
NtCPS2 基因对烟草腺毛发育及其分泌物合成的影响
2023-04-22 02:04韩文龙曹可心张路阳徐晓雯祖琼瑶徐世晓
河南农业科学 2023年3期
韩文龙,郑 聪,曹可心,张路阳,徐晓雯,谢 可,祖琼瑶,徐世晓
(1.河南农业大学 烟草学院,河南 郑州 450002;2.福建南平烟草公司,福建 南平 353000;3.河南省烟草公司许昌市公司,河南 许昌 461000)
烟草(Nicotiana tomentosiformisL.)腺毛是存在于烟草叶片上的一种特殊的多细胞结构,能够合成和分泌烟草重要的香气物质,对烟草的抗逆性和质量具有非常重要的作用[1]。烟草叶片的腺毛包括长柄腺毛、短柄腺毛以及保护毛[2]。其中,长柄腺毛和短柄腺毛合成烟草重要的致香物质西柏三烯二醇[3]。研究表明,烟草腺毛密度与烟草叶片中α-和β-西柏三烯二醇的含量呈正相关[4]。因此,通过调控烟草腺毛发育有助于提高和改善烟叶香气。
烟草腺毛的发育受多个基因调控,WANG 等[5]研究发现,敲除烟草NtCycB2(Cyclin B2)基因促进烟草长柄腺毛的形成,过表达NtCycB2基因抑制烟草长柄腺毛的生成;刘一华[6]研究发现,NtGIS(Glabeous inflorescence stems)基因过表达株系腺毛密度高于野生型(Wild type,WT),NtGIS基因干扰株系腺毛密度显著低于WT;崔丽鹏[7]研究发现,NtJAZ3(Jasmonate-zim-domain protein 3)基因负调控烟草多细胞腺毛的发育。另外,烟草腺毛的发育也受植物激素的调控[8-12]。娄亚楠等[10]和冯琦等[11]研究发现,外施茉莉酸甲酯可以提高烟草叶片腺毛密度;贺凌霄等[12]研究发现,外施赤霉素(GA3)可以提高烟草叶片腺毛的长度和密度。
NtCPS2基因是赖柏当类物质顺-冷杉醇(Cisabieion)的关键合成基因,在烟草腺毛中特异性表达[13]。贺凌霄等[14]研究发现,NtCPS2基因编辑后代GA3合成相关基因表达量提高,GA3含量增加。顺-冷杉醇和GA3都是烟草重要的二萜类物质[15],两者的代谢途径存在共同的合成底物香叶基香叶基焦磷 酸(Geranylgeranyl-PP,GGPP)[16]。目 前,关 于NtCPS2基因的研究多集中于其对顺-冷杉醇合成途径的影响方面,而NtCPS2基因与烟草腺毛发育及其分泌物合成的关系尚不明确。为此,以编辑NtCPS2基因的8306 纯合编辑系(KD)和WT 为材料[17],分别将KD 与WT 放置在正常生长条件下以及外施GA3和矮壮素处理的生长条件下,比较KD 和WT的叶片腺毛密度、西柏三烯二醇含量、GA3含量以及相关调控基因表达量的差异,以期明确NtCPS2基因对烟草腺毛发育及西柏三烯二醇合成的影响,为调控烟草腺毛发育和香气物质的合成提供新思路。
1 材料和方法
1.1 试验材料
试验材料为烟草8306 野生型WT 和纯合编辑系KD。其中,KD 是在分析8306NtCPS2基因CDS(编码区序列)和基因组序列的基础上,在第2 外显子区域设计了2 个gRNA 靶位点序列,构建RNA 载体,经过农杆菌介导的遗传转化得到的在靶位点上成功发生突变[17]的第二代纯合编辑系。
1.2 试验设计
选用饱满的8306 WT 和KD 种子,用1%KMnO4溶液浸泡消毒30 min 后,用蒸馏水洗净烟草种子表面的KMnO4溶液并播种于方形培养皿中,待种子发芽后移栽至装满基质(pH 值5.5~7.0、腐植酸含量≥5.0%、有机质含量≥25.0%)的黑色方形培养盆(高10 cm、直径8 cm)中。然后放入28 ℃恒温光照培养箱(12 h 光照/12 h 黑暗、湿度60%)中培养,待烟苗四叶一心时分别喷施0.025 mmol/L GA3及矮壮素(A),对照(CK)喷施蒸馏水。
1.3 测定指标及方法
1.3.1 腺毛形态和密度 在喷施后30 d 时,各处理均选取3 株长势均匀的烟株,取生长状态相近的中部叶片3 片,用2 g/L 罗丹明B 溶液进行染色处理,染色后用VHX-600V 超景深显微镜(日本基恩士公司)观察烟草叶片腺毛形态,并统计腺毛密度,每个叶片统计3个视野的腺毛密度。
1.3.2 西柏三烯二醇和顺-冷杉醇含量 在喷施后20 d时,各处理均选取5株长势均匀的烟株,取中间部位叶片5 片,保证叶片面积一致,去除叶片主脉,置于500 mL 的二氯甲烷溶液中进行浸提。收集浸提液后加入内标物质(1 mL正-17-烷醇),用旋转蒸发仪蒸发浓缩至50 mL 后进行衍生化反应,用GCMS 检测法进行分析并定性,最后根据内标物质含量进行西柏三烯二醇和顺-冷杉醇含量的定量分析。
1.3.3 GA3含量 在喷施后20 d 时,各处理均选取长势均匀的WT和KD中间部位叶片混样0.5 g,迅速用液氮冷冻后,放置于-80 ℃超低温冰箱中保存待测,随后采用间接酶联免疫吸附法(ELISA)[18]进行GA3含量测定。
1.3.4 GGPP 含量 在喷施后20 d 时,各处理均选取长势均匀的WT和KD中间部位叶片混样0.5 g,迅速用液氮冷冻后,放置于-80 ℃超低温冰箱中保存,随后采用间接酶联免疫吸附法(ELISA)[18]进行GGPP含量测定。
1.3.5 基因表达量 在喷施后20 d 时,各处理均选取长势均匀的WT和KD中间部位叶片混样0.1 g,采用总RNA 提取试剂盒(索莱宝生物公司)提取样品叶片中总RNA,放置于-80 ℃超低温冰箱中保存。利用PCR 仪将待测RNA 逆转录成cDNA,储存在-20 ℃冰箱中备用。
对赤霉素合成基因CPS(Ent-copalyl diphosphate synthase)、GA20ox(GA20-oxidase),顺-冷杉醇合成基因NtCPS2,GGPP 合成基因DXR(1-deoxy-D-xylulose 5-phosphate reductoisomerase),西柏三烯二醇合成基因CYC(Cembratrien-ol synthetase)、CYP71D16,腺毛负调控基因NtCycB2进行表达量分析,引物序列(表1)采用Roche LCPDS2软件设计,并由上海捷瑞生物工程有限公司合成。实时荧光定量PCR(qRT-PCR)扩增采用诺唯赞生物公司的TaqPro Universal SYBR qPCR Master Mix。20 μL 体系:2×TaqPro Universal SYBR qPCR Master Mix 10 μL,cDNA 0.4 μL,水8.8 μL,10 μmol/L正、反向引物各0.4 μL。qRT-PCR 程序:95 ℃预变性30 s;95 ℃10 s、60 ℃30 s,40 个循环。采用2-ΔΔCt法对基因表达量进行计算。

表1 荧光定量PCR引物序列Tab.1 Sequences of primers for fluorescence quantitative PCR
1.4 数据处理与分析
试验数据采用Excel 2016 软件进行计算和作图,使用SPSS 20.0统计软件进行差异显著性分析。
2 结果与分析
2.1 不同处理WT和KD中顺-冷杉醇含量及其关键合成基因NtCPS2表达量分析
由图1 可知,在CK 组中KD 顺-冷杉醇含量及NtCPS2基因相对表达量均显著低于WT,说明在编辑NtCPS2基因后顺-冷杉醇含量显著降低。外施GA3后,WT的顺-冷杉醇含量和NtCPS2基因相对表达量均显著降低;外施矮壮素后,KD 和WT的顺-冷杉醇含量和NtCPS2基因相对表达量总体上均显著升高。说明GA3代谢途径和顺-冷杉醇生物合成之间存在一定的联系。
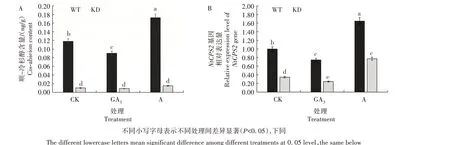
图1 不同处理KD和WT中顺-冷杉醇含量(A)及其关键合成基因NtCPS2相对表达量(B)Fig.1 The content of cis-abieion(A)and relative expression level of its key synthetic gene NtCPS2(B)in KD and WT under different treatments
2.2 不同处理WT和KD中GA3含量及其关键合成基因表达量分析
由图2A 可知,对于CK,与WT 相比,KD 中GA3含量升高25%;外施GA3后,KD 和WT 中GA3含量均显著增加,平均升高50%;外施矮壮素后,KD 和WT中GA3含量均显著降低,平均降低25%。由图2B—C可知,对于CK,KD中GA3关键合成基因CPS、GA20ox相对表达量均显著高于WT;外施GA3后,KD 和WT 中GA3关键合成基因CPS、GA20ox相对表达量均显著升高;外施矮壮素后,KD 和WT 中GA3关键合成基因CPS、GA20ox相对表达量均显著降低。整体上KD升高和下降的幅度大于WT。
2.3 不同处理WT和KD叶面腺毛形态和密度及其关键合成基因表达量分析
由图3A—B 可知,对于CK,KD 的叶片腺毛密度和长度均高于WT,平均升高28%,说明编辑NtCPS2基因后烟草叶片腺毛密度和长度增加;外施GA3后,KD 和WT 腺毛密度均显著增加,平均增加50%;外施矮壮素后,KD 和WT 腺毛密度均显著减少,平均减少56%。说明内源GA3含量的变化影响烟草叶片腺毛发育。图3C 表明,对于CK,编辑NtCPS2基因后KD 的NtCycB2基因相对表达量显著低于WT;外施GA3后,KD 和WT 的NtCycB2基因相对表达量均显著下降,KD 下降幅度大于WT;外施矮壮素后,KD 和WT 的NtCycB2基因相对表达量均显著升高,KD 升高幅度大于WT。此结果与叶片腺毛密度变化结果基本一致。
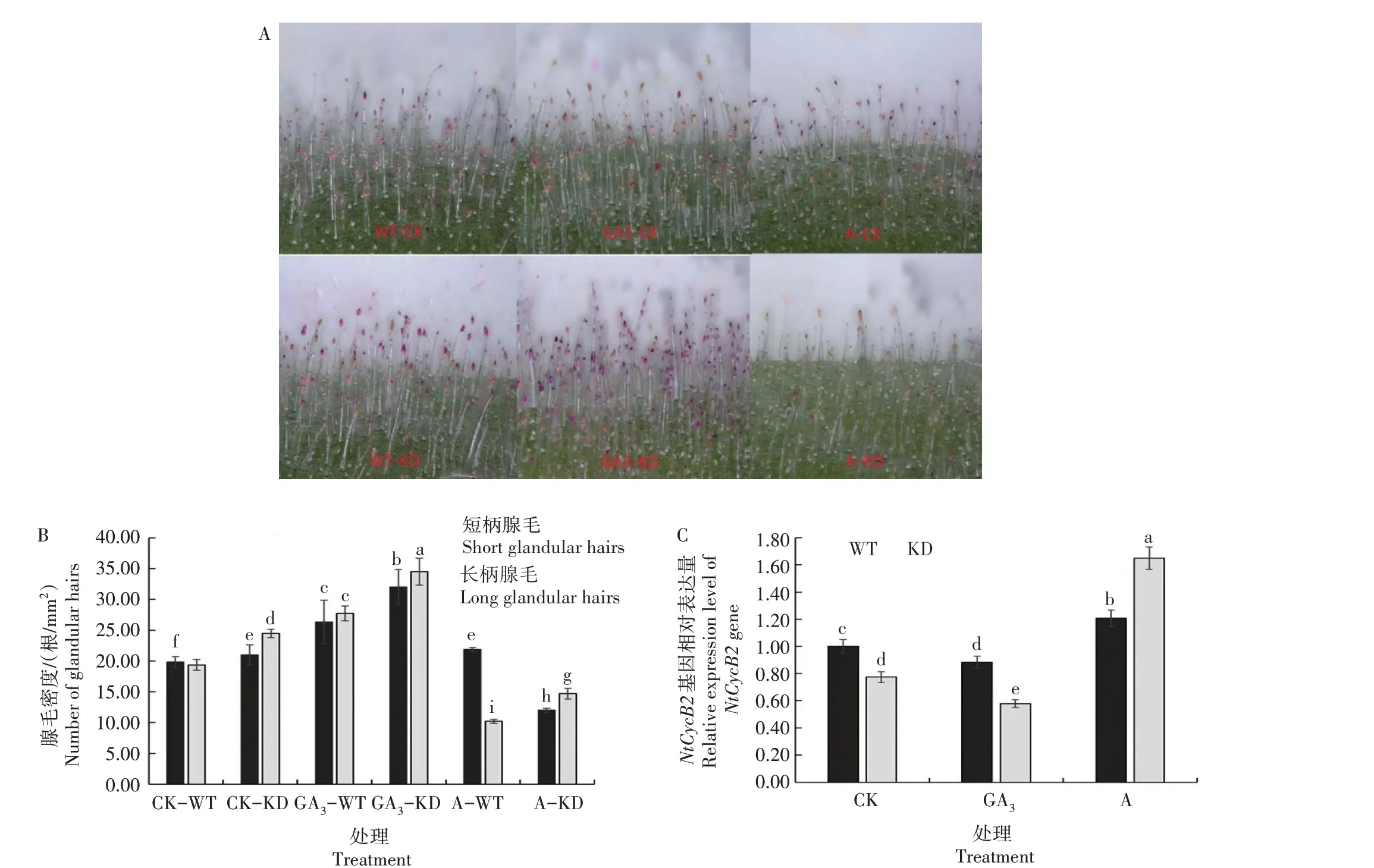
图3 不同处理KD和WT叶片腺毛形态(A)、密度(B)及其负调控基因NtCycB2相对表达量(C)Fig.3 The morphology(A)and number(B)of glandular hairs and the relative expression level of its negative regulator gene NtCycB2(C)in KD and WT under different treatments
2.4 不同处理WT和KD叶片腺毛西柏三烯二醇含量及其合成基因的表达量分析
图4A 表明,对于CK,KD 的叶片腺毛西柏三烯二醇含量高于WT,提高约25%,说明编辑NtCPS2基因提高了烟草叶片腺毛西柏三烯二醇分泌物含量;外施GA3后,KD 和WT 西柏三烯二醇含量均显著增加,平均增加50%;外施矮壮素后,KD 和WT 西柏三烯二醇含量均显著减少,平均减少46%。说明内源GA3含量的变化不仅影响烟草叶片腺毛发育,而且提高了烟草叶片西柏三烯二醇的分泌量。图4B—C 表明,对于CK,编辑NtCPS2基因后KD 的CYC和CYP71D16基因相对表达量均高于WT;外施GA3后,KD 和WT 的CYC和CYP71D16基因相对表达量均显著升高;外施矮壮素后,KD 和WT 的CYC和CYP71D16基因相对表达量均显著降低。整体上KD 升高和下降的幅度大于WT,这与西柏三烯二醇含量变化结果一致。
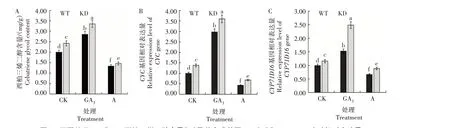
图4 不同处理WT和KD西柏三烯二醇含量(A)及其合成基因CYC(B)和CYP71D16(C)相对表达量Fig.4 The content of cembranoids(A)and relative expression level of its synthetic genes CYC(B)and CYP71D16(C)in WT and KD under different treatments
2.5 不同处理WT和KD中GGPP含量及其关键合成基因DXR表达量分析
由图5 可知,对于CK,KD 的GGPP 含量以及其关键合成基因DXR相对表达量均高于WT;外施GA3后,KD 和WT 的GGPP 含量和DXR基因相对表达量均显著升高;外施矮壮素后,KD 和WT的GGPP含量以及DXR基因相对表达量均显著下降。KD 的GGPP 含量以及DXR基因相对表达量升高或降低的幅度大于WT。
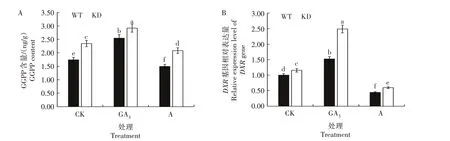
图5 不同处理WT和KD的GGPP含量(A)及其关键合成基因DXR相对表达量(B)Fig.5 GGPP content(A)and relative expression level of its key synthetic gene DXR(B)in WT and KD under different treatments
3 结论与讨论
本研究结果表明,与WT 相比,KD 的内源GA3含量升高。GA3和顺-冷杉醇生物合成途径存在共同底物GGPP,GGPP 在CPS 合酶的催化下参与GA3生物合成途径[19],在CPS2 合酶的催化下参与顺-冷杉醇合成途径[20]。NtCPS2基因编码CPS2,在编辑NtCPS2基因后,NtCPS2基因表达量下降,CPS2 活性降低,一方面减少了CPS2 对底物GGPP 的竞争,另一方面顺-冷杉醇合成途径受阻,间接导致底物GGPP 含量升高,更多的GGPP 参与GA3生物合成途径,内源GA3含量升高,这与贺凌霄等[14]的研究结果一致。
本研究结果表明,与WT 相比,KD 叶片腺毛密度增加,叶片腺毛分泌物西柏三烯二醇含量增加,同时KD 内源GA3含量升高,推测编辑NtCPS2基因影响了内源GA3的生物合成,进而影响了烟草腺毛的发育。因此,对KD 和WT 外施GA3和矮壮素,结果表明,与CK 相比,外施GA3后,KD 和WT 的内源GA3含量显著增加,叶片腺毛密度显著增加,NtCycB2基因表达量显著降低;外施矮壮素后,KD和WT 的内源GA3含量降低,叶片腺毛密度减少,NtCycB2基因表达量显著升高。说明内源GA3含量是影响烟草腺毛发育的因素之一。研究表明,GA3对拟南芥腺毛的生长和发育有重要的作用[21]。周忠静[22]研究发现,GA3的合成和运输对拟南芥腺毛的发育具有重要的调控作用。GA3信号传导影响腺毛生长相关关键基因的启动,研究表明,GL1(Glabrous 1)和TTG(Transparent testa glabra)基因是调控腺毛发育的2 个关键基因,其中GL1 蛋白序列与受GA3调 控 的 Myb(V-myb avian myeloblastosis viral oncogene homolog)家族转录因子同源,TTG基因对腺毛发育的调控作用可被与GA 信号传导有关的R(R1R2R3)基因所代替[23-26],烟草腺毛的发育调控机制与拟南芥存在一定的相似性,本研究发现,NtCPS2基因通过调控内源GA3的生物合成和信号转导进而影响烟草腺毛发育。刘一华[6]的研究结果表明,在烟草中外施GA3导致烟草叶片腺毛密度显著增加,而用VIGS 技术干扰GA3生物合成基因GA20ox后,干扰材料内源GA3含量降低,叶片腺毛密度显著降低,这与本研究结果一致。WANG 等[5]研究发现,烟草NtCycB2基因负调控烟草腺毛发育,过表达NtCycB2基因,烟草叶片腺毛密度降低,沉默NtCycB2基因,烟草叶片腺毛密度升高,这与本研究结果一致。
本研究发现,与WT 相比,KD 叶片腺毛分泌物西柏三烯二醇含量显著升高。西柏三烯二醇是烟草重要的香气物质,其主要由烟草叶片腺毛合成和分泌,一方面KD 叶片腺毛密度增加,西柏三烯二醇含量升高;另一方面,编辑NtCPS2基因后,西柏三烯二醇关键合成基因CYC、CYP71D16表达量升高,西柏三烯二醇含量增加。研究发现,烟叶表面腺毛密度与叶片腺毛分泌物含量呈正相关[27]。烟株内GA3含量升高,有助于烟草叶片腺毛分泌物西柏三烯醇的积累[28]。外施GA3提高了烟草叶片腺毛密度,外施矮壮素减少了叶片腺毛的密度[12]。此外,GA3也是二帖类化合物的一种[29],其与西柏三烯二醇具有共同底物GGPP,外施GA3提高了烟株内源GA3含量,进而节省了底物GGPP 在GA3合成途径中的消耗,提高了GGPP 含量,进而提高了西柏三烯二醇合成关键基因CYC、CYP71D16的表达量,西柏三烯二醇含量增加。外施矮壮素则降低了内源GA3含量,植物为了维持内源GA3的平衡,更多的GGPP 参与GA3合成,增加了GGPP 的消耗,进而抑制了西柏三烯二醇关键合成基因CYC、CYP71D16的表达,西柏三烯二醇含量减少[30-31]。
本研究发现,编辑NtCPS2基因后KD 内源GA3含量升高,叶片腺毛密度增加,西柏三烯二醇含量升高。此结果与外施GA3处理相同,与外施矮壮素处理相反。表明编辑烟草NtCPS2基因影响了烟草内源GA3的生物合成进而影响了烟草叶片腺毛的发育及其分泌物的合成,初步揭示了NtCPS2基因在烟草萜类代谢和腺毛发育中的作用。
猜你喜欢
奥秘(创新大赛)(2023年3期)2023-05-06
中国饲料(2022年5期)2022-04-26
陕西农业科学(2020年11期)2020-12-25
中国烟草学报(2018年2期)2018-05-09
浙江中西医结合杂志(2017年2期)2017-01-12
当代化工研究(2016年9期)2016-03-20
湖南农业科学(2015年3期)2015-10-09
邵阳学院学报(自然科学版)(2015年1期)2015-06-05
癌变·畸变·突变(2015年4期)2015-02-27
声屏世界(2014年6期)2014-02-28
