
藏东木本植物群落功能性状分布与环境的关系
2022-12-16 04:49黄汐月黄梦月杨文静石松林李景吉彭培好王国严
生态学报 2022年22期
黄汐月,陈 卓,黄梦月,杨文静,石松林,李景吉,彭培好,王国严
1 成都理工大学生态资源与景观研究所, 成都 610059
2 成都理工大学旅游与城乡规划学院, 成都 610059
3 成都理工大学生态环境学院, 成都 610059
植物功能性状是指植物体具有的与其定植、存活、生长和死亡紧密相关的一系列核心属性,反映了植物对其生长环境的适应,是植物与环境相互作用的结果[1—2]。大量研究表明:植物功能性状可以通过权衡和协同变化,形成性状间最佳的功能组合,更好地适应变化的环境[3—6]。叶干物质含量(Leaf dry matter content, LDMC)、比叶面积(Specific leaf area, SLA)、木质密度(Wood density,WD)等结构型功能性状能反映植物的生存适应策略,可代表植物功能性状变异的主要维度,并且测量相对便捷[4,7],因此被广泛应用于植物功能性状与环境关系的研究中。Kergunteuil等[8]发现在瑞士阿尔卑斯山高海拔地区,功能性状表现出株高低而SLA大的权衡方式,王超等[9]发现色季拉山急尖长苞冷杉通过协调多个性状(SLA较小而LDMC较大,叶面积与叶体积协同变化)来适应高寒环境;齐威等[10]发现青藏高原东部常见的4 科植物为应对寒冷的气候条件采取种子较大而SLA较小的权衡模式。
质量比假说认为物种功能性状对生态系统的影响程度与该物种在群落中的优势度密切相关[11]。Garnier等[12]为检验质量比假说,提出了群落加权平均性状值(Community-weighted mean, CWM),将群落物种相对丰富度与植物个体功能性状结合,代表群落水平上的功能性状[12—13]。群落水平上的功能性状能够更好地描述植物群落对环境变化的响应,正成为生态学研究的一个重要工具[13—14]。针对群落水平功能性状和环境之间的关系,学者们做了大量研究:Costa等[15]发现乞力马扎罗山植物群落水平功能性状对降水量的响应强烈;Gutiérrez-Girón等[16]发现西班牙地中海高山草原植物叶氮含量(Leaf nitrogen content, LNC)和叶片大小随土壤温度变化,LDMC与土壤pH有关;Standen和Baltzer[17]提出土壤活性层厚度是影响加拿大北方植物群落水平功能性状变化的主要因素;向响等[13]发现青海湖流域植物群落SLA、LNC和叶磷含量(Leaf phosphorus content, LPC)均随海拔的升高而显著增大。
研究群落对环境变化的响应,必须评估种内和种间变异对于植物功能性状变异的相对影响程度[18]。早期,物种间的性状差异被认为是群落物种共存的重要前提[19—20]。大量研究基于群落物种个体的性状平均值,以探讨性状的种间变异为主,导致性状的种内变异被忽略,认为性状的种间变异远大于种内变异[20—22]。在近期meta-analysis中,种内变异平均解释了群落内1/4的性状变异和群落间1/3的性状变异[23]。越来越多的研究证实,种内性状变异在植物群落对环境变化和群落构建的响应中发挥着基础性作用[18,24—26]。学者们对群落水平功能性状变异的来源展开了广泛探索: Roos等[18]发现种间变异是苔藓植物沿海拔梯度植物功能性状变异的主要驱动因素;Chelli等[27]的研究结果表明:植物功能性状变异在不同森林成熟度下主要由种间变异来解释;而Hu等[28]发现在氮沉降的影响下,叶片化学性状变异的主要来源是种内变异。这些研究能够帮助我们更好地把握不同植物群落对环境变化的响应规律。
青藏高原作为全球平均海拔(>4000 m)最高的独特自然地理单元,是我国乃至亚洲重要的生态安全屏障,也是全球气候变化的敏感区和生态脆弱带[29]。藏东昌都地区位于青藏高原东缘、横断山脉西北部,怒江、澜沧江、金沙江自北向南深切而过,形成了复杂的地形地貌和独特的植物区系特征,是青藏高原森林和灌丛生态系统的主要组分和国际生物多样性保护的热点地区。但由于历史交通条件等限制,该区域植被生态特征资料极为匮乏,与青藏高原生态屏障建设和保护目标不相适应。根据前人研究结果,我们假设青藏高原木本植物存在适应高寒环境的功能性状权衡;较大的海拔落差可能是驱动高原植物功能性状变化的主要因素;种间变异是植物群落功能性状随环境变化的主要成分。
本研究以藏东昌都地区森林灌丛优势木本植物为研究对象,在科学假设的基础上,拟解决以下3 个科学问题:(1)功能性状间是否存在权衡和协同变化,具体的权衡模式如何?(2)海拔是否驱动着植物功能性状变化,功能性状对环境变量的响应规律是怎样的?(3)种间变异是否在功能性状随环境变化中起主要作用?这些问题的解答有助于加深对该区域植物资源分布规律和生态功能的认识,促进区域自然资源和生物多样性保护。
1 材料与方法
1.1 研究区概况
研究区(97.20°—98.91°E,30.27°—32.05°N,海拔3143 —4628 m)位于青藏高原东部,重点集中在昌都地区(图1)。受青藏高原特殊的地理环境和独特的地形地貌影响,该区域的气候以寒冷为基本特点,年平均气温-2.1 —8℃;降雨集中,季节分布不均,年降水508 —621 mm,最暖季平均降水量329.4 mm,最冷季降水量15.2 mm。研究区内植被具有较强的耐寒性,能够忍耐冬季长达数月的积雪覆盖和冻土环境[30]。研究区内主要群落类型由低海拔向高海拔分别为:3100 —3700 m:砂生槐(Sophoramoorcroftiana)灌丛、川滇高山栎(Quercusaquifolioides)灌丛、钝叶栒子(Cotoneasterhebephyllus)灌丛;3500 —3800 m:糙皮桦(Betulautilis)林、川西云杉(Picealikiangensisvar.rubescens)林;3800 —4600 m:雪层杜鹃(Rhododendronnivale)灌丛、窄叶鲜卑花(Sibiraeaangustata)灌丛。
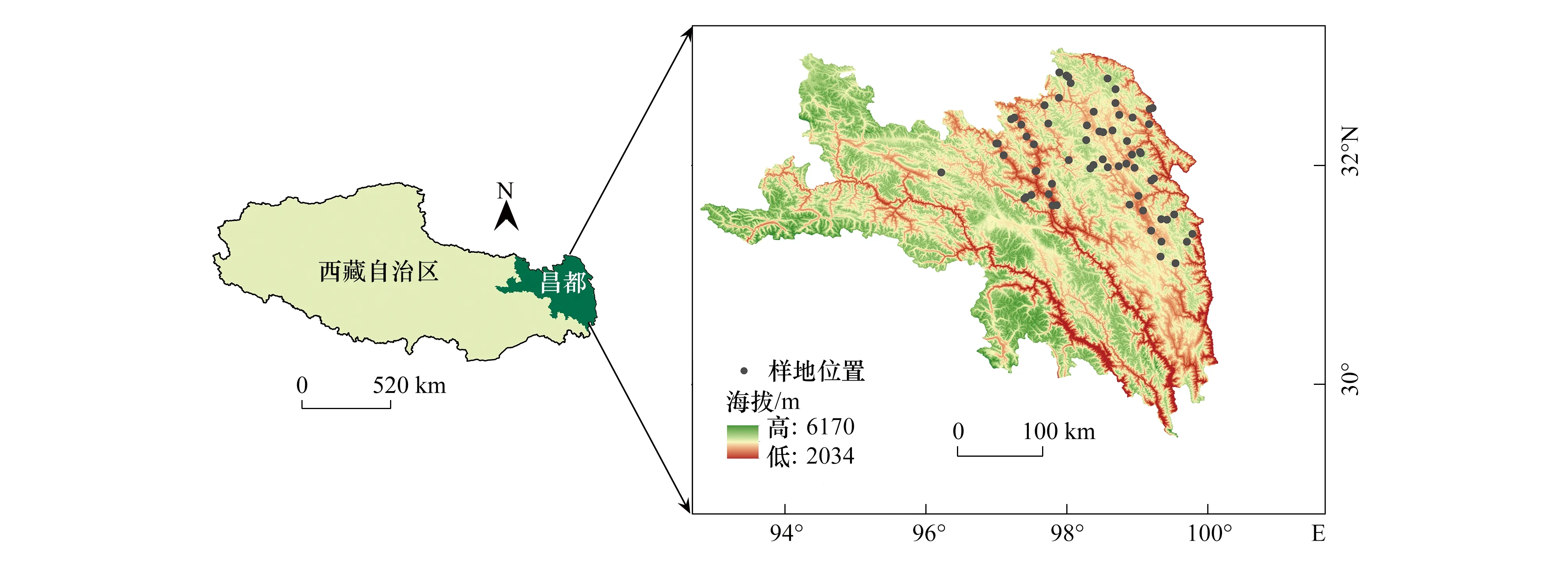
图1 研究区概况图
1.2 群落学调查
2021 年9 —10 月中旬,野外调查队在青藏高原东部通过分层随机抽样的方法布设样方,设置乔木样方20 mx20 m(共16 个),灌木样方5 mx5 m(共67 个),运用GPS、坡度测量仪测定样方的经纬度、海拔、坡度、坡向等基本信息。在选定的样方中进行植物群落调查,对胸径≥3 cm的乔木,采用每木检尺的调查方法,测量乔木的胸径、高度、枝下高、冠幅;对每种灌木的高度、基径、株丛数、盖度进行测量,并采集植物功能性状样品和土壤样品。
1.3 样品采集与测定
在每个植物样方中,同一物种选择10 —15 株长势良好、大小相对一致的个体作为取样植株,采集10 片新鲜完整的叶片和5 个长度15 cm左右的当年生小枝样品。为防止叶片脱水,样品采集后用被蒸馏水浸湿的滤纸包裹,装入自封袋,编号后置于冷藏箱中,带回实验室进一步处理。在每个样方的四个角及中心采集约200 g的表层(0 —10 cm)土壤样品,装入自封袋中,带回实验室,待筛去植物根系和砾石后,进行土壤理化性质的测定。在“WorldClim”(https://www.worldclim.org)上,根据样地经纬度运用ArcGIS提取对应的样地气候数据(年均温和年降水量)。本研究共选取11 个环境变量(表1)。
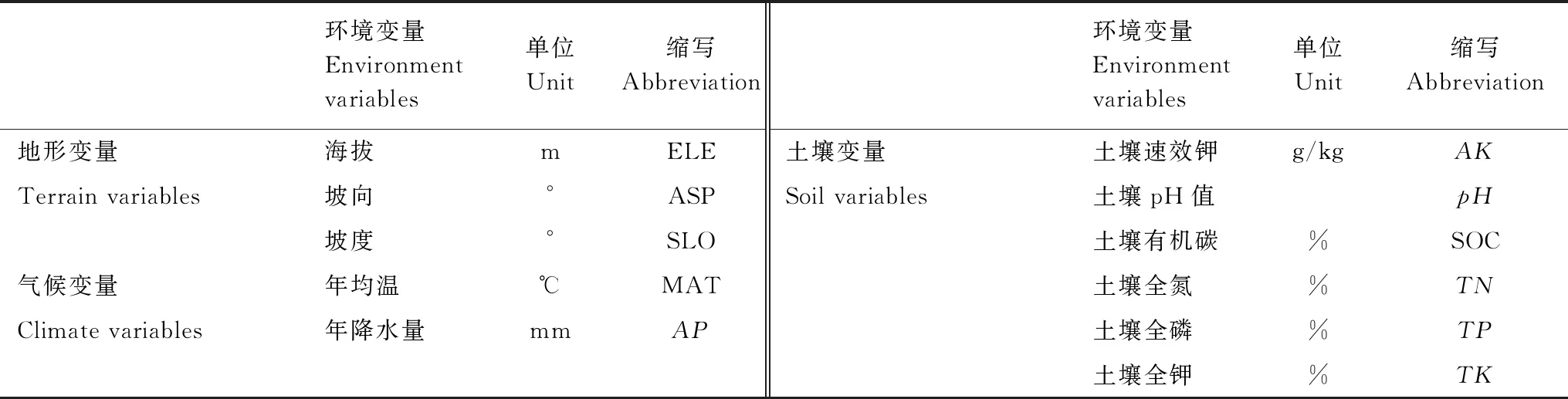
表1 11 个环境变量
参考功能性状手册[31],对各功能性状数据进行测量。用小刀削去枝条的树皮、韧皮部,仅留下木质部,利用排水法测量枝条的体积;用叶面积扫描仪(万深植物图像分析仪)获取叶面积;运用精度为0.0001 g的电子天平(上海力辰电子分析天平FA224C)测量叶片鲜重,之后放入烘箱,烘至恒重(65℃,72 h),测量叶片干重及枝条干重;运用精度为0.001 mm的电子螺旋测微器(日本三量电子外径数显千分尺螺旋测微器),避开叶脉,选择叶片前、中、后3 个部位对叶片厚度进行测量,取3 个部位的平均值作为叶片厚度值;叶片氮含量使用凯氏定氮仪用凯氏定氮法测得;叶片磷含量使用紫外分光光度计用钼锑抗比色法测得;叶片钾含量使用原子吸收分光光度计用火焰光度法测得。本研究共选取7 个功能性状(表2)。

表2 7 个功能性状的计算公式及生态学意义
1.4 研究方法
首先,利用物种的相对高度、相对基径和相对密度指标,计算物种的重要值(Important value,IV)。其次,将物种重要值作为权重,并与物种个体性状值结合,计算群落加权性状值(CWM),并对7 个性状的CWM进行“shapiro.test”正态检验,对不满足正态分布的CWM进行log对数转换。具体计算公式如下:
IV=(相对高度+相对基径+相对密度)/3
i为样方中第i个物种,s为样方物种总数,Pi为i物种在样方中的重要值,Ti为i物种在每个样方中的个体性状值。
进一步将PCA分析和线性回归模型相结合,揭示群落加权功能性状与环境变量之间的关系。最后,由于各CWM是由性状的种间和种内变异共同决定的,根据Lepš[33]提出的方法,采用方差分析和平方和分解的方法,在不同的环境变量下,对功能性状的变异来源进行分解。数据处理均在R4.1.2中进行,主要运用了以下程序包:“tidyvrse”[34];“dplyr”[35];“psych”[36];“MASS”[37];“ggplot2”[38]。
2 结果与分析
2.1 群落加权性状相关性分析
本研究对7 个群落加权性状进行了Pearson相关分析,结果见表3。总体来看,群落功能性状之间具有很强的相关性。具体而言:LDMC与SLA、叶体积(Leaf volume,LV)、叶钾含量(Leaf potassium content, LKC)、LPC呈不同程度的负相关,其中LDMC与SLA的负相关关系最为显著(P<0.001);LV与SLA、LKC呈极显著正相关(P<0.001),与其余性状无关;SLA与LPC、LKC呈极显著正相关(P<0.001);LPC与LKC呈显著正相关(P<0.01);WD仅与LPC呈正相关(P<0.05),与其他叶性状无关;只有LNC与其余性状均不相关。

表3 群落加权性状间Pearson相关系数矩阵
2.2 群落加权性状与环境变量之间的关系
本研究对11 个环境变量进行了PCA分析,结合Cattell碎石检验的结果,最终确定保留前两个主成分。前两个主成分共同解释了环境变量59.2%的方差,根据环境变量的载荷矩阵(表4),与PC1高度相关的变量有:土壤有机碳(Organic carbon, SOC)、土壤全氮(Total nitrogen,TN),且均与PC1呈正相关;与PC2高度相关的变量有:年均温(Annual mean temperature, MAT)、年降水量(Annual precipitation,AP),其中MAT与PC2呈负相关,而AP和ELE与PC2呈正相关。
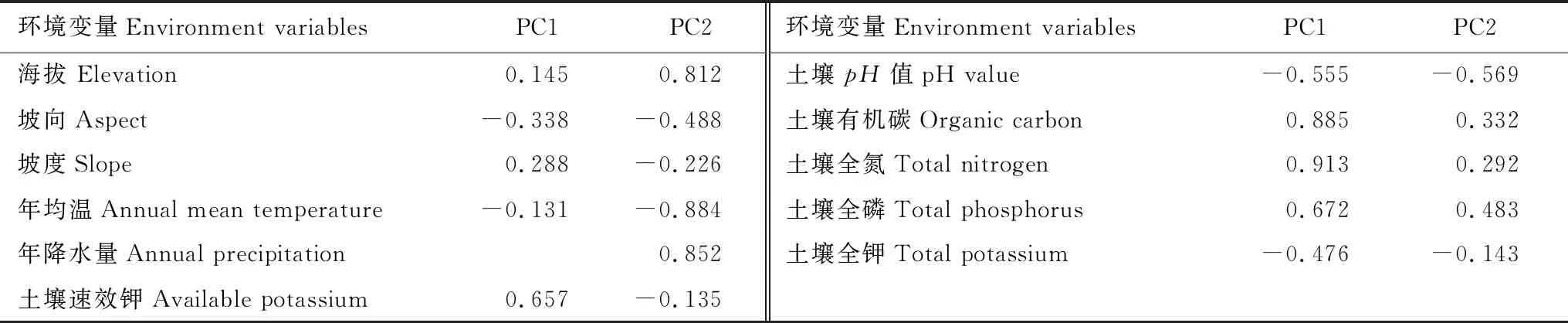
表4 环境变量载荷矩阵
因此,将11 个环境变量归为两类,分别为土壤养分变量(PC1)、海拔和气候变量(PC2)。进一步将这两类环境变量(PC1、PC2)作为解释变量,7 个群落加权功能性状作为响应变量构建线性回归模型,结果见表5。
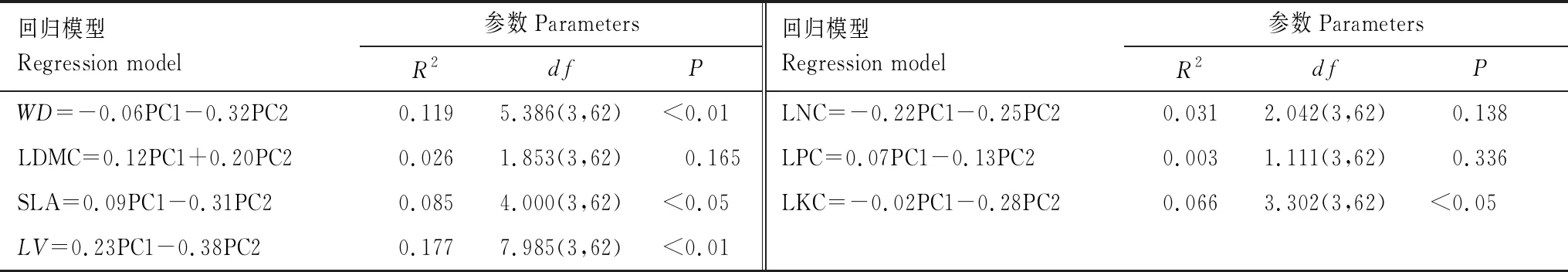
表5 功能性状与环境变量的PCA回归模型结果
从图2和图3可以得出以下结果:(1)LV随土壤养分含量的增加而增大;(2)海拔和气候变量对WD、SLA、LV、LKC均存在显著的负向影响;而(3)LDMC、LNC、LPC与这两类环境变量(PC1、PC2)均不存在显著关系。
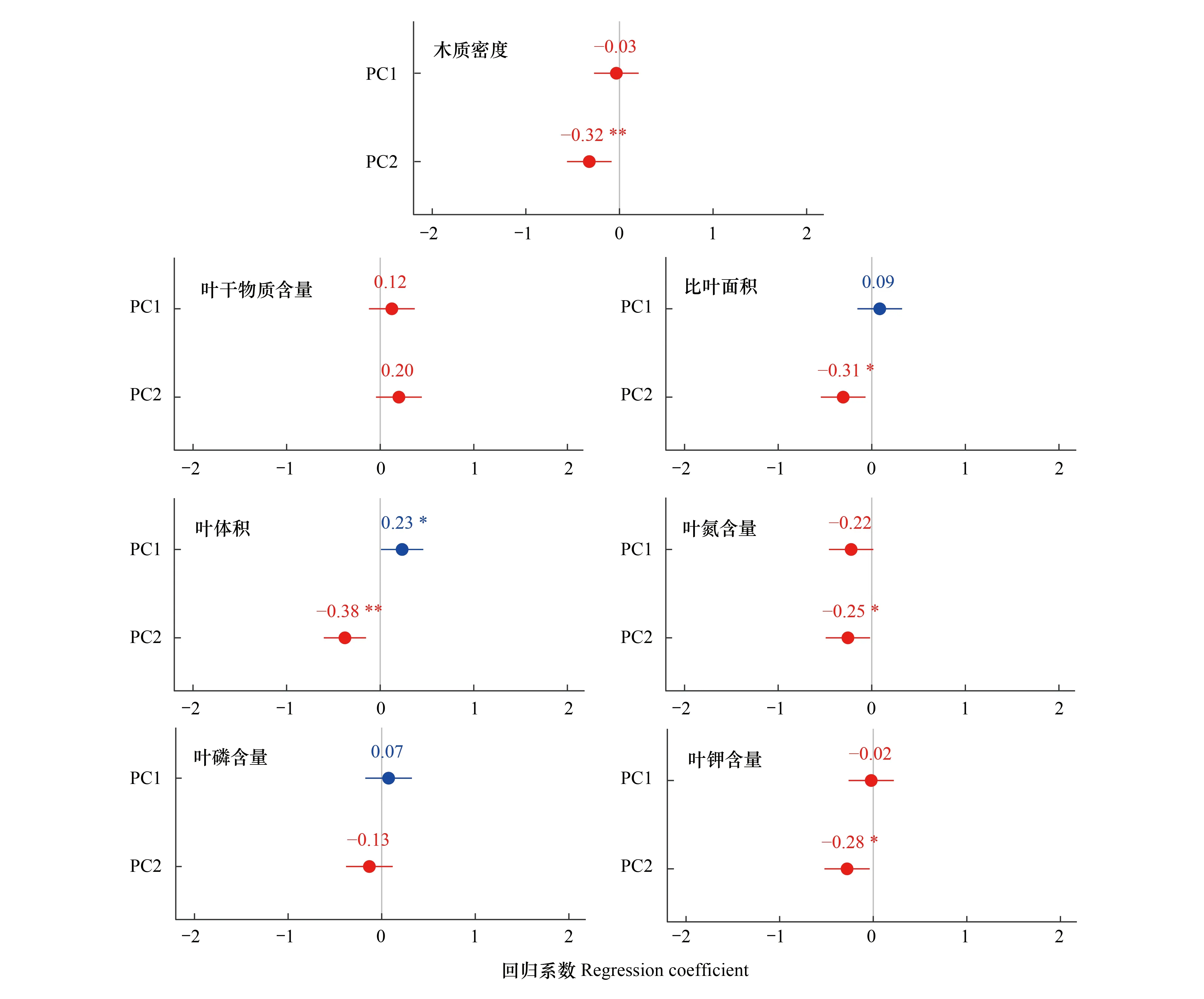
图2 群落加权功能性状与土壤养分变量(PC1)、海拔和气候变量(PC2)之间的关系
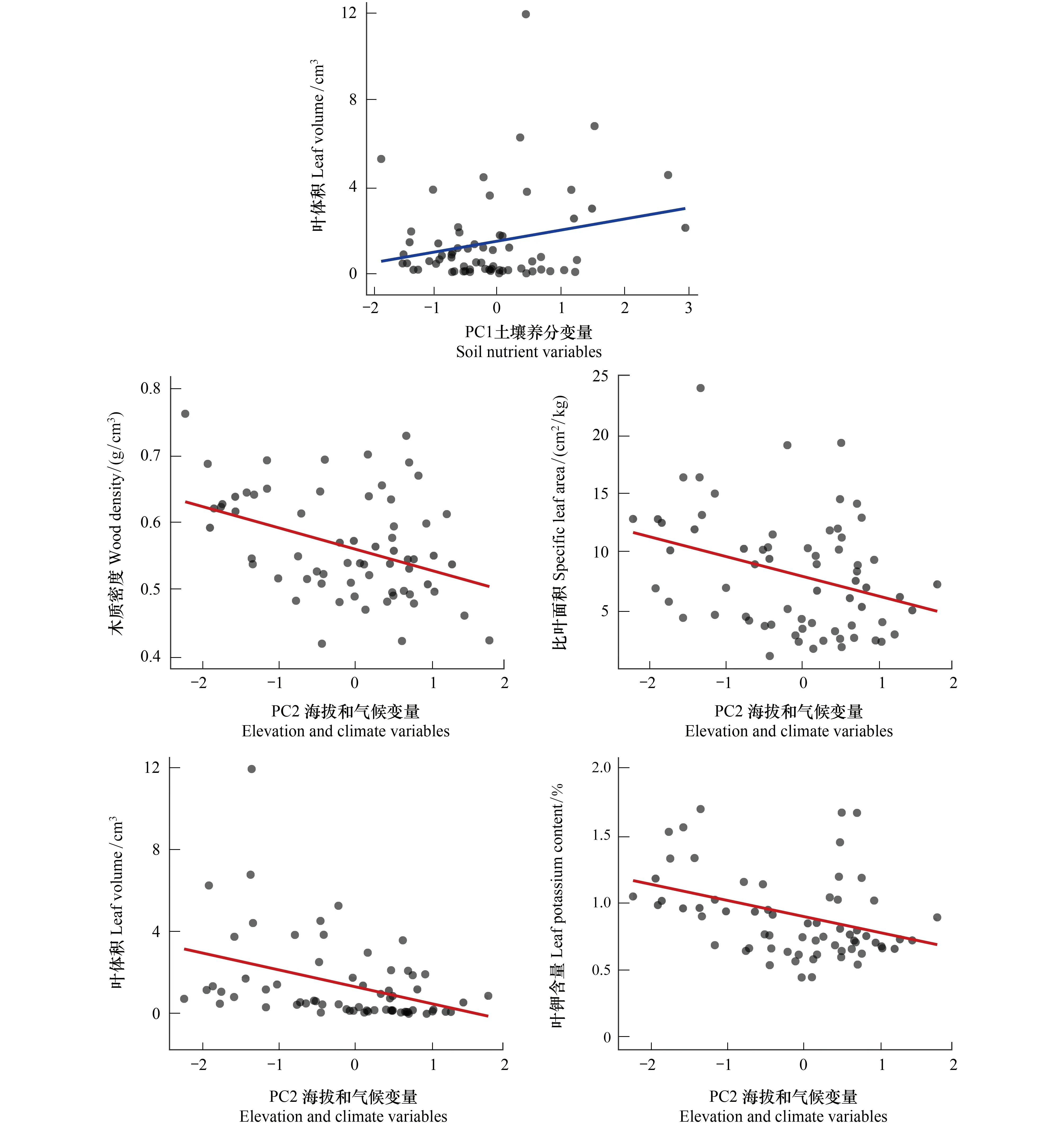
图3 土壤养分变量(PC1)、海拔和气候变量(PC2)对群落加权性状的显著影响
2.3 在海拔、年降水量和土壤养分(全磷)条件下对群落加权性状变异来源进行分解
本研究运用方差分析和平方和分解的方法,对每个性状的CWM的变异来源进行了分解。在11 个环境变量中,海拔(Elevation, ELE)、年降水量(Annual precipitation,AP)、土壤全磷含量(Total phosphorus,TP)对于群落加权性状变异的影响最显著。从图4中,可以得出以下结果:(1)在11 个环境变量中,海拔对群落加权功能性状变异的解释率最高;(2)7 个性状的种间变异解释率均大于种内变异;(3)种间变异与种内变异之间的协变均表现为负协变。(4)仅在LPC中,种间变异与种内变异均不显著,但两者之间存在负协变。
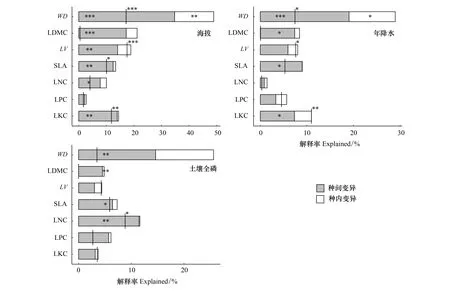
图4 群落加权性状变异来源分解
3 讨论
3.1 群落功能性状间的权衡与协同变化
植物可以通过各性状间的协同与权衡来适应环境的变化,最终形成一系列适应特定环境的最优性状组合[6,39]。研究不同地区植物功能性状间的相关关系,有助于理解不同生境间植物功能性状的适应策略,也有利于深入探索生态位的分化机理和物种共存机制[40]。WD能够反映植物的生长和防御能力,并在碳循环研究中起着关键作用[3,41—43]。在本研究中,WD与叶性状之间的相关关系很弱,仅与LPC呈正相关,而热带萨王纳树种及小笠原群岛上抗旱物种的WD与SLA显著负相关;在中国西南热带干旱喀斯特森林中,WD与比叶质量呈正相关[5];在色季拉山急尖长苞冷杉群落中,WD与SLA、LNC、LPC、LKC均呈显著正相关[44]。WD与叶性状的关系呈现出不同规律,可能是由于在不同的环境条件下,不同植物类群的WD与叶性状之间的适应策略不一致造成的。
SLA表示单位干生物量的光捕获表面积,与植物的资源竞争、环境耐受性及生活史策略相关[45],LDMC在一定程度上代表了植物对环境资源的利用能力,指示叶片建成投入的多少[46]。本研究发现SLA和LDMC是极显著的负相关关系,这一规律在大量的研究中得到验证[4,6,27,46—47]。此外,我们还发现SLA与LV呈极显著正相关关系,这与王超等[9]在急尖长苞冷杉林中发现的规律一致,而与张秀芳等[48]在柠檬桉(Eucalyptuscitriodora)和朴树(Celtissinensis)中发现的规律相反。这说明植物各性状间的权衡方式并不是一成不变的,植物会根据特定的环境条件采取不同的权衡策略,出现不同的性状组合,更好地适应环境,如:叶面积较小而LDMC较高的性状组合更能适应喀斯特生境[49];SLA较小而叶厚度、LDMC、叶组织密度较大的性状组合能够有效应对台风灾害频繁、土壤贫瘠的恶劣环境[48];叶绿素含量较高而LDMC较低的性状组合能够更好地利用生境中优越的光照资源[50]。本研究发现藏东木本植物功能性状形成了小SLA、LV,大LDMC的性状组合,在这样的性状组合下,植物的叶片小而厚,能够有效降低水分和养分的流失,叶片抗逆性更强,更有利于适应高寒环境。
叶片中的氮、磷和钾是协同元素,是植物体内许多重要有机化合物的组分,并以多种方式参与植物体内的各种代谢过程[51],它们之间一般呈正相关关系[47]。在本研究中,仅LPC与LKC之间存在显著正相关关系,LNC与LPC、LKC没有显著的相关关系,这与我国南方草地优势灌木巴柳(Salixetosia)、川莓(Rubussetchuenensis)[52]及摩天岭北坡草本植物叶片LNC、LPC的关系一致[53],而与胡耀升等[54]、任书杰等[55]、贺合亮等[51]的研究结果不同。植物对于氮、磷、钾元素的吸收会因物种的不同而产生特异性变化,并且各环境因子的波动也会对植物体内化学计量的积累产生影响。研究区内海拔跨度大,随着海拔的变化,群落优势物种和温度、降水、土壤养分等环境因子也会发生改变,影响土壤中氮、磷、钾元素的分布格局,植物根据立地环境的不同,对自身的养分吸收策略做出调整,导致叶片中LNC、LPC、LKC的协同方式出现差异。综上,藏东木本植物功能性状表现出适应高寒环境的权衡和性状间的协同变化,符合本研究的第一个科学假设。
3.2 群落功能性状对环境变量的响应
植物的功能性状决定了其在特定环境中的适应性,反映了植物对温度、湿度、土壤养分等环境因素的响应[56]。土壤是植物群落生长的物质和能量来源[47],在本研究中,土壤养分含量对LV产生了显著的正向影响,即LV随着土壤养分含量(PC1)的增加而增大(图3)。LV能够反映植物对资源和水分的获取能力,代表了植物的综合投资[57],LV的增大更利于植物获取和存储土壤中的养分资源。但更多的研究是分析了LV与单个土壤理化因子之间的关系,如:土壤可溶性有机碳对LV起正向作用[9];碱解氮与LV呈负相关,有效磷与LV呈正相关等[32]。
在本研究区内,海拔普遍较高,并且跨度较大(3143 —4628 m),海拔的变化会间接影响温度、降水、光照等因子产生再分配,导致气候环境的空间异质性,影响植物的生长发育并对植物功能性状的变化产生间接作用[32]。在本研究中,SLA随海拔的升高而降低,这与洪涛等[58]、齐威等[10]、王超等[46]的研究结论相同,而与Craine等[59]、Zhang等[60]相反。SLA小的植物更多见于高海拔、光照少、土壤贫瘠、干旱等胁迫性生境中[45,61],植物将更多的光合产物用于叶片防御组织的构建[9]。本研究还发现LV和海拔之间呈负相关关系,而色季拉山急尖长苞冷杉(Abiesgeorgeivar.smithii)LV与海拔之间不存在显著的线性变化趋势[9];军峰山长柄双花木(Disanthuscercidifoliusvar.longipes)为了更好的抢占资源,LV在中低海拔呈增大趋势[57]。不同地区研究结果的差异可能是因为不同山地海拔范围不同,气候、水分条件和物种组成也各不相同,导致群落功能性状对海拔的响应规律不一致。在藏东昌都地区,随着海拔的升高、气温的降低和年降水的增多,SLA、LV呈减小趋势,根据植物经济谱[4,62],植物叶片倾向于采取“高投入—慢回报” 的保守型策略来应对高海拔、低温的生长环境。
WD可作为木质经济谱的代表[3],WD低的植物生长速率更快,投入的机械支撑成本更低,枝干断裂和病原体侵入的风险更大,更易受到干旱胁迫[63—65]。已有研究表明WD高的物种更多生长在光照少、土壤贫瘠、干旱等胁迫性环境中[7,41,64]。而在藏东地区,随着海拔的升高(气温降低)和年降水量的增加,WD呈减小趋势,这可能是由于随着海拔升高,植被逐渐以低矮灌木为主,植物采取降低机械支撑成本、增加枝条御寒组织投入(如雪层杜鹃高(30—)60—90(—120)cm,小枝具有黑锈色鳞片[66])的策略,以提高其在严寒环境中的适合度;另外,高海拔地区生长季短,植物营养器官中的物质积累缓慢,较低的WD有助于其植株的快速生长和形态建成。综上,藏东木本植物功能性状对海拔和气候变量的响应最为显著,并且主要倾向于采取“高投入-慢回报”提高御寒能力的保守型策略来适应高海拔环境,支持并补充了第二个科学假设,即海拔和气候变量共同驱动着该区域植物功能性状的变化。
3.3 种内与种间变异在群落功能性状随环境变化中的作用
植物功能性状变异体现了植物对环境变化的适应能力差异;不同的物种对环境的适应能力不同,致使群落内的物种组成随环境变化而改变,造成功能性状的种间变异[67]。通过对群落加权性状的变异来源进行分解,我们发现:藏东植物群落功能性状对环境变化的响应主要由种间变异解释。这一结果与向响等[13]、Standen等[17]、Chelli等[27]的研究结论一致。研究区内的海拔从3143 m跨越到4628 m,环境过滤作用强烈,植物为了更好地适应环境,需要改变共存物种的竞争平衡,并且可能需要用其他物种替代部分或所有物种。本研究发现主要群落类型从较低海拔的砂生槐灌丛、川滇高山栎灌丛、钝叶栒子灌丛替代为较高海拔的雪层杜鹃灌丛、窄叶鲜卑花灌丛,物种出现了明显的更替。
但是,Hu等[28]发现在氮沉降的影响下,叶片的氮、磷含量及氮磷比主要由种内变异解释;Guo等[68]分别对比了CWM在施氮和干旱条件下的变异来源,发现在干旱条件下,植物的性状变异主要来自于种内变异。目前,已有大量的研究表明,种内变异的重要性不可忽视[7,26]。尽管种内变异对群落功能性状变异的直接影响很小,但它与种间变异的正或负协变产生的间接影响是巨大的[25]。藏东植物群落功能性状的种内变异主要通过与种间变异的负协变,间接对群落功能性状变异产生影响。
本研究发现海拔对群落功能性状变异的解释率最高,是影响群落功能性状变异最显著的环境变量。这与Lajoie等[69]在加拿大蒙特国家公园中发现的结论一致,同时,他们还发现在非气候因子(尤其是冠层间隙)中,种内变异贡献较大;Dwyer等[70]发现SLA的种内变异在土壤氮含量和土壤pH中解释率较高;Lepš等[33]发现种内变异在性状随土壤肥力的变化中发挥着重要作用。由此可见,植物功能性状种间、种内变异的相对贡献因环境变量而异。种间变异通常在环境变化强烈的区域中发挥显著作用,而在环境变化强度小的区域中,种内变异的重要性更加突出[33]。综上,海拔是影响藏东植物功能性状变异最显著的环境变量,种间变异在藏东植物群落功能性状随环境变化中起主要作用,符合本研究的第三个科学假设。种内变异主要通过与种间变异的负协变,间接对群落功能性状变异产生影响。
4 结论
藏东木本植物群落功能性状间的相关性很强,LPC和LKC具有协同性,但WD与叶性状的相关性低;植物在长期适应山地气候环境过程中,形成了相互权衡的性状组合(即:小SLA、LV,大LDMC)。海拔和气候变量(PC2)对于群落功能性状的影响最大,对WD、SLA、LV、LKC均存在显著的负向影响;并且LV还随着土壤养分含量(PC1)的增加而增大。我们还发现,藏东高海拔植物倾向于采取保守型策略适应高寒环境,种间变异在藏东植物功能性状随环境变化中起主要作用。本研究结论为高海拔植物功能性状对环境的适应机制提供了基础资料,对藏东地区生态系统功能的进一步研究和生物多样性保护具有重要意义。
猜你喜欢
军事文摘(2022年14期)2022-08-26
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
中国农业科学(2022年10期)2022-06-28
广东蚕业(2021年12期)2022-01-18
散文诗(2021年22期)2022-01-12
电脑爱好者(2020年10期)2020-07-28
电脑爱好者(2019年16期)2019-10-30
热带农业科学(2019年8期)2019-09-19
环球时报(2019-05-23)2019-05-23
青岛科技大学学报(社会科学版)(2015年4期)2016-01-25
