
连翘属种质资源抗寒性评价1)
2023-05-31 02:56徐洋赵峥畑吴雨桐申建双张启翔程堂仁王佳潘会堂
东北林业大学学报 2023年6期
徐洋 赵峥畑 吴雨桐 申建双 张启翔 程堂仁 王佳 潘会堂
(北京林业大学,北京,100083)
杨晓辉
(河北省平泉市黄土梁子镇人民政府)
连翘开花繁茂、花色金黄,是我国北方地区早春园林植物景观的“符号”性植物之一,在园林中广泛应用。连翘每年的生长期约为270~320 d,5月中下旬至7月中旬花芽分化,霜降时停止生长,次年早春气温达到6 ℃左右时花芽开始萌发,花期可持续40 d左右,盛花期15 d[1]。目前园林应用的主要种类包括连翘(Forsythiasuspensa)、金钟连翘(F.intermedia)和朝鲜连翘(F.koreana)的杂交品种,这些连翘品种花量大,适应性强,但一些品种的抗寒性较弱,在我国北方冬季严寒地区露地栽培不能正常越冬,成为影响其在北方地区广泛应用的重要因素[2]。因此,研究连翘的抗寒性,筛选出抗寒性较强的连翘种质,具有十分重要的意义。
目前,有关连翘抗寒性的鉴定方法及抗寒生理的研究不多。回彦哲等[3]对低温胁迫下连翘和金钟连翘枝条的相对电导率、超氧化物歧化酶(SOD)活性和可溶性蛋白质量分数进行测定,发现美国金钟连翘(F.intermedia‘Zabel’)的抗寒性强于连翘(F.suspensa)。李盼华等[4]以10种早春开花植物为试材,人工模拟低温处理,测定相对电导率,结合Logistic方程计算其半致死温度(LT50),试验结果表明,抗寒性最强的植物是迎春,其次是迎红杜鹃和连翘,上述3种植物的抗寒性强于其他的7种。袁金凤等[5]测定了在春季冻融交替时期几种灌木枝条在低温胁迫下的生理指标,综合评价抗寒性的排序由强到弱为小花溲疏、紫叶小檗、太平花、红王子锦带、迎春、连翘、金叶女贞、棣棠、金山绣线菊。
本研究在河北省平泉市露地栽植10个连翘属种与品种,通过测定露地越冬期间1年生枝条的生理生化指标变化和水培萌发的情况,以及越冬后调查植株的田间表现,评价连翘属植物的抗寒性。以期为连翘的抗寒性评价提供一种科学、正确、高效的鉴定方法,并筛选出抗寒育种亲本,为东北、内蒙古等冬季严寒地区连翘属植物的应用提供可靠依据。
1 试验区概况
试验地位于河北省平泉市黄土梁子国有林场,为河北省、辽宁省、内蒙古自治区三省区交界地(东经118°21′3″~119°15′34″,北纬40°24′~40°40′17″),年平均降水量为540.1 mm。
利用HOBO-U23温湿度记录仪测定距地面1.5 m高度处的温度和湿度,利用HOBO-U23土壤温度记录仪测定土壤深度20~30 cm处的温度,每1 h记录1次数据,记录时间从2019年10月20日—2020年4月20日。得到试验地气温、地温和空气相对湿度情况如图1~图3所示。1a中,试验地最低气温和地温出现在12月31日,分别为-32.14℃、-12.86℃。
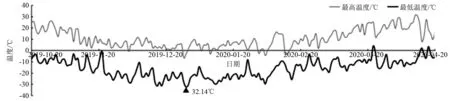
图1 试验区每天的最高气温和最低气温变化
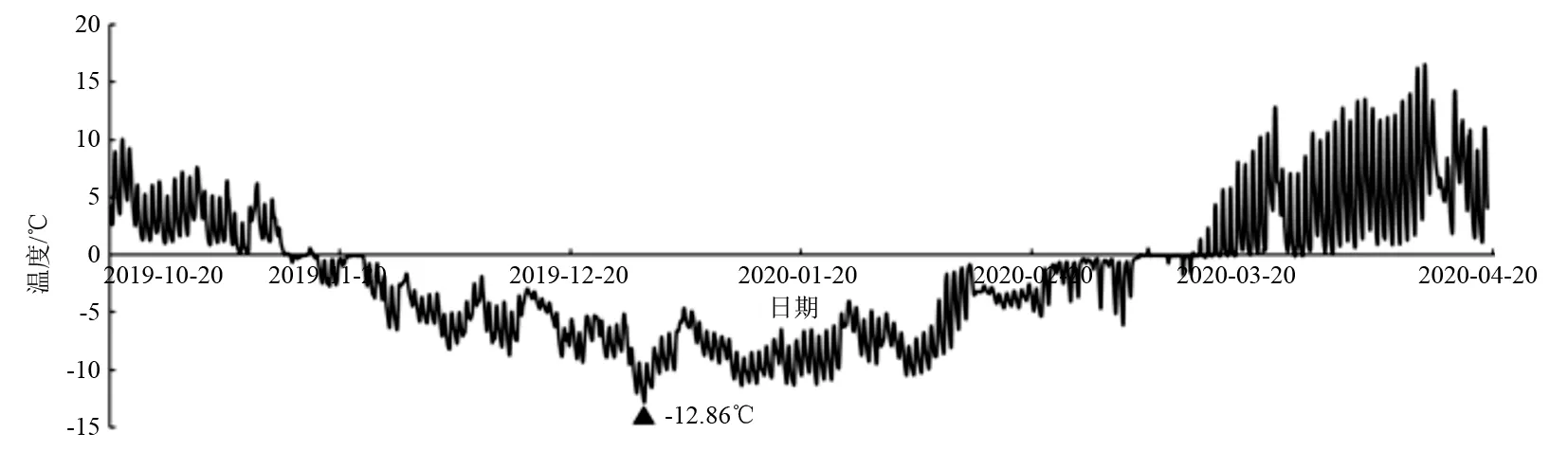
图2 试验区地温变化情况
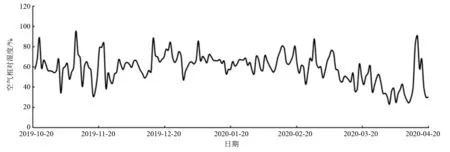
图3 试验区空气相对湿度变化情况
2 材料与方法
2013年5月在黄土梁子国有林场定植10个连翘种和品种的扦插苗(3个原生种,分别是连翘、东北连翘、朝鲜连翘,其他7个是人工培育品种),分别为:连翘(F.suspensa)、东北连翘(F.mandschurica)、朝鲜连翘(F.koreana)、‘Courtalyn’(F.intermedia‘Courtalyn’)、‘Courtaneur’(F.intermedia‘Courtaneur’)、‘Goldrausch’(F.intermedia‘Goldrausch’)、‘Spectabilis’(F.intermedia‘Spectabilis’)、‘Courtasol’(F.intermedia‘Courtasol’)、‘Lynwood’(F.intermedia‘Lynwood’)、‘Kumsum’(F.koreana‘Kumsum’)。株行距为1.5 m×1.5 m,每种或品种栽植20株。第1年在植株基部培土防寒越冬,从第2年起不再做任何防寒措施。
参照任丽等[6]的方法,分别于2019年10月20日、11月20日、12月20日、2020年1月20日在当年生休眠枝条顶部向下数第3节起剪取长15~20 cm的枝条,每种或品种剪取8~10个枝条,带回实验室后立刻测定相对电导率,其余枝条在液氮中研磨后,置于冰箱中-80 ℃保存备用。
2.1 测定指标及方法
相对电导率测定。参照田景花等[7]和刘强等[8]的方法,将田间采回的粗细一致的枝条切成长度为0.5 cm的小段,称取0.1 g装入15 mL的离心管中,加入5 mL的去离子水,摇床震荡24 h,采用DDS-307A电导仪测定初电导值(C1),将其放入沸水浴中30 min,冷却至室温后测定终电导值(C2)。每个连翘种或品种重复测定3次。计算公式为:相对电导率=(C1/C2)×100%。
采用蒽酮比色法测定连翘枝条的可溶性糖质量分数,采用考马斯亮蓝G-250染色法测定可溶性蛋白质量分数,采用氮蓝四唑法测定超氧化物歧化酶(SOD)活性,采用愈创木酚比色法测定过氧化物酶(POD)活性[9]。
越冬期间水培枝条萌芽观测。将不同时期从田间剪取的枝条(每种各3个枝条)插入自来水中,置于光照培养箱中,设置培养温度为25 ℃,湿度为60%,每天光照10 h,观测萌芽状况,直到不再有新萌发的侧芽为止。
田间越冬表现观测。参照李晓宇等[10]的方法,分别在2019年6月和2020年6月,统计连翘种质资源的干梢枝条数量占枝条总量的比例,以及枝条干梢长度占枝条总长度的比例。
2.2 隶属函数值计算方法
使用Excel2010软件和SPSS23软件对获得的生理生化指标的原始数据进行统计和分析,隶属函数值的计算参考王海鸥等[11]的方法。
与抗寒性呈正相关关系的参数指标公式为:
U(Xij)=(Xij-Xjmin)/(Xjmax-Xjmin);
与抗寒性呈负相关关系的参数指标公式为:
U(Xij)=1-(Xij-Xjmin)/(Xjmax-Xjmin)。
式中:U(Xij)为i品种j指标的抗寒性隶属函数值,Xij为某一指标的测定值;Xjmin为某一指标测定值中的最小值;Xjmax为某一指标测定值中的最大值。
3 结果与分析
3.1 越冬期间枝条的生理生化指标变化
从表1可以看出,越冬期间各品种连翘种质枝条的相对电导率均逐渐增加,在温度最低的1月份达到最大值。其中‘Courtalyn’、‘Goldrausch’、‘Spectabilis’、‘Lynwood’等4个品种枝条电导率增幅较大,1月20日相对电导率分别为95.12%、68.63%、94.62%、86.81%,与10月20日相比分别升高了77.13%、46.71%、73.90%、67.32%;其他种质1月份的相对电导率低于49.95%。东北连翘1月份的相对电导率为32.70%,显著低于其他参试种质(P<0.05);东北连翘的相对电导率最为稳定,1月份较10月份仅增加了8.96%。
从表1可以看出,多数连翘种和品种的可溶性糖质量分数表现出逐渐升高的趋势。‘Lynwood’和东北连翘的可溶性糖质量分数在越冬期间增幅较大,10月份的可溶性糖质量分数分别为2.26%、2.39%;1月份分别为5.75%、5.87%;增长了154.42%、145.61%。‘Goldraush’的可溶性糖质量分数在12月份达到峰值,1月份明显下降。连翘(F.suspensa)的可溶性糖质量分数在整个越冬期间的变化较为平缓。
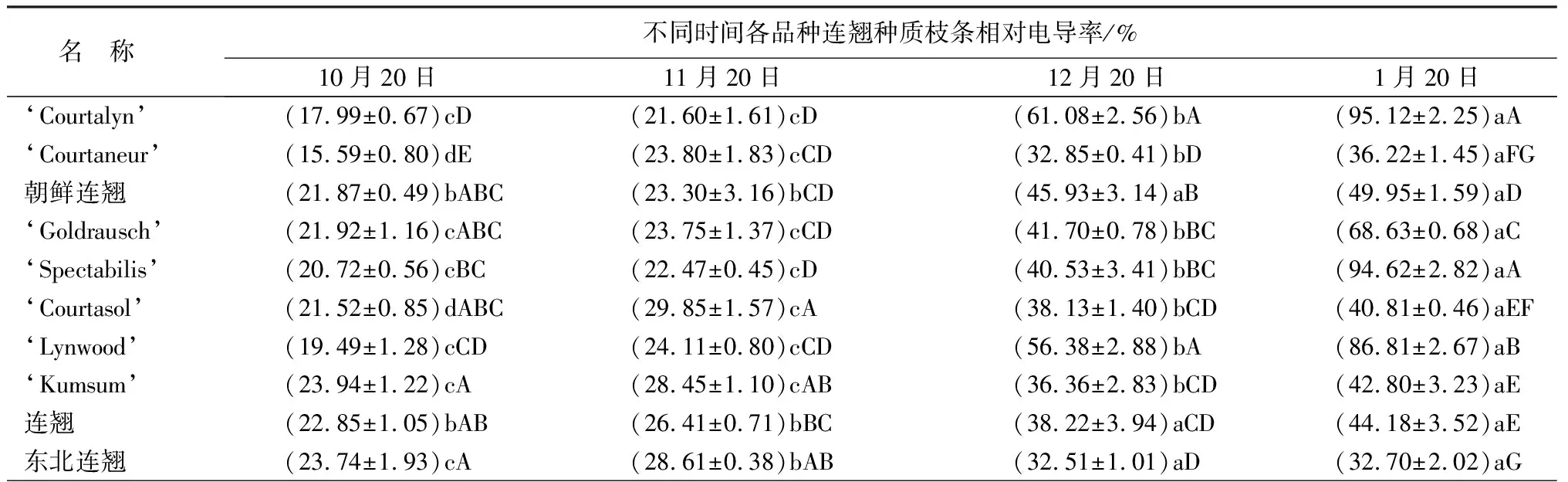
表1 越冬期间连翘种和品种枝条各测定指标的变化
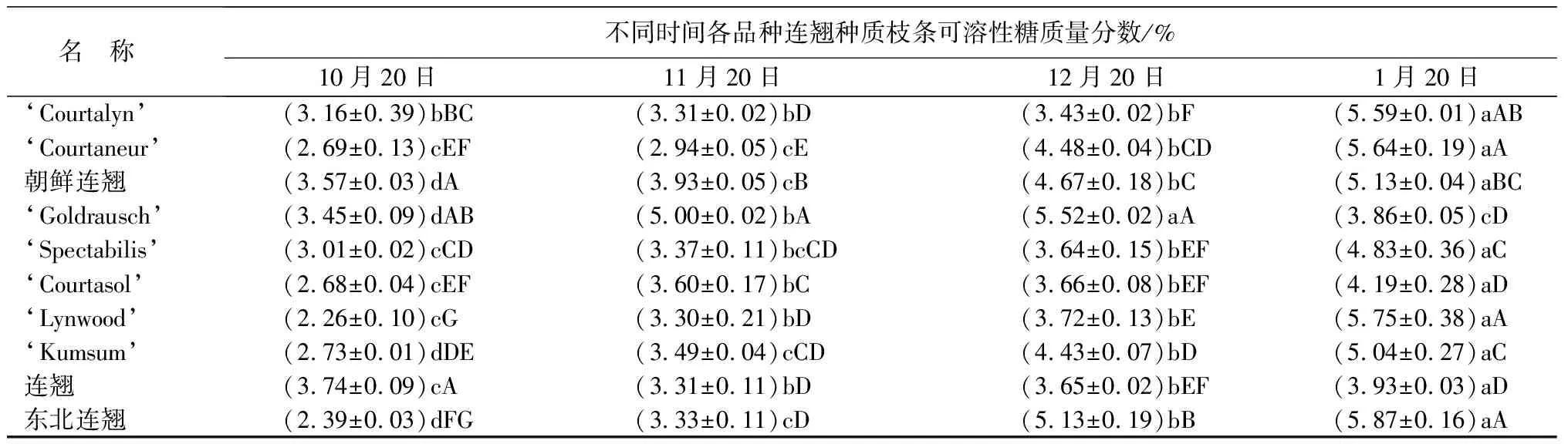
名 称不同时间各品种连翘种质枝条可溶性糖质量分数/%10月20日11月20日12月20日1月20日‘Courtalyn’(3.16±0.39)bBC(3.31±0.02)bD(3.43±0.02)bF(5.59±0.01)aAB‘Courtaneur’(2.69±0.13)cEF(2.94±0.05)cE(4.48±0.04)bCD(5.64±0.19)aA朝鲜连翘(3.57±0.03)dA(3.93±0.05)cB(4.67±0.18)bC(5.13±0.04)aBC‘Goldrausch’(3.45±0.09)dAB(5.00±0.02)bA(5.52±0.02)aA(3.86±0.05)cD‘Spectabilis’(3.01±0.02)cCD(3.37±0.11)bcCD(3.64±0.15)bEF(4.83±0.36)aC‘Courtasol’(2.68±0.04)cEF(3.60±0.17)bC(3.66±0.08)bEF(4.19±0.28)aD ‘Lynwood’(2.26±0.10)cG(3.30±0.21)bD(3.72±0.13)bE(5.75±0.38)aA‘Kumsum’(2.73±0.01)dDE(3.49±0.04)cCD(4.43±0.07)bD(5.04±0.27)aC连翘(3.74±0.09)cA(3.31±0.11)bD(3.65±0.02)bEF(3.93±0.03)aD东北连翘(2.39±0.03)dFG(3.33±0.11)cD(5.13±0.19)bB(5.87±0.16)aA
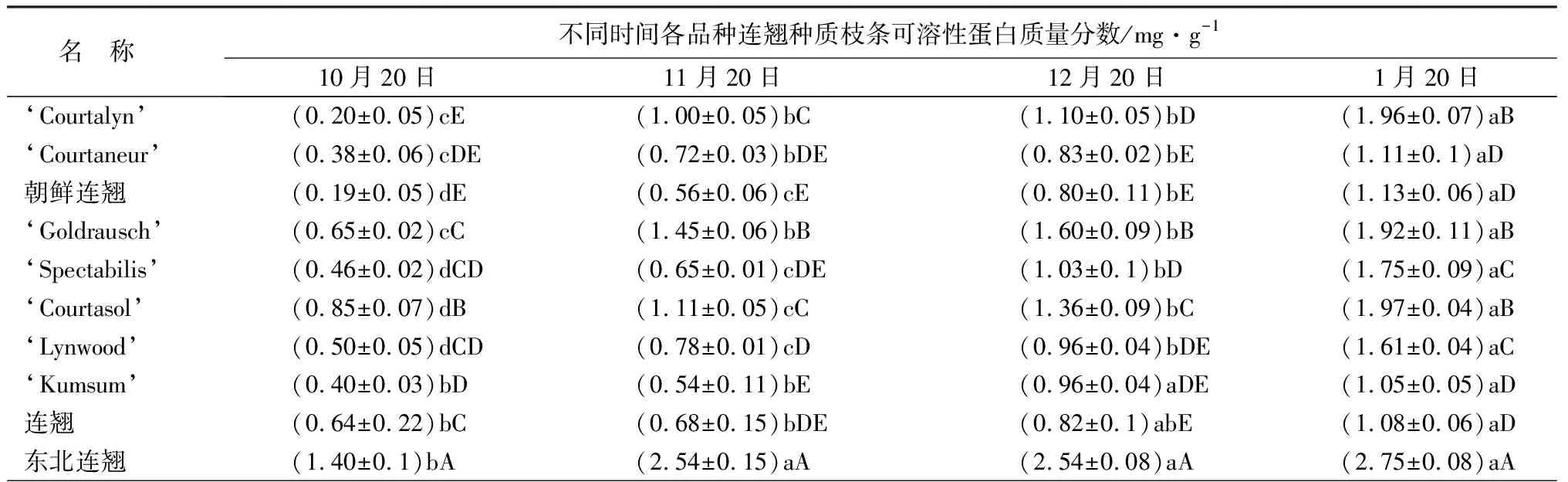
续(表1)
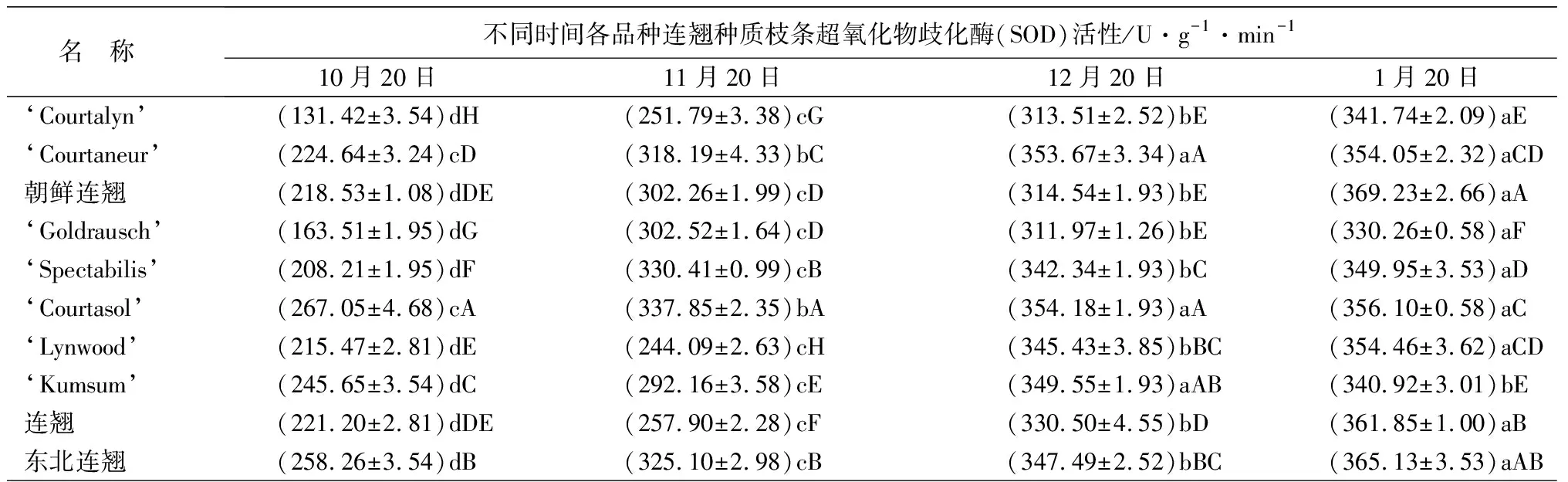
名 称不同时间各品种连翘种质枝条超氧化物歧化酶(SOD)活性/U·g-1·min-110月20日11月20日12月20日1月20日‘Courtalyn’(131.42±3.54)dH(251.79±3.38)cG(313.51±2.52)bE(341.74±2.09)aE‘Courtaneur’(224.64±3.24)cD(318.19±4.33)bC(353.67±3.34)aA(354.05±2.32)aCD朝鲜连翘(218.53±1.08)dDE(302.26±1.99)cD(314.54±1.93)bE(369.23±2.66)aA‘Goldrausch’(163.51±1.95)dG(302.52±1.64)cD(311.97±1.26)bE(330.26±0.58)aF‘Spectabilis’(208.21±1.95)dF(330.41±0.99)cB(342.34±1.93)bC(349.95±3.53)aD‘Courtasol’(267.05±4.68)cA(337.85±2.35)bA(354.18±1.93)aA(356.10±0.58)aC ‘Lynwood’(215.47±2.81)dE(244.09±2.63)cH(345.43±3.85)bBC(354.46±3.62)aCD‘Kumsum’(245.65±3.54)dC(292.16±3.58)cE(349.55±1.93)aAB(340.92±3.01)bE连翘(221.20±2.81)dDE(257.90±2.28)cF(330.50±4.55)bD(361.85±1.00)aB东北连翘(258.26±3.54)dB(325.10±2.98)cB(347.49±2.52)bBC(365.13±3.53)aAB
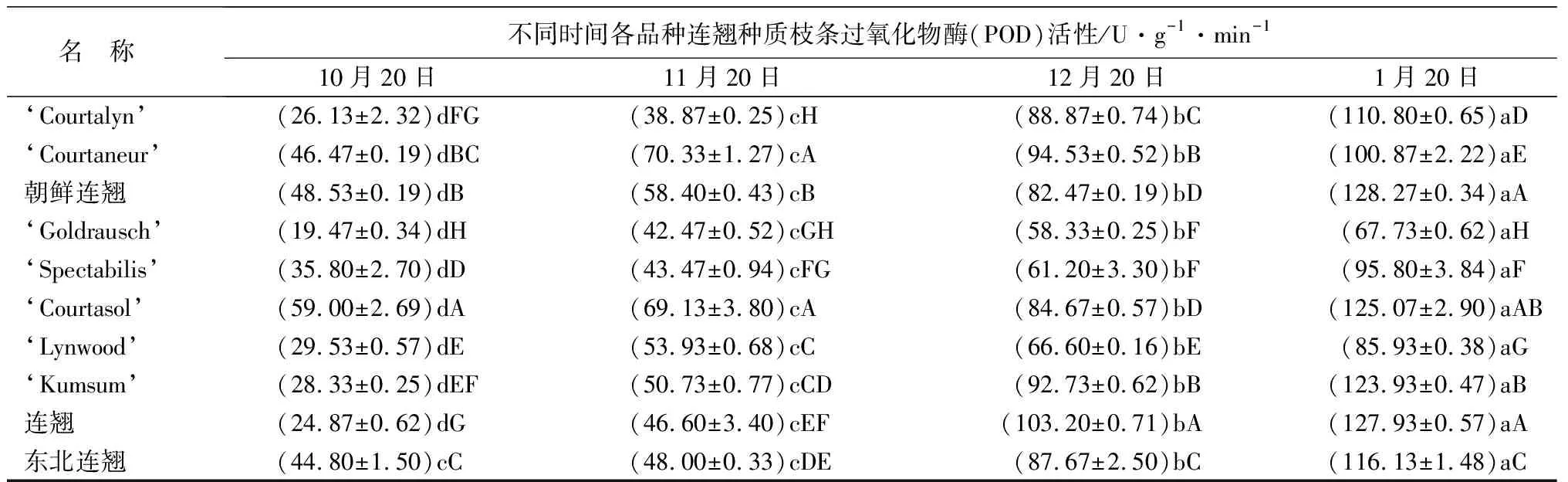
名 称不同时间各品种连翘种质枝条过氧化物酶(POD)活性/U·g-1·min-110月20日11月20日12月20日1月20日‘Courtalyn’(26.13±2.32)dFG(38.87±0.25)cH(88.87±0.74)bC(110.80±0.65)aD‘Courtaneur’(46.47±0.19)dBC(70.33±1.27)cA(94.53±0.52)bB(100.87±2.22)aE朝鲜连翘(48.53±0.19)dB(58.40±0.43)cB(82.47±0.19)bD(128.27±0.34)aA‘Goldrausch’(19.47±0.34)dH(42.47±0.52)cGH(58.33±0.25)bF(67.73±0.62)aH‘Spectabilis’(35.80±2.70)dD(43.47±0.94)cFG(61.20±3.30)bF(95.80±3.84)aF‘Courtasol’(59.00±2.69)dA(69.13±3.80)cA(84.67±0.57)bD(125.07±2.90)aAB ‘Lynwood’(29.53±0.57)dE(53.93±0.68)cC(66.60±0.16)bE(85.93±0.38)aG‘Kumsum’(28.33±0.25)dEF(50.73±0.77)cCD(92.73±0.62)bB(123.93±0.47)aB连翘(24.87±0.62)dG(46.60±3.40)cEF(103.20±0.71)bA(127.93±0.57)aA东北连翘(44.80±1.50)cC(48.00±0.33)cDE(87.67±2.50)bC(116.13±1.48)aC
越冬期间所有连翘种和品种的可溶性蛋白质量分数均持续上升,1月份达到最大值。东北连翘11月份的可溶性蛋白质量分数较10月份增幅较大,增长了1.14 mg/g,增幅为81.43%;在整个越冬期间东北连翘的可溶性蛋白质量分数都显著高于其他9份连翘种质;‘Courtaneur’、朝鲜连翘、‘Kumsum’和连翘(F.suspensa)在整个越冬期间的可溶性蛋白质量分数处于较低水平;其中连翘(F.suspensa)的可溶性蛋白质量分数在越冬期间的上升幅度最小,增幅为68.75%(表1)。
所有连翘种和品种的超氧化物歧化酶(SOD)活性均逐渐上升,不同取样时间之间大多存在显著性差异。‘Kumsum’的SOD最大活性出现在12月份,1月份有所下降。SOD活性在自然越冬期间的增速均呈由快到慢的趋势,‘Courtalyn’从10到11月份增加了91.59%,11到12月份增加了24.51%,12到下一年1月份仅增加了9.00%(表1)。
越冬期间各连翘种和品种枝条的过氧化物酶(POD)活性均持续上升,到1月份达到最大值,不同取样时间之间的变化幅度明显,均存在显著性差异。在整个越冬期间,‘Goldrausch’的POD活性整体表现偏低,大多时间显著低于其他种质(P<0.05)(表1)。
3.2 隶属函数法综合评价抗寒性
对10份连翘种质枝条生理生化指标的隶属函数值进行计算并求平均值,平均值越大的表明其抗寒性越强。由表2可得,10个连翘属种质的抗寒性由强到弱排序为:东北连翘、‘Courtaneur’、‘Courtasol’、朝鲜连翘、‘Kumsum’、连翘(F.suspensa)、‘Spectabilis’、‘Goldrausch’、‘Lynwood’、‘Courtalyn’。
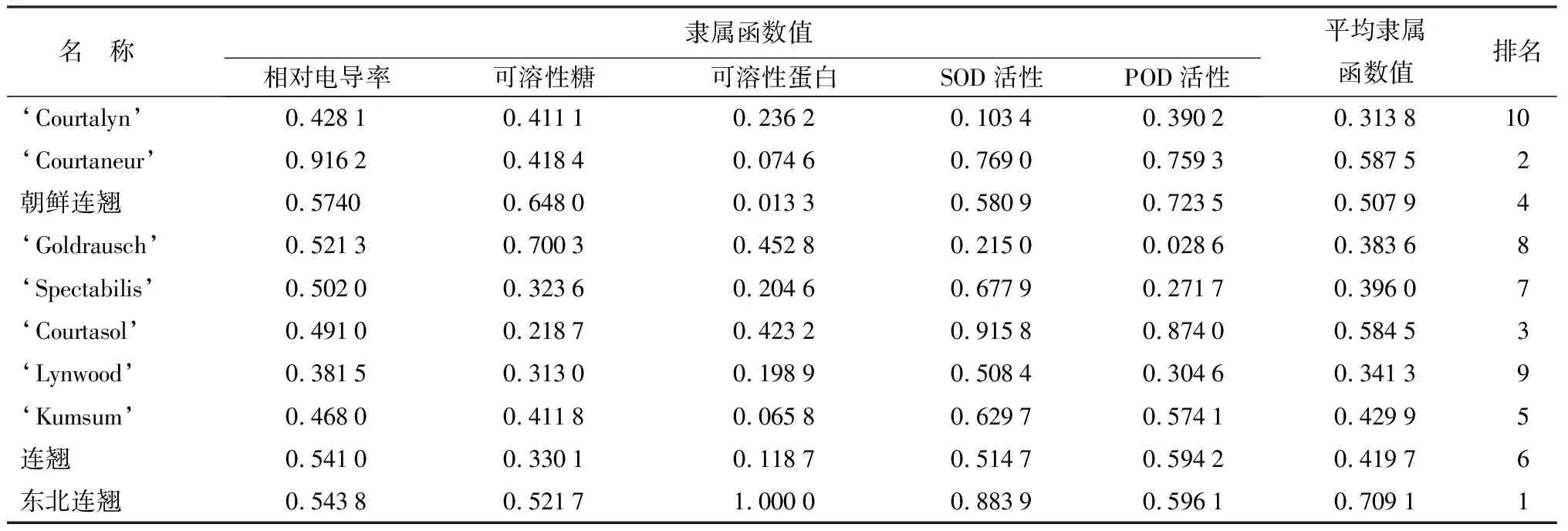
表2 连翘种和品种抗寒性的综合评价
3.3 枝条水培萌芽情况
各个时期枝条水培萌芽情况如图4所示。10月20日和11月20日剪取的枝条全部能够萌发;12月20日‘Courtalyn’、‘Lynwood’的枝条干枯,水培不再萌发;1月20日‘Goldrausch’和‘Spectabilis’枝条也已干枯,水培不再萌发,说明这些品种的抗寒性不足以抵抗此时的低温环境。以上4种干枯的连翘种质的抗寒性较弱,‘Courtalyn’、‘Lynwood’对低温的忍耐力低于‘Goldrausch’和‘Spectabilis’。其余6个连翘种和品种在4个时间段水培枝条均能正常萌芽。
3.4 连翘露地越冬后的生长表现
10份连翘种质露地自然越冬后田间调查指标情况如表3所示,生长状况如图5所示。越冬后,连翘(F.suspensa)、‘Lynwood’、‘Spectabilis’、‘Goldrausch’、‘Courtalyn’的地上部分全部干枯,即干梢枝条数量占总枝条数量的比例、干梢长度占枝条总长度的比例均为100%。‘Kumsum’、朝鲜连翘、‘Courtasol’干梢枝条数量比例在30%到75%,干梢长度比在10%到15%,基本不能在试验地露地条件下开花。‘Courtaneur’的干梢枝条数量比为5%,干梢长度比为20%,可少量开花。在10份材料中,只有东北连翘的地上部分越冬后完全不受影响,能够正常生长、开花。结果表明,东北连翘抗寒性最强,‘Courtaneur’次之,‘Kumsum’、朝鲜连翘、‘Courtasol’抗寒性较强,连翘(F.suspensa)、‘Lynwood’、‘Spectabilis’、‘Goldrausch’、‘Courtalyn’抗寒性较弱,这一结果与隶属函数法综合评价抗寒性得到的排序基本相符合。

从左向右依次为:Courtalyn’、‘Courtaneur’、朝鲜连翘、‘Goldrausch’、‘Spectabilis’、‘Courtasol’、‘Lynwood’、‘Kumsum’、连翘、东北连翘。

图5 10个连翘种和品种露地越冬后生长状况(2020年6月)
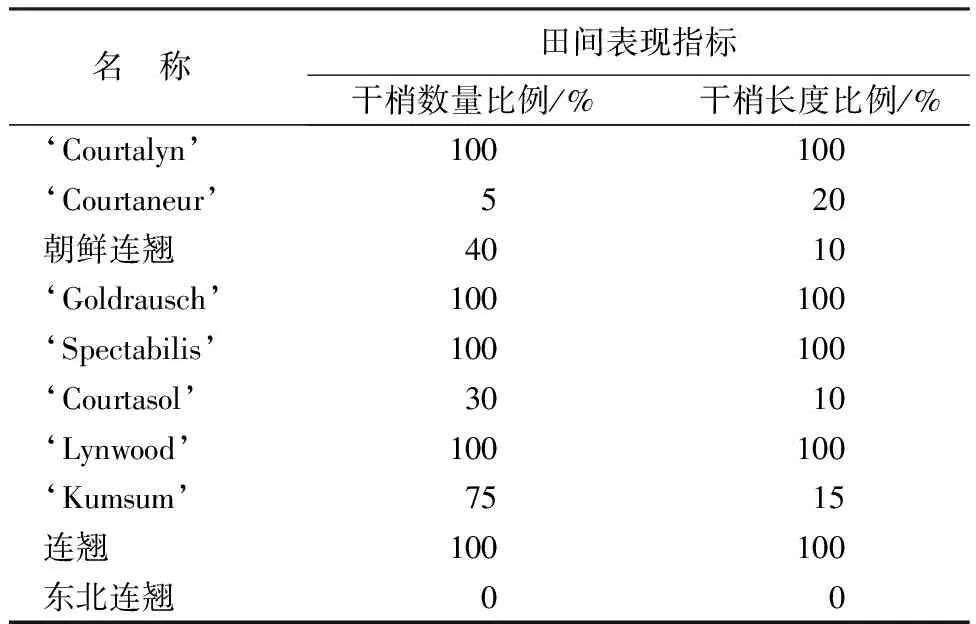
表3 露地自然越冬后田间表现指标
4 结论与讨论
自然越冬期间10个连翘属种和品种当年生枝条的相对电导率、可溶性糖质量分数、可溶性蛋白质量分数、SOD活性和POD活性整体上逐渐升高,利用隶属函数法进行综合抗寒性评价,得到的抗寒性由强到弱排序为:东北连翘、‘Courtaneur’、‘Courtasol’、朝鲜连翘、‘Kumsum’、连翘、‘Spectabilis’、‘Goldrausch’、‘Lynwood’、‘Courtalyn’。越冬后,10种连翘的田间表现调查结果和上述排序结果基本相符合。
低温胁迫会抑制植株正常的生长发育和繁衍,植物感受到了低温的侵害后,会尽快做出防御提升其耐寒性,在这个过程中存在大量的低温感受、生理反应、信号转导等调控途径[12-13]。Lyons[14]提出的著名的“膜脂相变”假说,即当植物体受到低温胁迫时,细胞膜会由液晶状态变成凝胶状态来保护自己。低温胁迫下,细胞膜透性变化比外部形态变化早且明显,因此细胞膜透性可作为植物低温胁迫下抗寒性评价的重要生理指标之一[15]。植物的抗寒性强,细胞膜透性增大的程度较慢,而抗寒性弱的植物则细胞膜透性增大的程度较快[16]。用相对电导率评价植物抗寒性已经广泛运用于核桃、蔷薇等植物中[17]。本研究表明,在自然越冬过程中连翘种质的枝条相对电导率均逐渐升高,抗寒性差的连翘品种相对电导率增长较快。东北连翘枝条的相对电导率最为稳定,1月份比10月份仅升高了8.96%,表明其细胞膜伤害程度轻,具有极强的抗寒性。
低温胁迫过程中,植物体内的淀粉分解为可溶性糖,可溶性糖质量分数的增加能够提高细胞液浓度,以降低冰点,维持植株的正常生长,因此可溶性糖的质量分数与植物的抗寒性呈正相关关系[18]。本研究中,10月份的测定值中,连翘(F.suspensa)的可溶性糖质量分数最高,但在随后的3个月中变化不大,因此未能在越冬过程中有效提高植物细胞抵抗低温的能力。‘Goldrausch’的可溶性糖质量分数1月份较12月份显著下降,可能由于此时的枝条已经干枯,植物细胞严重受损导致代谢紊乱,可溶性糖质量分数急剧下降。
低温条件下,可溶性蛋白质量分数的增加可以增加植物中束缚水的含量,以提高植物细胞的保水能力[19]。在本研究中,抗寒性强的东北连翘枝条的可溶性蛋白质量分数在10月20日到11月20日之间急剧上升了81.02%,之后维持在一个较高的水平;相反,连翘(F.suspensa)的可溶性蛋白质量分数一直偏低,且变化幅度不大,不足以提高保水能力及其抵抗低温的能力。‘Courtalyn’、‘Spectabilis’、‘Courtasol’和‘Lynwood’可溶性蛋白的生理响应较慢,到1月份时枝条的可溶性蛋白质量分数得到显著提升,而此时它们已经受到严重冻害,因此未能做到及时有效地应对低温伤害。
低温胁迫下,植物体内自由基增多,启动膜脂过氧化反应,破坏了植物细胞,诱发SOD、POD等保护酶活性上升,清除体内自由基,降低细胞膜通透性,保护细胞膜结构完整性,缓解低温伤害[20]。一定范围内,抗氧化酶活性会随温度的降低而增强,抗寒性强的品种的抗氧化酶活性上升幅度大于抗寒性弱的品种[21]。本研究也获得了类似的结果,在自然越冬过程中,连翘种质的SOD、POD活性均逐渐升高以抵抗低温伤害。其中,‘Goldrausch’的POD活性在整个越冬期间处于较低水准,因而表现出较弱的抗寒性。
植物在低温胁迫下发生组织破裂、失水萎蔫、器官褐化等症状,田间鉴定法是在自然越冬条件下,对生长的植物进行包括存活状况和外部形态的调查,进而由此判断植物的抗寒性强弱[22]。本研究中,在河北省平泉市,10个连翘属种和品种露地越冬后,只有东北连翘不存在干梢的现象,说明它可以普遍栽植应用。而其他几种连翘发生了不同程度的干梢现象,说明它们不适于在当地大面积应用。连翘(F.suspensa)枝条在1月20日仍能水培萌发,但越冬后的田间调查发现,地上部分全部干枯,可能存在两种情况:第一,在1月20日之后,再经过一段时间的低温积累,极大损伤了组织细胞导致枝条不再萌芽;第二,如图1、2、3试验地环境因子变化情况可知,3月下旬到4月上旬空气相对湿度降低,4月4日仅为23.17%,而这一时期的气温适合植株萌动,需水量较大,虽然地面已经解冻,但植物没有完全恢复吸水能力,枝条供水不及时,容易导致生理干旱,发生抽条现象[23]。
植物的生理过程是错综复杂的,受多种因素影响,孤立地用某一指标表示这一复杂的抗寒生理过程,不能真实反映植物的抗寒水平[24]。利用隶属函数法对多个指标进行综合评价,能克服只利用少数指标进行评价的不足,以揭示连翘属不同种和品种间对低温反应特性的实质,从而提高抗寒性鉴定的准确性。在本研究中,在露地自然越冬过程中,测定连翘属种和品种的多个生理生化指标,利用隶属函数法综合评价抗寒性,得到了抗寒性排序。越冬后调查得到的田间表现情况与上述排序结果基本保持一致,说明用隶属函数法评价连翘抗寒性具有科学性和可靠性。因此,利用隶属函数法综合评价抗寒性的方法可以作为评价连翘属植物抗寒性较为准确的方法。
猜你喜欢
今日农业(2021年21期)2021-11-26
中国民间疗法(2021年8期)2021-07-22
今日农业(2020年16期)2020-09-25
中成药(2018年10期)2018-10-26
现代园艺(2017年11期)2017-06-28
新疆农垦科技(2016年2期)2016-08-21
中国农业文摘-农业工程(2016年5期)2016-04-12
中国果菜(2016年9期)2016-03-01
现代农业(2016年4期)2016-02-28
中国马铃薯(2015年5期)2016-01-09
