草(Leymus chinensis)种群采用地上个体小型化,地下主根变细,分叉,向土壤深层扩展的生长策略[7]。耐旱性强的水稻(Oryza sativa)品种在30~50 cm 土层中具有更高的根系长度、根数量和根重,有助于植物吸取更深层的土壤水分而避免干旱胁迫[8]。更有研究表明,西北半干旱区生态恢复中禾本科(Poaceae)幼苗根系在30~60 cm土层中的总根长和总根表面积远高于豆科(Fabaceae)植物,分别是豆科植物的6 倍和7 倍,相应地,禾本科植物表现出比豆科植物更强的抗旱性[9]。不同植物应对土壤水分和土壤质地表现出具有差异的抗逆性和适应性。根系的生长具有很强的可塑性[10],尤其在逆境胁迫下,根系的形态建成和根系构型显著影响植物对水分的获取[11]。
草坪草、饲草和牧草等草本植物由于对环境具有较强适应性,占据了两倍于世界粮食作物的土地面积[12]。禾本科牧草为全球近百万草食家畜的生产和无数野生动物的生存提供了保证。干旱胁迫严重地限制了农业生态系统的初级生产(牧草)和次级生产(动物)[13]。在全球气候变暖、干旱区愈加干旱的严峻形势下(IPCC 2013)[14],研究耐旱型牧草的生存策略和适应性机制,对推进牧草育种、优化草地管理措施和畜牧业可持续发展具有重要意义。
多年生黑麦草(Lolium perenne)是世界温带地区最重要的禾本科牧草之一,原产于南欧、中东、北非及中亚。1677年英国首先栽培,后随人类活动而遍布世界各地。雀麦(Bromus valdivianus)是一种耐旱的速生型多年生禾草,原产于南美洲南部温带湿润气候区[15-16]。研究发现,多年生黑麦草和雀麦在20%~25%植物有效水(plant available water,PAW)的极端干旱条件下表现出较强的抗性,具有相近的产草量和草品质[15-18]。这两种牧草以不同的生长策略适应干旱环境,例如不同的分蘗能力、叶片伸展速率和根系深度[18-19]等。但是,对这两种优质禾草根系的干旱适应性方面的研究较少。因此,本研究选择这两种禾本科牧草为研究对象,分析其在干旱条件下根系生长状态和根系形态变化特征,探寻其在干旱胁迫下根系的适应性,为推进禾本科牧草育种,更好地开发利用分布广泛的禾本科牧草资源提供理论依据。
1 材料与方法
1.1 材料的种植
供试材料多年生黑麦草和雀麦由新西兰梅西大学López Ignacio F教授提供。
采用温室土壤水分控制盆栽试验,于2018年9月开始至2019年3月在新西兰梅西大学(40.37°S、175.61°W)农业与环境学院和植物生长中心(Plant Growth Unit,PGU)实施。
种植基质由30%的马纳瓦图砂壤土和70%的细砂组成(体积比)。每60 kg 基质中加入长效肥60 g、速效肥30 g、白云石45 g,混合均匀后装盆。基质的化学性质如下:pH 6.3(土∶水=1∶2),有效磷35 mg·L—1,可交换钾3.4 me·kg—1,可交换钙24.0 me·kg—1,可交换镁6.0 me·kg—1,可交换钠<0.5 me·kg—1,阳离子交换容量30.0 me·kg—1,硫酸盐83 mg·kg—1。
2018年9月23日开始育苗。将种植基质装入上口径24 cm、下口径17 cm、深24 cm 的种植盆中。盆内左右两边各插入1 根内径4 mm 的滴灌管,管口接dripper,以引导水分渗入深层基质,dripper 入土约6~8 cm 深。每盆等距离扎24 个小孔(图1),每孔播2 粒种子。待种子出苗以后,每孔留1 棵健壮幼苗。正常水分供给至幼苗生长到12 周,此后开始土壤水分限制。为避免边缘效应,仅中部的10 株苗为供试材料。

图1 育苗钵种子穴分布图Fig.1 The seeds distribution in pots
1.2 土壤水分控制处理
分别取额外的多年生黑麦草和雀麦各3 盆,用TDR 土壤水分测定仪(Mini Trase with Soil-moisture TDR Technology,USA)确定基质的最大田间持水量(field capacity,FC)和植物永久萎蔫点(plant wilting point,PWP)分别为体积含水量的16%和2%。本研究土壤水分限制水平为20%~25%(FC-PWP)(对照)和80%~85%(FC-PWP)(极干旱)两个水平,分别标记为80%~85% PAW(plant available water,植物有效水;对照)和20%~25% PAW(极干旱)。
多年生黑麦草和雀麦的2 个土壤水分处理(80%~85% PAW 和20%~25% PAW)采用随机完全区组设计[20]。每种牧草的每个水分处理各2 盆,5 个重复区组,即每种牧草的每个水分处理各有10 盆,共计40 盆。牧草生长至2018年12月23日,即3 个月(90日龄),开始进行为期25 d 的干旱胁迫处理。
使用TDR 土壤水分仪每天测定并严格控制基质含水量。每种牧草和干旱胁迫水平分别设置5 个额外种植盆,放置在试验盆旁边,与试验盆相同时间、相同条件下进行播种和管理。每天测定这些种植盆的基质含水量,根据López 等[15]和Descalzi 等[21]的研究从多年生黑麦草和雀麦额外种植盆中收集土壤水分信息,以避免TDR 感应杆对试验盆土壤和根系产生影响和干扰。
通过这种方法,依据额外种植盆的含水量计算每个处理的日失水量进行补水。每日灌水量计算公式如下:

式中:I为灌水量(mL);IC 为水分处理所设定的基质含水量(%);WC 为实际基质含水量(%);SW 为每盆基质干重(kg);WD 为水的密度,此处取值1000(水的密度1 g·mL—1=1 kg·1000 mL—1)。
1.3 根系观察与测量
土壤水分限制处理第26 天(2019年1月17日)进行采样和测量。倒置种植盆取出所有植株,轻轻抖落基质,分离植株,清水冲洗根系。每个种植盆仅取位于中部的10 株作为供试材料。从每盆的供试材料中随机选取2 株,每株剪取1~2 个根在Leica MZ12 体视显微镜(德国)下观察根段和根尖外观形态,5 次重复。
另从每盆的供试材料中再随机选取2 株,剪取全株根系进行分析,10 次重复。根系分析采用Epson Perfection V700 Photo Scanner 扫描仪,进行8-bit灰度扫描(分辨率为400 dpi),用WinRHI-ZO(Pro.2013e)根系分析系统对扫描得到的根系灰度图像进行分析。得到总根长、平均直径、总表面积、总体积、根尖数和交叉数等根系相关参数的数据。
在剩余的供试材料中再随机选取2 株,由根颈处分离地上部茎叶和地下部根系,分别置于烘箱,70 ℃烘至恒重,电子天平称量干物质重,10 次重复。
1.4 数据统计分析
对获取的数据采用Minitab18 Statistical Software(美国)进行数据统计分析。首先对单株的地上部分干重、根系干重及根系扫描所获得的根长、平均根直径、根表面积、根体积、根尖数和交叉数多变量进行了正态分布的Ryan-Joiner 检验(与Shapiro-Wilk 类似)[22]。其中地上部分干重、根系干重和平均根直径符合正态分布;对非正态分布的根长、交叉数和根尖数进行了常用对数转化,根表面积、根平均直径和根体积进行了平方根转化,使所有变量均符合正态分布。然后结合两种牧草和2 个土壤水分处理进行了方差齐性检验。最后在满足正态分布和方差齐性一致的情况下,对变量进行Tukey 法检验的多变量方差分析[23]。
对地上部分干重、根系干重、根冠比、根长、平均根直径、根表面积、根体积、根尖数和交叉数9 个变量进行相关性分析和主成分分析(Principal Component Analysis,PCA)。在PCA 分析中,根据相关矩阵的特征分析,提取第1 主成分(PC1)和第2 主成分(PC2),并对所有数据变量的PC1 和PC2 得分进行GLM ANOVA 分析,以获取多年生黑麦草和雀麦两种牧草响应水分胁迫的信息。
2 结果与分析
2.1 极干旱下两种牧草的根系外观特点
多年生黑麦草和雀麦在不同土壤水分条件由根颈至根尖形态均有差异(图2)。80%~85% PAW 处理下,须根生长至一定长度就开始大量生成侧根(图2 A1,C1)。根尖由根冠(图2 A4、B4、C4、D4 根端白色区顶部)、分生区(图2 A4、B4、C4、D4 根端白色区)、伸长区(图2 A3、B3、C3、D3)和根毛区(图2 A2、B2、C2、D2)组成。多年生黑麦草和雀麦的根尖差异较大。雀麦根毛密而长;黑麦草根毛稀疏,根毛区不如雀麦发达(图2 A2 vs C2)。雀麦分生区细胞幼嫩、致密,呈白色,逐渐过渡到伸长区,伸长区较长(图2 A4);黑麦草分生区细胞排列呈圆锥形,与伸长区细胞的圆柱形排列分界明显,伸长区粗而短(图2 C4)。
20%~25% PAW 极干旱水分胁迫下,雀麦根系纤长、粗细均匀,根毛区依然发达,根毛长而密,吸水能力较强,外观形态未见明显受害;多年生黑麦草根系粗细不均匀,部分根段出现畸形(图2 D1-D4 星号处)。
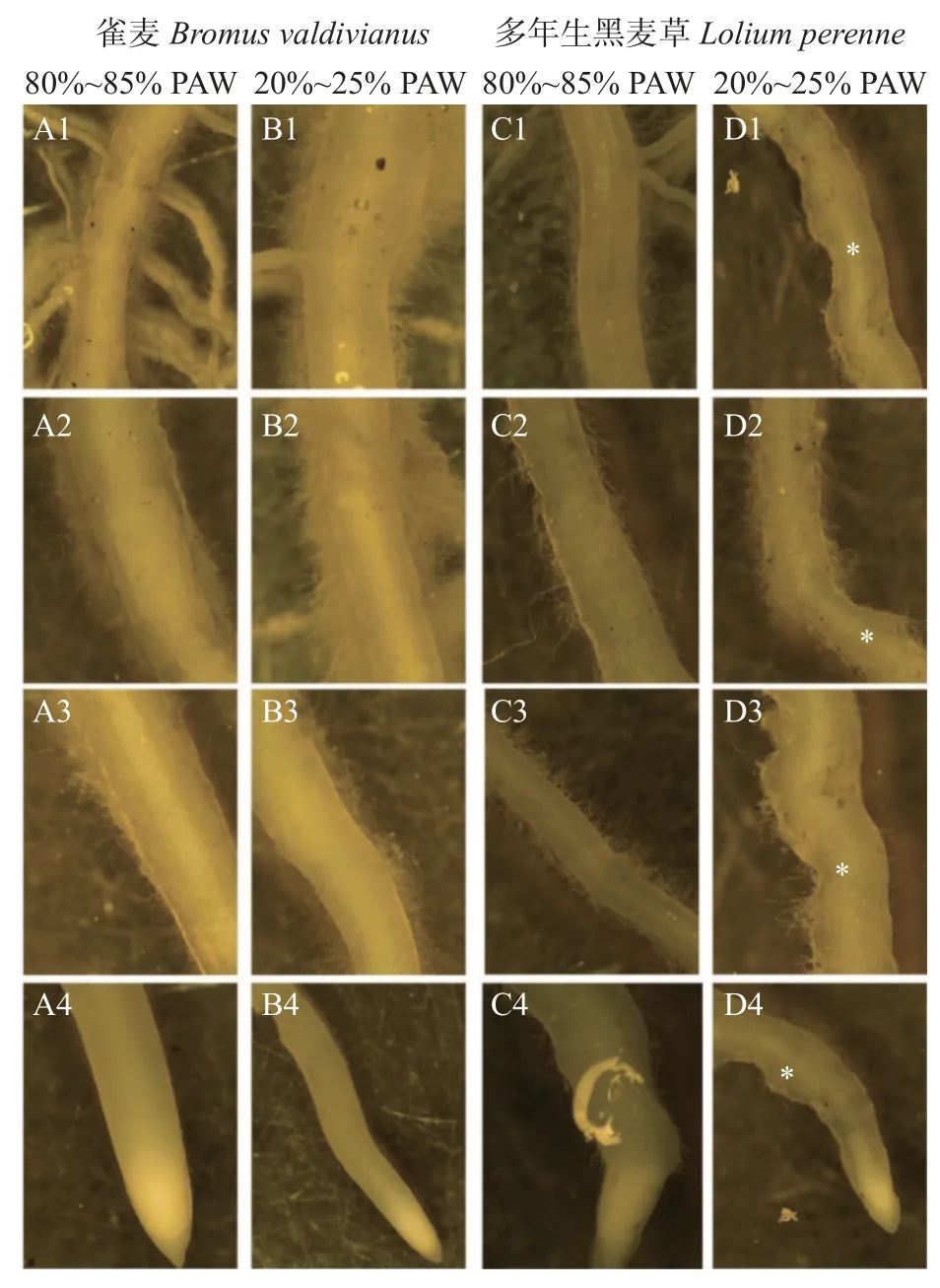
图2 80%~85%植物有效水(PAW)和20%~25% PAW土壤水分条件下多年生黑麦草和雀麦根段与根尖图Fig.2 Root segments and root tips of perennial ryegrass(Lolium perenne)and pasture brome(Bromus valdivianus)under soil water conditions of 80%—85% plant available water(PAW)and 20%—25% PAW
2.2 极干旱下两种牧草的根系生物量积累
雀麦和多年生黑麦草地上部分茎叶和地下部分根系的生物量积累和分配没有显著的种间差异(表1),20%~25% PAW 处理与80%~85% PAW 处理间也无显著的组间差异,牧草种与水分处理的交互作用对干物质积累无显著影响。由于不同器官的干物质积累无显著差异,造成根冠比也无显著的种间和组间差异。

表1 80%~85%植物有效水(PAW)和20%~25% PAW 土壤水分条件对多年生黑麦草和雀麦地上部分和地下部分干物质积累、根冠比的影响Table 1 Shoot dry weight,root dry weight and root to shoot ratio of perennial ryegrass(Lolium perenne)and pasture brome(Bromus valdivianus)grown under soil water conditions of 80%—85% plant available water(PAW)and 20%—25% PAW
2.3 极干旱下两种牧草的根系形态学特点
将20%~25% PAW 土壤水分条件下生长的雀麦和多年生黑麦草的须根系由根颈处剪下,对根系进行图像扫描。雀麦和多年生黑麦草须根系由根结发育生长的根和根上由根原基发育形成的侧根组成。雀麦侧根较短,上半部分的侧根分枝呈鲱鱼型,下半部分呈二分枝-鲱鱼型,且下半部分侧根分枝粗而短(图3 左图箭头处),局部分枝密度高。多年生黑麦草根系较长,侧根分枝呈二分枝-鲱鱼型,侧根细而长(图3 右图)。

图3 20%~25%植物有效水(PAW)土壤水分条件下雀麦和多年生黑麦草根系扫描图Fig.3 Root scanning images of pasture brome(Bromus valdivianus)and perennial ryegrass(Lolium perenne)under soil water condition of 20%—25% plant available water(PAW)
根系扫描显示,雀麦和多年生黑麦草单株根系的总长度、总根表面积、根平均直径、根尖数和分叉数存在极显著差异(P≤0.01),而根系总体积无显著差异(表2)。多年生黑麦草根系发达,根量极显著多于雀麦,其根总长度、根面积、根尖数和分叉数分别是雀麦根系的1.76 倍、1.46 倍、2.21 倍和2.05 倍。多年生黑麦草根细而多;雀麦与黑麦草相比,根少而粗。雀麦和多年生黑麦草根平均直径分别为0.315 mm 和0.259 mm。
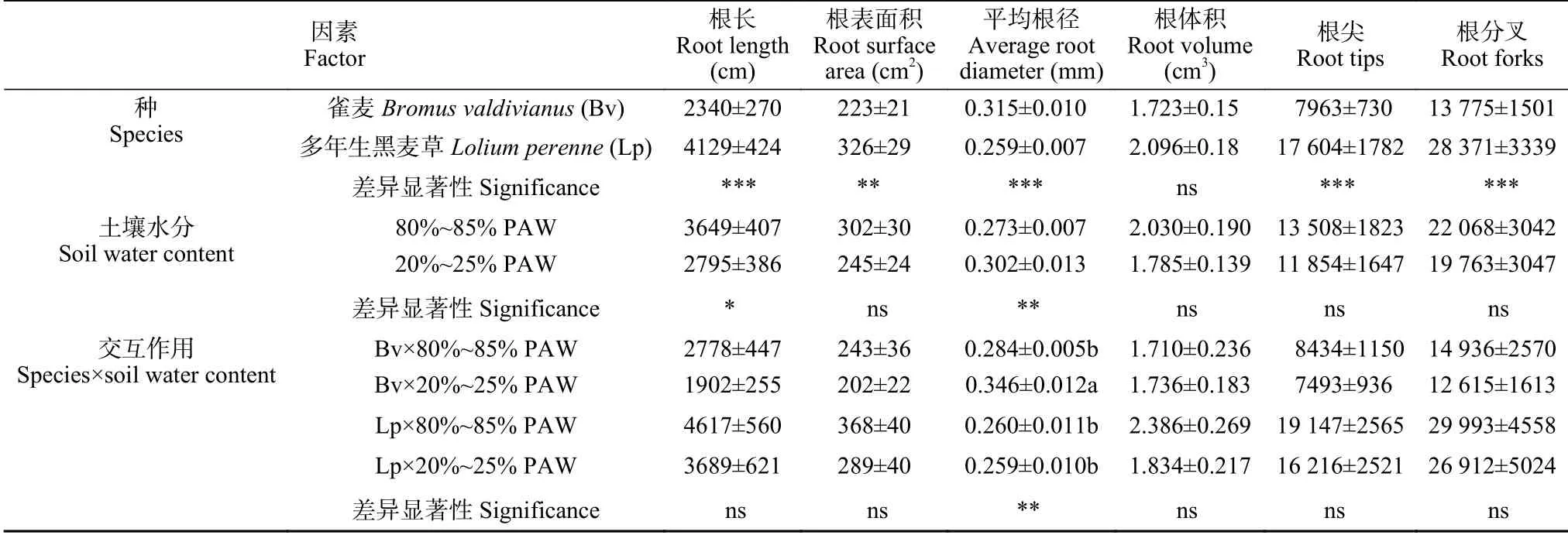
表2 多年生黑麦草和雀麦在80%~85%植物有效水(PAW)和20%~25% PAW 土壤水分条件下根系总长度、总根表面积、根平均直径、根体积、根尖数和分叉数Table 2 Root length,surface area,average diameter,volume,tips and forks of perennial ryegrass(Lolium perenne)and pasture brome(Bromus valdivianus)grown under soil water conditions of 80%—85% plant available water(PAW)and 20%—25%PAW
水分胁迫明显影响了雀麦和多年生黑麦草根系的生长,总根长显著减小(P≤0.05);20%~25% PAW极干旱水分胁迫显著促进了根系变粗(P≤0.01),平均直径从0.273 mm 增粗到0.302 mm。牧草种和水分胁迫的交互作用对根长、根表面积、根体积、根尖数和分叉数等方面均无显著影响,只对根系平均直径影响显著(P≤0.01),主要是由于干旱促进了雀麦根系的径向生长。
对多年生黑麦草和雀麦的根系组成进行分级分类,依根系平均直径(mm)分为[0.00,0.50)、[0.50,1.00)、[1.00,1.50)、[1.50,2.00)、>2.00 共5 类,分别对总根长、根表面积、根体积和总根尖数进行分类分析(图4)。两种牧草的须根系主要由<1 mm 的细根组成,分别占总根长的98.00%~99.56%、总根表面积的90.71%~97.45%、总根体积的72.99%~91.40%、总根尖数的99.81%~99.97%;≥1.00 mm 以上的根数量极少。其中,以[0.00,0.50)的毛细根为主,组成了总根长的85.86%~91.00%、总根表面积的62.54%~73.43%、总根体积的33.32%~43.32%和总根尖数的98.63%~99.29%。
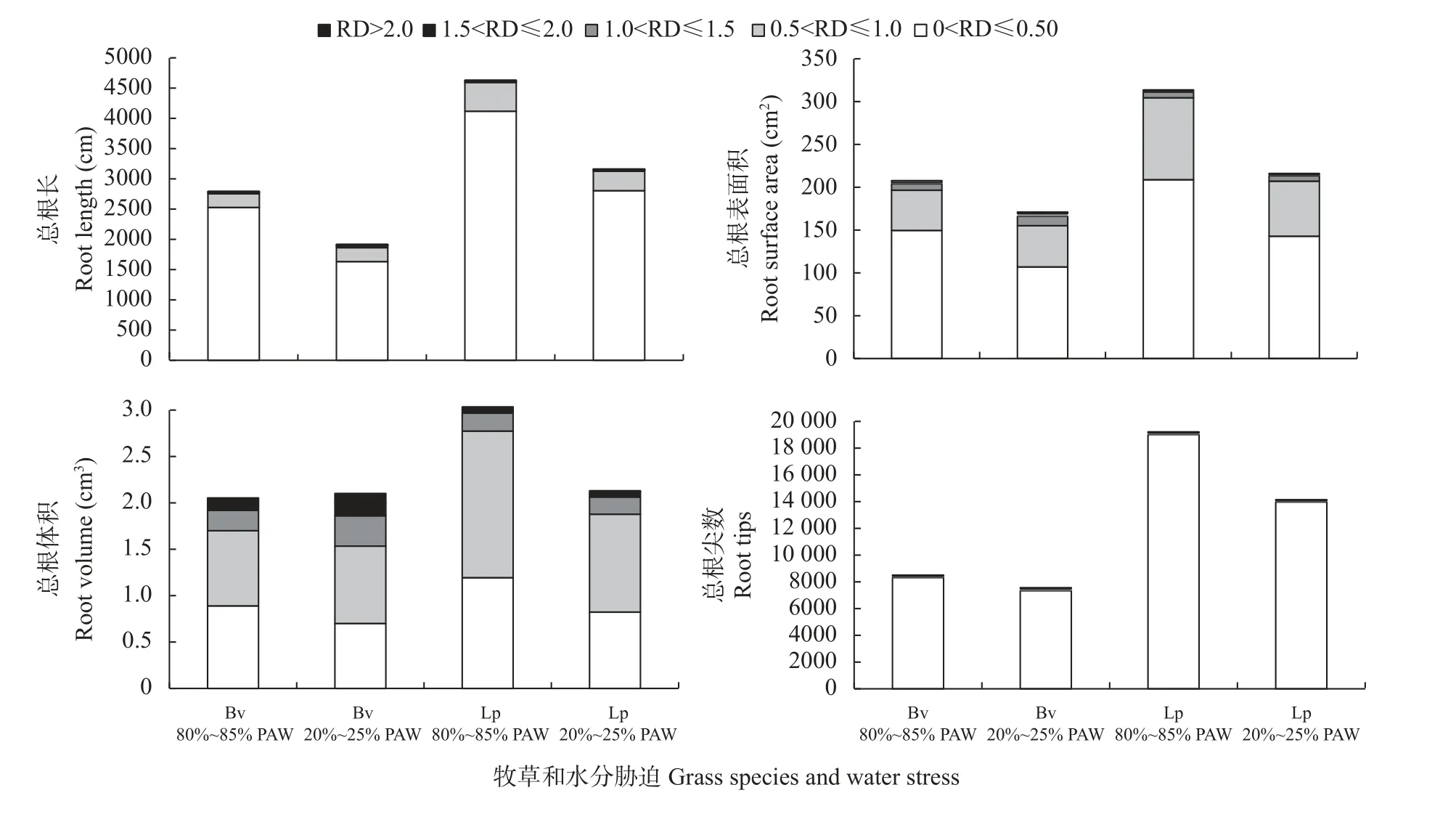
图4 80%~85%植物有效水(PAW)和20%~25% PAW 土壤水分条件下多年生黑麦草(Lp)和雀麦(Bv)不同直径(RD,mm)根系的总根长、根表面积、根体积和总根尖数Fig.4 Total root length,root surface area,root volume and total root tip number of different diameters(RD,mm)roots of perennial ryegrass(Lolium perenne,Lp)and pasture brome(Bromus valdivianus,Bv)under soil water conditions of 80%—85% plant available water(PAW)and 20%—25% PAW
2.4 多年生黑麦草和雀麦生物量、根系形态变量的相关性研究
对80%~85% PAW 正常灌水和20%~25% PAW极干旱胁迫下,多年生黑麦草和雀麦的地上部分和地下部分生物量干重、根冠比及须根系的长度、表面积、平均直径、体积、根尖和根分叉数量,共9个变量进行了相关性分析(表3)。极度干旱胁迫下两种牧草地上部分对阳光、地下部分对水分的获取均受到限制,干物质的积累和分配也受到一定的制约。本研究地上部分茎叶干重与地下部分根干重存在显著正相关关系(P≤0.01)。根系生物量与根系生长呈显著正相关:根干重与根长、根表面积、根体积、根分叉和根尖数量具有极显著正相关(P≤0.01或P≤0.001)。根长、根表面积、根分叉、根尖数量等反映根系数量的变量之间相互极显著正相关(P≤0.001),反映根系大小的根平均直径与反映根系数量的根长、根表面积、根分叉、根尖数量变量之间极显著负相关(P≤0.01 或P≤0.001)。

表3 多年生黑麦草和雀麦生物量、根系形态变量间的相关性Table 3 Correlation between variables of root biomass and root morphologh of perennial ryegrass(Lolium perenne)and pasture brome(Bromus valdivianus)
2.5 多年生黑麦草和雀麦生物量、根系形态学变量的主成分分析
对9 个变量进行主成分分析(principal component analysis,PCA),根据相关矩阵的特征值,提取主成分1(PC1)和主成分2(PC2),两者累积方差贡献率为0.784(表4)。说明这两个主成分可以解释9 个变量78.4%的变异,其中,PC1 占63.3%,PC2 占15.1%(表4)。
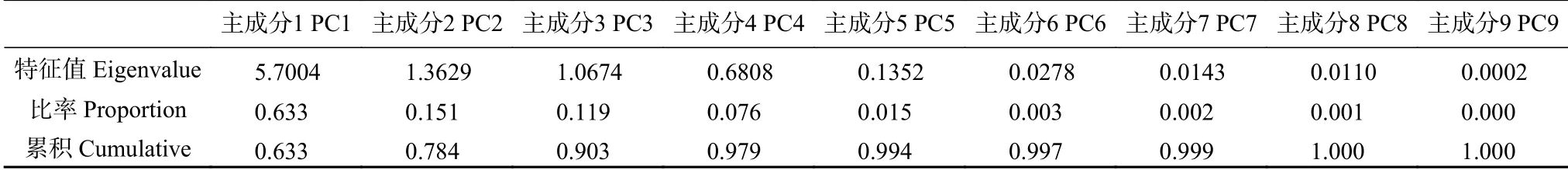
表4 多年生黑麦草和雀麦生物量和根系形态9 个变量主成分分析相关矩阵的特征分析Table 4 Eigenvalues of the correlation matrix of 9 variables of biomass and root morphology in principal component analysis of perennial ryegrass(Lolium perenne)and pasture brome(Bromus valdivianus)
根据9 个变量的特征向量的报告结果(表5),提取的前两个主成分可以记为:

表5 多年生黑麦草和雀麦生物量和根系形态的9 个变量主成分分析的特征向量估计结果Table 5 Eigenvector estimation results of 9 variables of biomass and root morphology in principal component analysis of perennial ryegrass(Lolium perenne)and pasture brome(Bromus valdivianus)

PC1 与根干重、根长、根表面积、根体积、根分叉、根尖数正相关,且因子荷载为0.315~0.411,处于中等水平,反映牧草地下部分根系的数量;PC2 与茎叶干重高度正相关,因子荷载0.783,反映牧草地上部分茎叶生长状况,与根直径也正相关,因子荷载较低,为0.389,中等相关水平。
将9 个变量的两个主成分得分绘制散点图(图5),并分别对所有观测变量的PC1 和PC2 估计值进行方差分析(表6)。第1 主成分PC1 反映黑麦草和雀麦两牧草之间具有极显著差异(表6,P≤0.01)。在散点图上,雀麦分布于PC1 坐标轴的左侧(图5A),与根平均直径正相关(图5B);多年生黑麦草位于PC1 右侧(图5A),与根长、表面积、根尖数、根冠比等体现根系数量的变量正相关(图5B)。结合方差分析说明这9 个变量的63.3%的变异主要来源于牧草种间差异(表6),多年生黑麦草的根量极显著多于雀麦根量(表6,PC1 与species 的P值≤0.01)。此外,两牧草20%~25% PAW 在PC1 坐标轴方向上分别位于80%~85% PAW 的左边(图5A),说明水分胁迫限制了根的生长,但未达显著水平(表6)。

图5 多年生黑麦草(Lp)和雀麦(Bv)生物量干重和根系形态9 个变量的第1 主成分和第2 主成分得分散点图Fig.5 Scatter plots of the first and second principal components of 9 variables of biomass and root morphology of perennial ryegrass(Lolium perenne,Lp)and pasture brome(Bromus valdivianus,Bv)
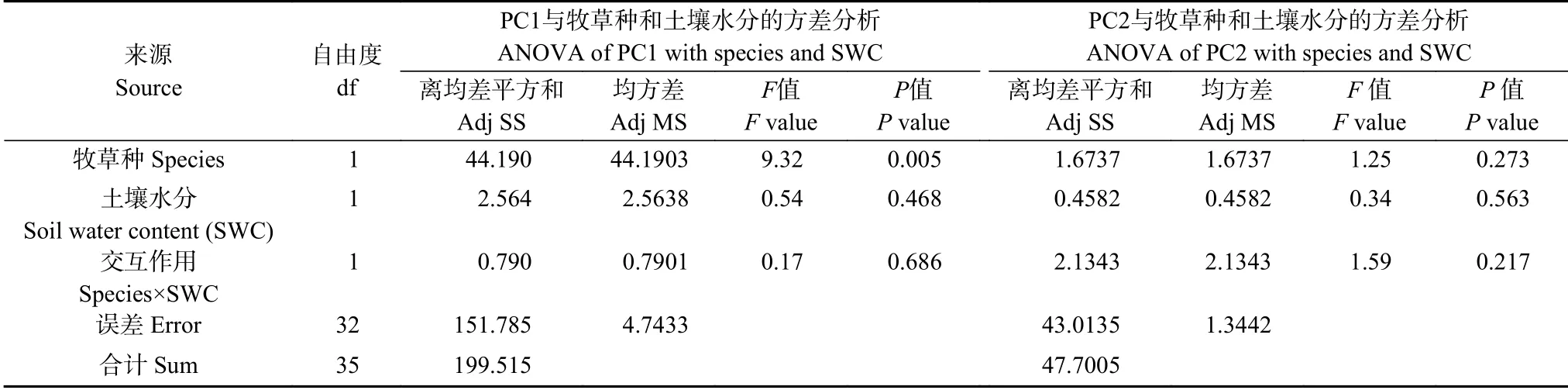
表6 一般线性模型:第一主成分、第二主成分与牧草种、土壤水分含量的方差分析Table 6 General linear models:ANOVA of PC1,PC2 and forage species,soil water content
PC2 反映地上部分茎叶干重在牧草种间和水分胁迫间虽有差异,但未至显著性差异(表6),只能解释9 个变量间15.1%的差异。
3 讨论
3.1 牧草生物量积累对干旱胁迫的适应性改变
生物量积累和分配情况可反映植物在不同环境下的生存策略和适应机制,当土壤水分和养分条件发生变化时,地上部分茎叶和地下部分根系的生物量分配也将随之发生改变[11,24]。本研究中雀麦和多年生黑麦草(90日龄)的地上部分干重和根系干重没有显著的种间差异,充分灌水和极干旱胁迫两种土壤水分模式之间也无显著差异,根冠比也无显著的种间和组间差异。这一结果与前人研究结果不尽相同。大多数研究表现出干旱胁迫促进了生物量向根系积累,满足根系向深层土壤的生长,造成根冠比增大[24-25]。25日龄的野生型二粒小麦渐渗杂交系(IL20)给予9 d 的干旱胁迫后,生物量积累表现出茎叶干重略有下降(—8%),根干重略有增大(+11%),从而引起根冠比显著增加21%[24];大麦(Hordeum vulgare)幼苗随土壤含水量由85% FC 减少至35% FC,根冠比和根分支数均呈上升趋势[25];干旱胁迫影响了大麦生殖生长期生物量的积累,使大麦从扬花期到蜡熟期根冠比不断升高,最终导致小穗和千粒重分别减少20.16%和7.75%,侧根数显著增大[26]。Smucker和Aiken[27]指出,在水分胁迫下,光合产物优先分配给根系,使根冠比增大;反之,则根冠比减小。
根冠比通常能够反映由非生物因素和生物因素引起的地上生物量和地下生物量之间的投资差异。本研究结果两种牧草受土壤水分胁迫而根冠比无显著变化,不同于前人的研究结果。说明植物生物量的积累与分配不仅与土壤水分条件有关,而且可能与植物种植密度、种植方式、生育时期等有关。
最优分配理论(optimal partitioning theory,OPT)是基于植物地上部分和地下部分在生物量分配上存在着权衡的假设基础之上。该理论认为植物分配生物量是为了获得最受限制的资源[28-29]。如果植物的生长更强烈地受地下因素(如土壤水分、土壤养分、土壤物理性质、土壤化学性质等)的影响,植物将分配相对更多的生物量给根系;当植物的生长更强烈地受地上因素(温度、光照、湿度、CO2浓度等)的影响,植物则分配相对更多的生物量给茎叶[30]。不同于前人大多选择植物幼苗为研究对象,本文是针对牧场生产实践中,牧草主要是以生长3 个月以上、可刈割的成株为利用对象;且牧场或草场中的牧草由于分蘗或混播而存在较强的种内竞争或种间竞争的实际情况而展开的研究。所选多年生黑麦草在野外生长环境下是浅根型草本,雀麦是深根型牧草[18]。本研究在同一种植盆中,种植了24 株同种牧草,生长90日龄的成株进行为期25 d 的水分胁迫处理,存在较强的种内竞争。无论是地下根系对水分、养分的竞争,还是地上茎叶对阳光的争夺都十分强烈,说明在当前的生长状况下两种牧草对地下水分的需求和对地上阳光的需求同样重要、同样迫切,进而严重影响了生物量的积累和光合产物的分配。此外,两种牧草生物量积累的结果进一步说明植物器官在复杂环境中具有产生一系列不同的相对适合的表型的潜能;植物根系作为感受土壤水分和养分变化最敏感的器官,其生长具有较强的表型可塑性[10,31-32],根系的生长机制随环境条件(生物环境和自然环境)的改变而能够及时地做出适应性改变。
3.2 干旱胁迫下牧草根系形态的改变
不同植物、同种植物不同生育时期、不同基因型以及在不同的栽培条件下,植物根系的生长、发育和形态对干旱胁迫的响应并不一致,表现出极大的可塑性和变异性。本研究,20%~25% PAW 极干旱水分胁迫明显抑制了雀麦和多年生黑麦草须根的生长,总根长显著减小,根系总表面积、体积和根尖数虽有减少,但未至显著水平。这与前人对小麦、白羊草(Bothriochloa ischaemum)等草本植物根系的研究结果基本一致[31,33]。干旱胁迫抑制了小麦幼苗根数和根系表面积,且不同耐旱型小麦品种对干旱胁迫的适应性存在差异[34];随干旱胁迫程度持续加重,白羊草根系总根长和根表面积显著下降[31]。
但是,不同植物遭受水分胁迫时也有不同的表现。干旱胁迫促进了小麦总根长显著增加,根系直径明显降低[34]。干旱抑制了白羊草根系生长,但对白羊草根系平均直径影响不显著[31]。干旱胁迫促使大麦拥有了更多的侧根[26]。本研究20%~25% PAW 极干旱水分胁迫虽然明显降低了雀麦和多年生黑麦草的总根长,但对根系直径的影响不同。干旱胁迫促进了雀麦根径的加粗,对黑麦草根径无显著影响(表2,图5)。从根系扫描图观察发现雀麦较粗的根系是根端分枝生长的侧根(图3)。结合主成分分析结果,雀麦20%~25% PAW 与根直径正相关(图5)。因而推测,雀麦根端侧根的生长是其适应干旱条件做出的积极响应。
植物总根长体现了根系的生长和伸长能力;根系的分叉数越多,根系在土壤中的分布范围越广;根尖反映根系的生理代谢能力;根表面积是根系吸收水分和养分的重要指标[35]。本研究中根系扫描结果显示,多年生黑麦草单株根系的总长度、总根表面积、根尖数和分叉数极显著多于雀麦(表2,P≤0.01)。多年生黑麦草的根系发达,数量极显著多于雀麦,根系生长范围可能也广于雀麦根系,因而汲取水分的活力和机会也高于雀麦。
根毛是由表皮细胞从根表面延伸到土壤中的管状细胞。根毛将根系固定在生长基质上,与土壤微生物区系相互作用,吸收土壤中的无机养分和水[36]。雀麦根毛密而长,且受干旱影响较小(图2 A2、B2),雀麦根尖的根毛区发达,根毛的增殖大大增加了根系的总表面积,相应增加了根系与土壤接触面积,保证了雀麦根系对水分的有效汲取。
4 结论
对多年生黑麦草和雀麦进行干旱胁迫处理,由于试验中牧草植株存在着强烈的种内竞争和水分胁迫的双重压力,多年生黑麦草和雀麦光合产物的积累和分配在地下和地上器官之间无明显种间差异;而两种牧草根系的生长和根形态差异较大。多年生黑麦草根量显著多于雀麦,体现在根长、根面积、根尖数和分叉数显著高于雀麦。多年生黑麦草获取土壤水分主要以根数量(量)取胜,采取广泛吸水的适应策略。雀麦根尖的根毛区发达,根毛长而密,且受干旱影响较小;此外,干旱促进了雀麦根系的径向生长。雀麦汲取水分以根毛发达,侧根分枝密度高(质)为主,采用确保有效吸水的适应策略。
猜你喜欢
奥秘(创新大赛)(2022年8期)2022-09-14
农业工程学报(2022年10期)2022-08-22
农业工程学报(2022年5期)2022-06-22
水土保持学报(2022年3期)2022-05-26
中国果业信息(2021年10期)2021-12-07
现代农村科技(2021年11期)2021-11-05
小小说月刊·下半月(2021年8期)2021-09-05
中国饲料(2021年8期)2021-05-17
新农业(2019年9期)2019-09-17
动漫界·幼教365(大班)(2018年3期)2018-05-14
