
草鱼朗格汉斯细胞的组织分布及其特征基因CD207的功能分析
2023-02-15 06:22朱曜良吴志新陈孝煊
淡水渔业 2023年1期
朱曜良,田 丁,吴志新,2,陈孝煊,2
(1.华中农业大学水产学院,武汉 430070;2.湖北省水生动物病害防控工程技术研究中心,武汉 430070)
1868年,LANGERHANS[1]在人的皮肤中发现了一种常驻的树枝状形态的细胞并将其命名为朗格汉斯细胞(Langerhans cells,LCs),朗格汉斯细胞是最早发现的一类树突状细胞(dendritic cell,DCs)。LCs特异性分布于哺乳动物的皮肤和其它黏膜组织中,在机体抗病原感染过程中作为专职抗原提呈细胞,摄取、加工处理并递呈抗原给T淋巴细胞,诱导相应的免疫反应[2]。LCs的直接前体细胞是一种高表达髓系分化抗原Gr-1分子的单核细胞,且集落刺激因子1受体(colony-stimulating factor-1 receptor,CSF-1R)、生长转化因子β(transforming growth factor-β,TGF-β)、粒细胞-巨噬细胞集落刺激因子(granulocyte-macrophage colony stimulating factor,GM-CSF)等一系列可溶性细胞因子在LCs的迁移、发育过程中发挥关键作用[3-5]。LCs的表面标志物主要由主要组织相容性复合体Ⅱ(major histocompatibility complex Ⅱ,MHC-Ⅱ),CD1a,E-钙黏蛋白(E-cadherin)和Langerin/CD207组成[6]。LCs对抗原的摄取加工需要表达Langerin/CD207,一种参与伯贝克颗粒(Birbeck granule)形成的C型凝集素[7]。CD207是Ⅱ型跨膜糖蛋白,为LCs特征性膜表面分子标记,因此CD207的表达也被广泛应用于朗格汉斯细胞的鉴定[8]。
关于哺乳动物LCs的研究已较为深入,而在硬骨鱼类中,鲜有关于LCs的研究报道。近年来,由于缺乏鱼类特异性CD207抗体,人源CD207多克隆抗体被普遍应用于硬骨鱼类LCs的鉴定。LOVY等[9]在孢子虫感染的大鳞大麻哈鱼(Oncorhynchustshawytscha)的鳃中发现了LCs。随后,在安大略鳟(Salmosalar)、虹鳟(O.mykiss)、斑点叉尾鮰(Ictaluruspunctatus)、美洲红点鲑(Salvelinusfontinalis)以及多种辐鳍亚纲(Actinopterygii)鱼类的头肾及脾脏中也发现了LCs[10-13]。最近,WANG等[14-15]利用透射电镜和人源CD207多克隆抗体在黄颡鱼(Pelteobagrusfulvidraco)脾脏、头肾和草鱼脾脏中分别观察到了LCs及其特征性标志物-伯贝克颗粒(Birbeckgranule),证明了黄颡鱼和草鱼体内朗格汉斯细胞的存在。
本实验室在前期的工作中制备了草鱼CD207多克隆抗体,可以特异性识别组织和细胞水平的内源性CD207蛋白[16]。目前草鱼体内LCs的分布及其特征性基因CD207的功能尚未阐明,本研究利用草鱼CD207多克隆抗体对LCs在草鱼相关免疫组织的分布情况进行鉴定,结合qPCR技术对CD207在感染中的表达情况进行分析,从而探讨LCs在草鱼抗病原免疫过程中的功能,为后续深入研究草鱼LCs及其它树突状细胞亚群的生物学功能提供基础资料。
1 材料与方法
1.1 实验材料
1.1.1 实验用鱼
实验草鱼(Ctenopharyngodonidella)于2021年9月购自湖北省黄冈市团风县百容水产良种有限公司。选取每尾体质量(200~300)g的健康草鱼,在循环水系统中暂养3周(华中农业大学水产学院水产养殖教学实验中心),水温(26±2)℃。期间按体质量的4%投喂商业饲料(湖北海大集团),饲料营养成分主要包括32%粗蛋白,13%粗灰分,1.6%赖氨酸,4%粗脂肪。每日更换1/3的水并清洁残余的饲料残渣。
1.1.2 实验试剂
LPS(E.coli055:B5,L6529)、PolyI:C(P9582)、Percoll分离液购自Sigma公司,HRP标记山羊抗兔IgG(H+L)、Alexa Fluor 555标记山羊抗兔IgG(H+L)购自武汉爱博泰克公司,DAPI染色液购自上海碧云天生物技术有限公司,FastPure Cell/Tissue Total RNA Isolation Kit V2、HiScript Ⅱ 1st Strand cDNA Synthesis Kit(+gDNA wiper)、ChamQ Universal SYBR qPCR Master Mix购自南京诺唯赞生物科技股份有限公司。
利用大肠杆菌表达系统表达草鱼CD207的胞外区片段作为抗原免疫新西兰大白兔(Oryctolaguscuniculus),制备草鱼CD207多克隆抗体。酶联免疫吸附剂测定(ELISA)其效价为1∶128 000,可识别组织和细胞水平分别对应的内源性蛋白,特异性较高[16]。
AIM液:含有47.5 mL L-15培养基,2.5 mL双抗,2.5 μg/mL两性霉素B和25 μg/mL庆大霉素,置于4 ℃备用。
细胞原代培养基:含有44.5 mL L-15培养基,0.5 mL双抗,5 mL胎牛血清(FBS),2.5 μg/mL两性霉素B和25 μg/mL庆大霉素,置于4 ℃备用。
1.1.3 引物列表
从NCBI GenBank数据库中获取CD207(GenBank:GBKA01017435.1)、β-actin(GenBank:M25013)基因的核苷酸序列,利用Primer 5.0软件分别设计CD207、β-actin的特异性引物。引物序列见表1,引物由武汉擎科生物技术有限公司合成。
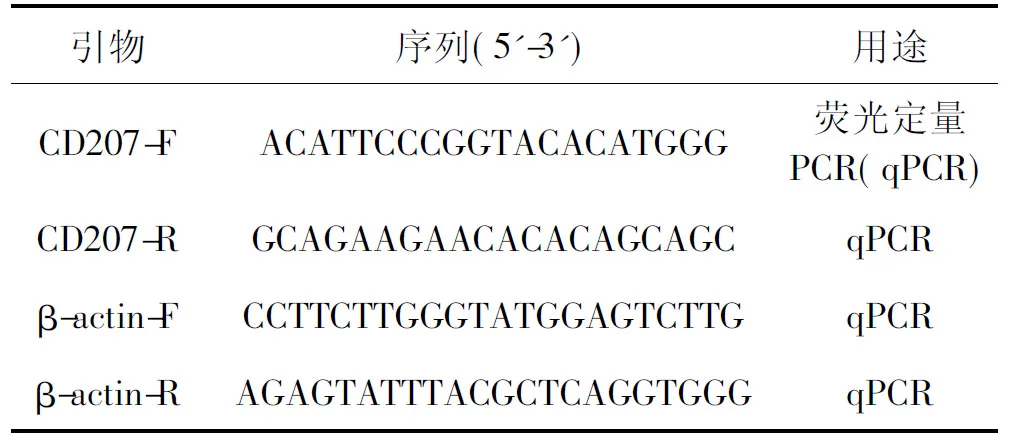
表1 引物碱基序列Tab.1 Nucleotide sequences of primers
1.2 免疫荧光
(1)取材固定:分别取草鱼头肾、鳃、皮肤、肠道的新鲜组织块,避免牵拉、挫伤与挤压,组织体积不超过1 mm×1 mm×1 mm,迅速投入4%多聚甲醛固定液固定24 h。将组织从固定液中取出,在通风橱中用手术刀修平整,放入脱水盒内。
(2)脱水、透明、浸蜡:采用从低浓度到高浓度逐级升高的梯度乙醇对组织块进行脱水,然后浸入二甲苯中将组织块透明,脱水透明后的组织块在熔化的石蜡包埋剂中浸蜡。
(3)包埋、切片、脱蜡至水:将完成浸蜡的组织块放入包埋框内包埋,于-20 ℃冻台冷却,将预冷的蜡块固定在石蜡切片机上切4 μm厚度切片,将切片在摊片机的温水(45 ℃)上摊平,放置65 ℃烤片机上烤1 h,再放置烘箱内烘烤2 h,之后经二甲苯脱蜡、梯度乙醇复水,最后蒸馏水洗。
(4)抗原修复、画圈、血清封闭:切片置于盛满乙二胺四乙酸(EDTA)抗原修复缓冲液(pH 8.0)的修复盒中于微波炉内进行抗原修复,自然冷却后将玻片置于PBS(pH 7.4)中在脱色摇床上晃动洗涤3次,每次5 min,切片稍甩干后用组化笔在组织周围画圈,在圈内滴加牛血清白蛋白(BSA)孵育30 min。
(5)加兔抗草鱼CD207一抗:轻轻甩掉封闭液,在切片上滴加1∶200(体积比)稀释的一抗,切片平放于湿盒内4 ℃孵育过夜。
(6)加Alexa Fluor 555 Goat Anti-Rabbit IgG(H+L):玻片置于PBS(pH 7.4)中在脱色摇床上晃动洗涤3次,每次5 min。切片稍甩干后在圈内滴加1∶200(体积比)稀释的二抗,避光室温孵育50 min。
(7)DAPI复染细胞核:切片稍甩干后在圈内滴加DAPI染液,避光室温孵育10 min。
(8)自发荧光淬灭:切片稍甩干后,在圈内加入自发荧光淬灭剂5 min,流水冲洗10 min。
(9)封片:玻片置于PBS(pH7.4)中在脱色摇床上晃动洗涤3次,每次5 min,切片稍甩干后用抗荧光淬灭封片剂封片。
(10)镜检拍照:切片置于扫描仪下采集图像或荧光显微镜下拍照。(DAPI紫外激发波长330~380 nm,发射波长420 nm,发蓝光;Alexa Fluor 555激发波长510~560,发射波长590 nm,发红光;DAPI染细胞核在紫外的激发下为蓝色,阳性表达为相应荧光素标记的红光)。
1.3 草鱼头肾白细胞的分离和处理
在无菌条件下取出草鱼头肾,AIM液中浸泡,用无菌注射器橡胶塞轻轻按压通过200目(70 μm)细胞筛。向15 mL离心管中依次加入51% Percoll分离液、34% Percoll分离液、过细胞筛的单细胞悬液,800g室温离心20 min后,吸取界面层之间的“白膜层”细胞,无菌PBS洗涤两次,加入细胞原代培养基重悬并调整密度为1×106cells/mL备用。
参考WANG等[17]和SU等[18]的方法,实验组白细胞分别加入终浓度为100 μg/mL的LPS和终浓度为50 μg/mL的PolyI:C,对照组白细胞加入等量无菌PBS溶液,处理12 h,每组设置三个重复。
1.4 草鱼CD207基因的表达分析
采用相对定量的方法对草鱼CD207基因经不同抗原处理后的表达量进行测定。分别离心收集步骤1.3处理所得细胞,草鱼头肾白细胞总RNA的提取和反转录,以及荧光定量PCR(qPCR)分别参照对应试剂盒说明书进行。qPCR采用20 μL反应体系:SYBR qPCR Master Mix(2×)10 μL,上下游引物(10 μmol/L)各0.5 μL,cDNA模板1 μL,ddH2O 8 μL。扩增条件:94 ℃预变性5 min,94 ℃变性15 s,60 ℃退火30 s,72 ℃延伸30 s,共40个循环。以2-ΔΔCt方法计算各组相对表达量,β-actin为内参基因,并进行显著性差异分析。“*”表示与对照组差异显著(P<0.05)。
1.5 草鱼CD207蛋白的表达分析
将步骤1.3处理后所得细胞样品分别与SDS-PAGE蛋白上样缓冲液(5×)在金属浴中100 ℃处理10 min,进行SDS-PAGE凝胶电泳和蛋白质印迹检测,采用半干法,将凝胶上的蛋白样品转移至PVDF膜上,首先加入5% 脱脂奶粉封闭2 h,然后用相应抗体(CD207多抗和β-actin内参抗体)与PVDF膜室温孵育1 h,TBST缓冲液洗膜15 min,重复4次;最后用HRP标记山羊抗兔IgG(H+L)二抗室温孵育50 min;TBST缓冲液洗膜10 min,重复3次后将ECL化学发光液覆盖在膜上,置于化学发光成像仪中成像。ImageJ软件(National Institutes of Health)进行灰度分析,并进行显著性差异分析。“*”表示与对照组差异显著(P<0.05)。
1.6 数据处理
所有数据采用平均值±标准误差表示,P<0.05时为统计学差异显著。采用GraphPad Prism 8.0软件绘图,Adobe Illustrator 2020修图。采用SPSS 24.0软件对实验数据进行统计学分析。
2 结果
2.1 朗格汉斯细胞在草鱼相关免疫组织分布情况
通过免疫荧光技术,在草鱼的头肾(图1a),鳃的鳃小片(图1c),皮肤的表皮及真皮(图1e)和肠道固有层(图1h)中均发现CD207阳性细胞(红色)的存在。免疫荧光显示LCs具有肾形或椭圆形的细胞核,头肾中LCs的胞体呈不规则形状突起,而鳃、皮肤、肠道中LCs的胞体则呈椭圆形。此外,LCs的细胞膜和细胞质中均有CD207蛋白的分布。
2.2 LPS、PolyI:C分别处理对CD207基因表达的影响
qPCR的结果显示,在处理12 h后,LPS(图2a)、PolyI:C(图2b)均能够诱导草鱼CD207基因相对表达量发生显著上调。
2.3 LPS、PolyI:C分别处理对CD207蛋白表达的影响
蛋白质印迹的结果显示,LPS(图3a)、PolyI:C(图3b)均能够诱导CD207蛋白表达量增多。ImageJ进行灰度分析,显示CD207蛋白相对表达量均发生显著上调,β-actin为内参蛋白。
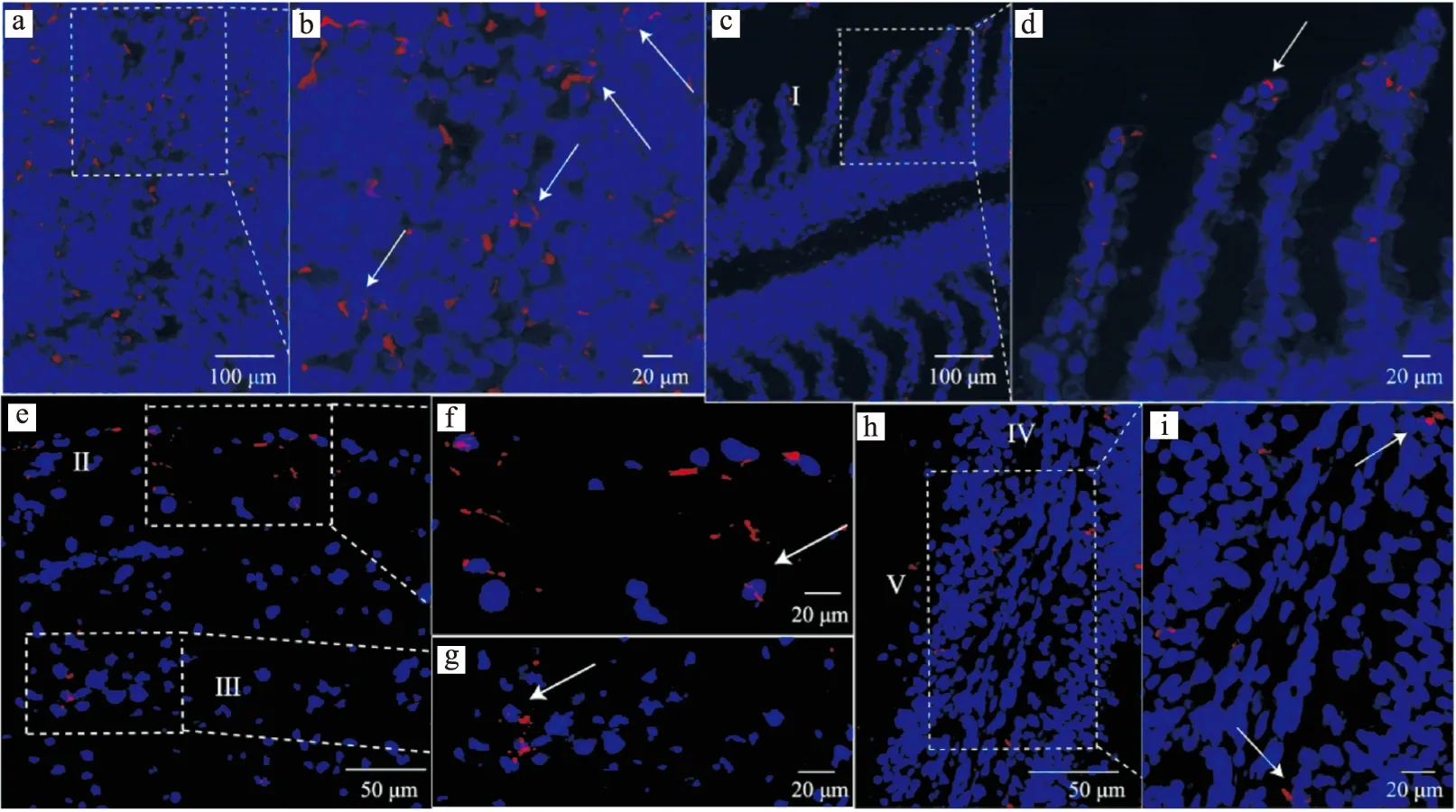
图1 草鱼头肾、鳃、皮肤和肠道中的CD207阳性细胞Fig.1 CD207 positive cells in head kidney,gill,skin and intestine of C.idellaa~b:头肾;c~d:鳃;e~g:皮肤;h~i:肠道;I:鳃小片;Ⅱ:表皮;Ⅲ:真皮;Ⅳ:固有层;Ⅴ:肠腔。图中箭头所示为CD207阳性细胞。

图2 实时荧光定量PCR检测LPS、PolyI:C对草鱼CD207基因相对表达量的影响Fig.2 Effects of LPS and PolyI:C on the relative mRNA expression of CD207 gene in C.idella detected by qPCR“*”为差异显著(P<0.05)。下同。
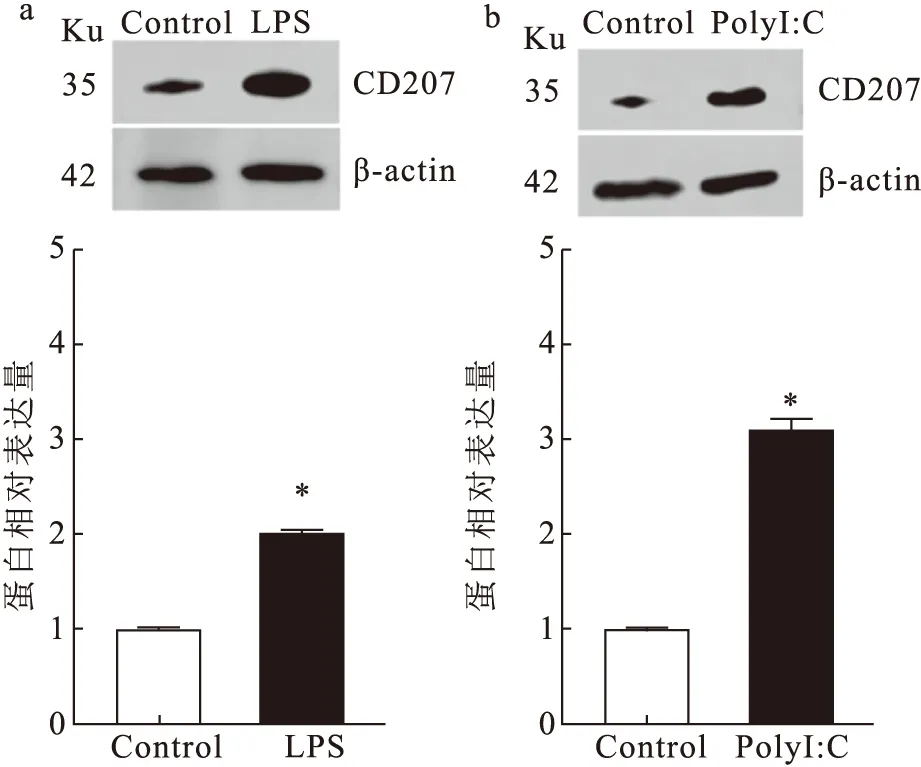
图3 LPS、PolyI:C对草鱼CD207蛋白相对表达量的影响Fig.3 Effects of LPS and PolyI:C on relative expression of CD207 protein in C.idella
3 讨论
3.1 草鱼朗格汉斯细胞在相关免疫组织的分布情况
树突状细胞在连接微生物病原体入侵机体所产生的先天性免疫和适应性免疫反应中起关键性桥梁作用,且树突状细胞各个亚群的形态、表面表型分子、组织分布和生物学功能也不尽相同[19]。LCs作为主要分布于黏膜免疫组织的树突状细胞亚群,已成为病原体通过黏膜入侵机体和维持机体稳态的研究热点[20]。头肾作为硬骨鱼类所独有的重要免疫器官,其中分布有大量的白细胞(包括单核细胞)[21],而LCs也由单核细胞分化而来,免疫荧光显示LCs在草鱼的头肾中分布,这提示头肾可能是LCs的发生场所之一。
皮肤、鳃和肠道组成了硬骨鱼类黏膜相关淋巴组织(mucosal associated lymph tissue,MALT)。这些组织富含黏膜层,与外部水环境直接接触,构成了鱼体免疫系统的第一道免疫防线[22]。鱼类皮肤中包含大量的黏液细胞和各类免疫细胞,与多种免疫相关因子形成了抵御病原微生物感染的“额外器官”[23]。免疫荧光定位LCs在存在于草鱼皮肤的表皮和真皮中,表明LCs是鱼类皮肤免疫系统的组成部分,也为后续深入解析鱼类皮肤免疫的生态网络结构提供基础资料。和皮肤黏膜类似,鱼类的鳃也是与水环境相互作用密切的黏膜器官。鳃中分布的各类免疫细胞,可以在病原体入侵机体时参与抗原摄入和激活免疫应答反应[24]。免疫荧光显示LCs分布于草鱼鳃的鳃小片中,提示草鱼患鳃部疾病时,LCs能够识别呈递抗原,参与草鱼机体的免疫监视。硬骨鱼类缺乏肠道相关淋巴组织(gut associated lymphoid tissues,GALT),如派尔集合淋巴结(Peyer patch)等,但是肠道固有层和上皮中含有大量的免疫细胞,包括淋巴细胞、颗粒细胞和肥大细胞等[25]。免疫荧光显示LCs分布于草鱼肠道固有层,这说明当颗粒状或大分子抗原入侵草鱼肠道时,来自肠腔的抗原经由肠上皮层包裹处理,后传递到肠道固有层,通过LCs的识别、加工和呈递给淋巴细胞,产生相应的免疫应答。
开展鱼类黏膜免疫中的抗原摄取、递呈和后续免疫反应的研究,完善黏膜免疫的应答机制,可以改善渔用疫苗的实验效果[26]。本实验鉴定了LCs在草鱼相关免疫组织的分布情况,丰富了草鱼树突状细胞亚群的研究内容,同时为黏膜疫苗的研发以及水产疫苗浸泡免疫的研究和推广等提供基础理论资料。
3.2 草鱼朗格汉斯细胞特征性基因CD207的功能分析
LCs在系统早期的细菌感染起到关键作用[27],CD207能识别病原体表面碳水化合物结构(如甘露糖、岩藻糖和N-乙酰葡萄糖胺),将抗原肽与MHC分子结合形成抗原肽-MHC复合体,从而呈递抗原参与适应性免疫应答[28]。VANDER等[29]发现人LCs能通过CD207摄取麻疹病毒(measles virus,MV)抗原并呈递给特异性CD4+T细胞以维持人体稳态。VANDEN等[30]也证明CD207促进人类免疫缺陷病毒(human immunodeficiency virus,HIV)转移并刺激LCs及部分树突状细胞亚群成熟,从而诱导CD8+T细胞的增殖活化。这表明LCs可以通过表达其特征性分子标记物CD207,在摄取、加工处理、呈递抗原进而引发特异性T淋巴细胞的产生,抗细菌、病毒感染方面都有不可忽视的作用。
原代培养细胞仍然保留部分甚至全部同在体细胞一样的生理功能特性,细胞仍然保持高度的分化性以及功能性,因此原代培养细胞被认为可以取代动物活体开展部分生命科学研究[31]。而且免疫刺激硬骨鱼的细胞模型后,各种免疫基因表达水平的快速反应也已被多方证实[32]。在培养鱼类原代细胞过程中,使用L-15原代细胞培养基(含10% FBS)同时维持培养基pH为7.0~7.4,并放置于28 ℃细胞培养箱,从而保持细胞在离体条件下的生理功能完整性,也避免了因细胞生长代谢受到抑制而引起的胞内mRNA的降解[33]。
免疫荧光显示LCs在草鱼头肾中分布,为排除正常体内自体调节和应激反应可能产生的整体因素影响,本实验对草鱼头肾白细胞进行分离培养,为天然免疫基因CD207的功能分析提供了可行工具。利用LPS、PolyI:C分别处理草鱼头肾白细胞,结果显示CD207的mRNA和蛋白的相对表达量均有显著性上调(P<0.05)。这表明在细菌或病毒入侵机体时,草鱼LCs可以通过上调CD207的表达,识别并呈递外源性抗原,参与到草鱼的适应性免疫调节反应中,提示草鱼LCs的特征性基因CD207表现出与哺乳动物CD207分子同系物相似的免疫功能。对草鱼CD207分子的研究,将有助于我们更好地了解LCs在硬骨鱼类免疫系统中的作用,为养殖过程中细菌性、病毒性疾病的防控提供理论基础。
4 结论
综上,本实验利用免疫荧光鉴定出LCs在草鱼的头肾,皮肤的表皮和真皮,肠道固有层和鳃的鳃小片中分布。运用蛋白质印迹法和qPCR对LPS、PolyI:C处理后草鱼头肾白细胞中的CD207表达情况进行分析,发现CD207的mRNA和蛋白的相对表达量均有显著性上调。结果表明,LCs分布于草鱼相关免疫组织中,特别是黏膜相关淋巴组织,可以通过表达CD207参与到草鱼抗细菌、病毒等外源性抗原的免疫反应中去。
猜你喜欢
当代水产(2022年7期)2022-09-20
当代水产(2022年3期)2022-04-26
广东第二课堂·小学(2018年9期)2018-10-24
电信科学(2016年11期)2016-11-23
中国组织化学与细胞化学杂志(2016年3期)2016-02-27
西南医科大学学报(2015年1期)2015-08-22
电子与信息学报(2015年6期)2015-07-12
医学研究杂志(2015年6期)2015-07-01
中国当代医药(2015年17期)2015-03-01
癌变·畸变·突变(2015年3期)2015-02-27
