
川西亚高山天然林岷江冷杉的更新结构与数量动态
2023-06-14 08:17杨玉婷李茂萍薛盼盼岳喜明
生态学报 2023年10期
陶 琼,缪 宁,杨玉婷,李茂萍,薛盼盼,岳喜明
四川大学生命科学学院,教育部生物资源与生态环境重点实验室,成都 610065
川西岷江上游的亚高山天然林是成都平原的重要水源地和生态屏障,在长江经济带发展和国家生态安全构建中具有重要作用[1]。20世纪50年代到1998年期间,川西亚高山林区内以冷杉(Abiesspp.)和云杉(Piceaspp.)为优势树种的原始暗针叶林被大面积采伐,随后在采伐迹地上发生了人工更新和天然更新[2]。1998年以来,林区相继实施了天然林资源保护、退耕还林等生态工程,全面实施了禁伐封育。经历近50年的自然恢复与人工更新,逐渐形成了不同林龄的人工林、天然次生的针阔混交林(如桦木-岷江冷杉林)、次生阔叶林(如桦木林)、人工林及天然林的混交林和残留的原始暗针叶林(如杜鹃-岷江冷杉林和箭竹-岷江冷杉林)镶嵌分布的景观格局[3—4]。岷江冷杉(Abiesfargesiivar.faxoniana)为该区原始暗针叶林的主要优势种,其天然更新是川西亚高山次生林自然恢复的关键[5]。
不同森林类型的立地条件以及群落结构差异使得岷江冷杉的更新状况或质量表现各异[5]。研究表明,在卧龙的华西箭竹(Fargesianitida)-岷江冷杉原始林中,岷江冷杉在幼树阶段的死亡密度最高[6];在桦木-岷江冷杉原始林中岷江冷杉的天然更新良好,种群大体呈增长型[7—8]。在王朗的岷江冷杉原始林中岷江冷杉幼苗个体丰富,种群结构为增长型[9];岷江冷杉老龄林中,岷江冷杉幼苗在基径0.7—0.8 cm时处于关键和敏感阶段[10]。在米亚罗林区对比藓类和箭竹岷江冷杉林型之间的天然更新发现,该区藓类林型的岷江冷杉天然更新优于箭竹林型[5]。以往对岷江冷杉的更新状况的研究多集中在原始林或者次生林中,对原始林与天然次生林间岷江冷杉更新状况的比较研究较少[5]。
以往对岷江冷杉更新的研究主要包括林窗更新[10—13]、箭竹(Fargesiaspp.)对其更新的影响[6,13—17]、幼苗根系生物量[18]、增温对幼苗生长的影响[19—23]、不同径级优势种对其幼龄个体空间分布的关联分析[24—27]、林线交错带内的更新动态[28]等。生命表与存活曲线分析可以反映其存活状态,结合结构动态量化分析可以进一步分析种群受干扰状况及其变化趋势[10],对岷江冷杉幼龄植株更新结构和数量动态的研究有利于揭示其存活瓶颈期和存活数量变化趋势。然而,以往关于不同林型之间岷江冷杉更新质量以及幼龄植株存活瓶颈期的对比研究则鲜见。
因此,本研究以川西亚高山4种典型天然林(杜鹃-岷江冷杉林、箭竹-岷江冷杉林、红桦-岷江冷杉林和红桦林)为研究对象,分析各林型中岷江冷杉的更新结构与数量动态,明确4种林型的更新质量以及制约岷江冷杉幼龄植株存活的瓶颈阶段,以期为探索川西亚高山不同林型天然林中促进岷江冷杉更新的经营管理技术提供参考和依据。
1 研究方法
1.1 研究区概况
研究区域位于川西阿坝州理县米亚罗林区(31°24′—31°55′ N,102°35′—103°4′ E),地处青藏高原向四川盆地过渡的高山峡谷区,海拔2200—5500 m,坡度多在30°以上。米亚罗林区的森林状况和经营历史,在整个川西林区具有典型性和代表性[29]。本区地处青藏高原气候区,冬寒夏凉。年均气温约9 ℃,年降水量700—1000 mm,年蒸发量1000—1900 mm[4]。米亚罗林区植被呈明显的垂直带状分布,亚高山暗针叶林是海拔2700—4000 m的原生林型,主要优势树种为岷江冷杉[30]。本研究中,红桦林与红桦-岷江冷杉林采伐前均为岷江冷杉原始林。箭竹-岷江冷杉林、红桦-岷江冷杉林及红桦林森林景观主要分布在米亚罗海拔2800—3600 m的阴坡、半阴坡,以及沟谷两旁山坡下部的阴湿生境中;杜鹃-岷江冷杉林森林景观主要分布在海拔3600 m以上阴坡和半阴坡的高寒生境中[3]。
1.2 样地选取与调查
在米亚罗林区的4种典型林型:杜鹃-岷江冷杉林(原始林,2018年调查)、箭竹-岷江冷杉林(原始林,2014年调查)、红桦-岷江冷杉林(次生针阔混交林,2005年调查)和红桦林(次生阔叶林,2005年调查)中各选择1块1 hm2样地进行调查(表1)。箭竹-岷江冷杉林样地内的乔木层郁闭度为0.67,优势种为岷江冷杉,伴生有红桦(Betulaalbosinensis)、云杉(Piceaasperata)等;灌木盖度为0.50,优势种为华西箭竹,伴生有陕甘花楸(Sorbuskoehneana)、华西蔷薇(Rosamoyesii)、栒子木(Cotoneastermultiflorus)等。杜鹃-岷江冷杉林样地内的乔木层郁闭度为0.47,优势种为岷江冷杉,伴生有红杉(Larixpotaninii)、理县杜鹃(Rhododendrontrichogynum)等,灌木盖度为0.06,优势种为红背杜鹃(Rhododendronrufescens),伴生有冰川茶藨子(Ribesglaciale)、唐古特忍冬(Loniceratangutica)等。红桦林样地内的乔木层郁闭度为0.60,优势种为红桦,伴生有岷江冷杉、中华槭(Acersinense)、川西云杉(Picealikiangensisvar.rubescens)等,灌木层盖度为0.35,主要有华西蔷薇、冰川茶藨子、唐古特忍冬等。红桦-岷江冷杉林样地内的乔木层郁闭度为0.64,优势种为岷江冷杉和红桦,伴生有陕甘花楸、桦叶荚蒾(Viburnumbetulifolium)、云杉等,灌木层盖度为0.45,主要有华西箭竹、栒子木、唐古特忍冬等。
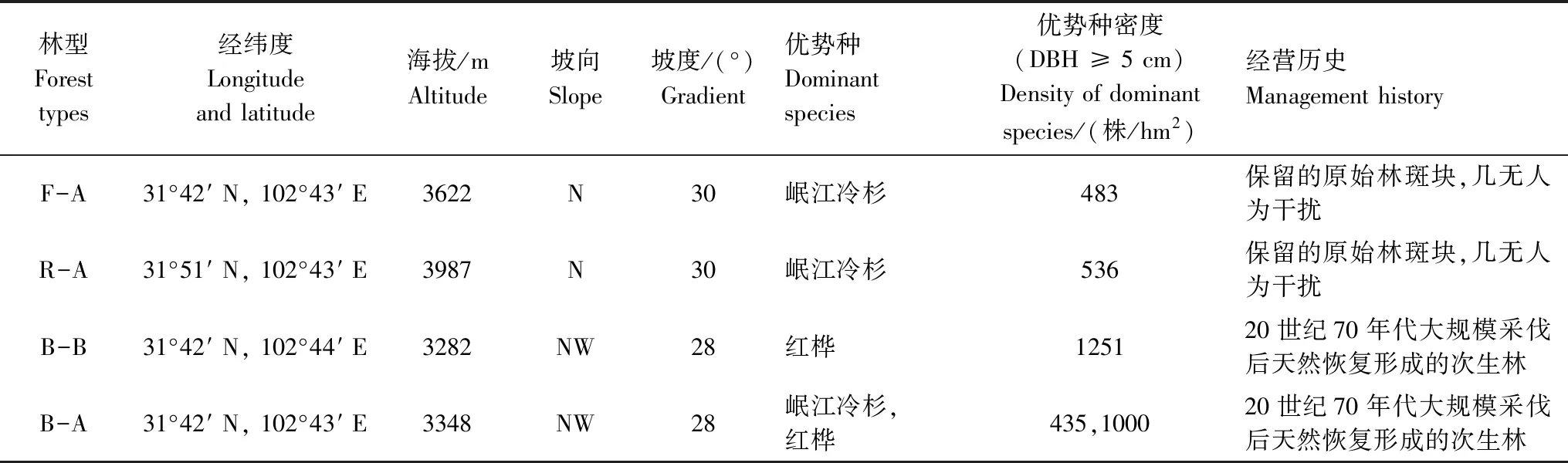
表1 四个样地的基本情况Table 1 Basic information of the four research plots
将每块样地分为25个20 m×20 m的样方,测定其中岷江冷杉的高度(H)和胸径(DBH)数据,并记录植株的生长(死活)状态。不同生长阶段岷江冷杉的划分标准为:幼苗(H<30 cm);幼树(30 cm≤H<2 m);母树(DBH≥25 cm,年龄超过110年)[26]。4种林型的1 hm2样地中,岷江冷杉幼苗和幼树数量均较多,无法全部查数年龄。因此,为满足建立年龄与树高的回归模型样本量,我们对样地中的部分幼龄植株(幼苗和幼树,H<2 m)年龄采用查数轮生枝法确定。杜鹃-岷江冷杉林、箭竹-岷江冷杉林、红桦-岷江冷杉林和红桦林中的幼龄植株样本量分别为407、787、519株和1259株。
1.3 更新质量衡量
根据岷江冷杉的高度、胸径和生长(死活)状况等数据,统计岷江冷杉各生长阶段内个体数量,将样地中幼苗和幼树密度之和与母树密度的比值(粗密度Crude density)作为衡量岷江冷杉更新质量的指标。个别20 m × 20 m样方内岷江冷杉母树或幼龄植株的数量为零或因聚集而数量极多,为了避免岷江冷杉空间分布的聚集性对林型之间更新质量对比分析产生影响,4种林型的粗密度数值以整个样地内的母树、幼树和幼苗密度来计算。用单因素方差分析(ANOVA)对不同林型中幼苗、幼树和母树密度的差异显著性进行分析,选择LSD模型进行多重比较。
1.4 幼龄植株的静态生命表编制
幼龄植株(幼苗和幼树,H<2 m)的静态生命表能在一定程度上反映岷江冷杉幼龄植株的定居过程。由于岷江冷杉更新存在年际波动性,生命表有可能会出现死亡率为负值的情况,因此参照江洪的匀滑技术对各龄级的实际个体数(Ax)进行匀滑修正得到ax(实际存活个体数经匀滑处理后所得存活数)[31]。以5年为一个龄级,将已知年龄的幼龄植株划分为8个龄级,编制出岷江冷杉幼龄植株的静态生命表。分别以标准化存活个体数的对数lnlx和死亡率qx为纵坐标,龄级为横坐标,绘制岷江冷杉幼龄植株的存活曲线和死亡率曲线。
1.5 幼龄植株的种群数量动态分析
为更准确判定岷江冷杉幼龄植株受到外界干扰的影响或制约较大的年龄阶段,用陈晓德[32]的种群结构动态量化方法对(幼龄)种群进行定量描述。相邻龄级间的个体数量动态量化指数(Vn)、整个种群年龄结构的数量变化动态指数(Vpi)与外部干扰纳入影响因子后的年龄结构动态指数(Vpi′)取正值、负值和零的意义分别反映相邻龄级或种群个体数的增长、衰退和稳定的动态关系[32]。计算公式如下:
式中,Vn为种群从n到n+1级的个体数量变化动态;Vpi为种群年龄结构的数量变化动态指数;Sn与Sn+1分别为第n与第n+1年龄级种群个体数;max(Sn,Sn+1)表示取括号中数列最大值;k为种群大小级数量。Vpi仅适用于不考虑外部干扰时的种群结构动态比较,当考虑外部干扰时,则Vpi还与年龄级数量(k)及各年龄级个体数(Sn)两因素相关,因此对Vpi修正为:
式中,P极大为干扰下种群承担的最大风险概率;min(S1,S1,S3,…,Sk)表示取括号中数列最小值。
统计分析在SPSS 26.0中进行,图表绘制采用Origin 2018进行,检验显著性水平α=0.05。
2 结果与分析
2.1 4种林型中岷江冷杉的种群结构
从整体看,四个林型中岷江冷杉种群均呈现为增长型(图1)。即使在不同林型间,岷江冷杉种群的径级结构依然相似。其中,红桦-岷江冷杉林与箭竹-岷江冷杉林树高≤1.3 m个体占比均在50%—60%间,树高>1.3 m个体占比随着径级的增大而先减后增;红桦林的种群结构波动较大,体现在0—20 cm径级的个体占比不随径级的增大而减小,而是呈现先增后减的波动趋势。杜鹃-岷江冷杉林中树高≤1.3 m的个体占比(82.59%)与其他径级差距极大,其余径级占比最高仅为4.29%。
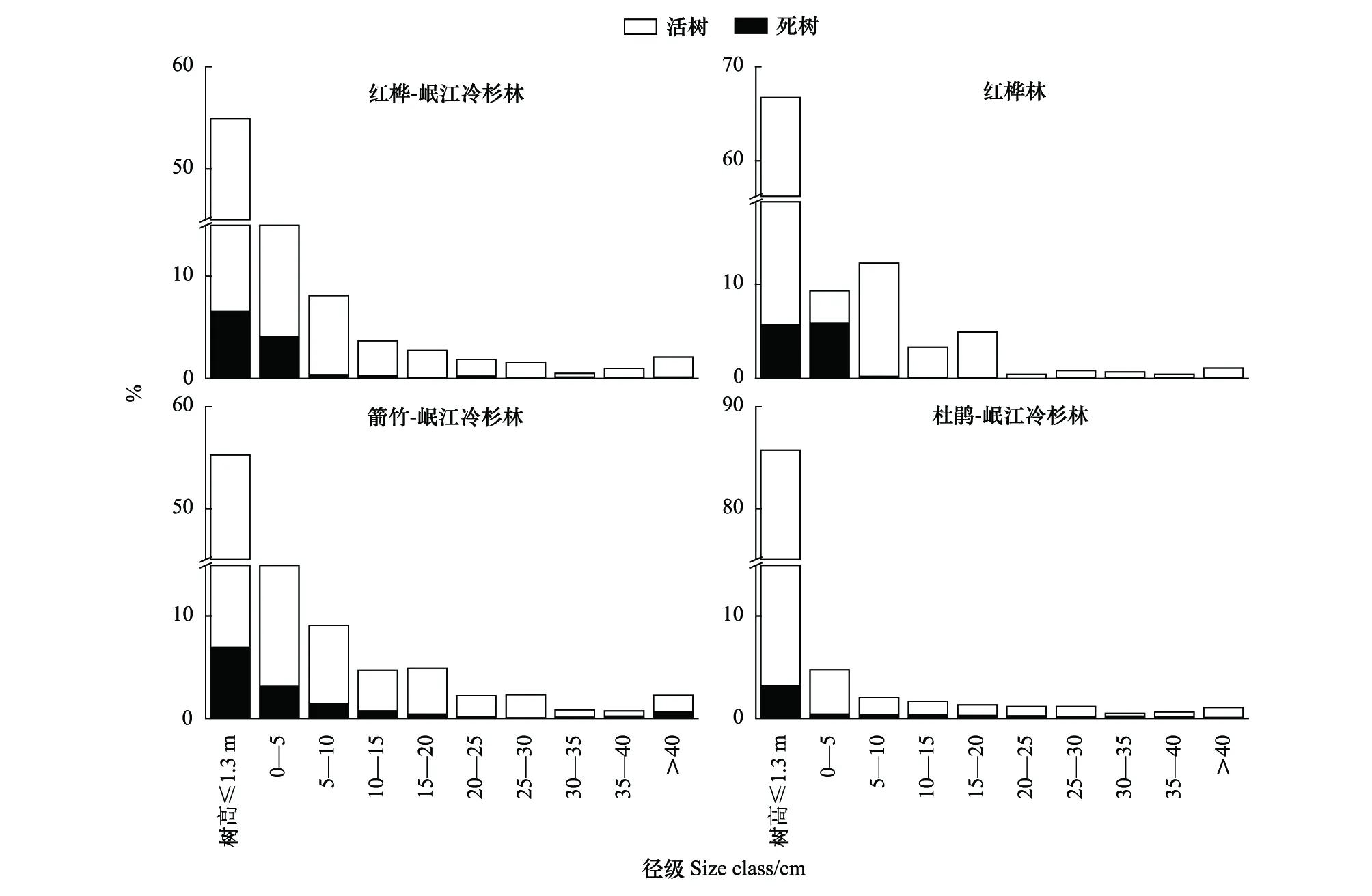
图1 4种林型中岷江冷杉种群的径级结构Fig.1 Size-class distributions of the Minjiang fir in the four forest types
随着岷江冷杉径级增大,死树占比越来越小。不同林型间树高≤1.3 m的幼龄存活个体占其种群的比例依次为:杜鹃-岷江冷杉林(82.59%)>红桦林(61.03%)>红桦-岷江冷杉林(48.38%)>箭竹-岷江冷杉林(48.27%)(图2)。
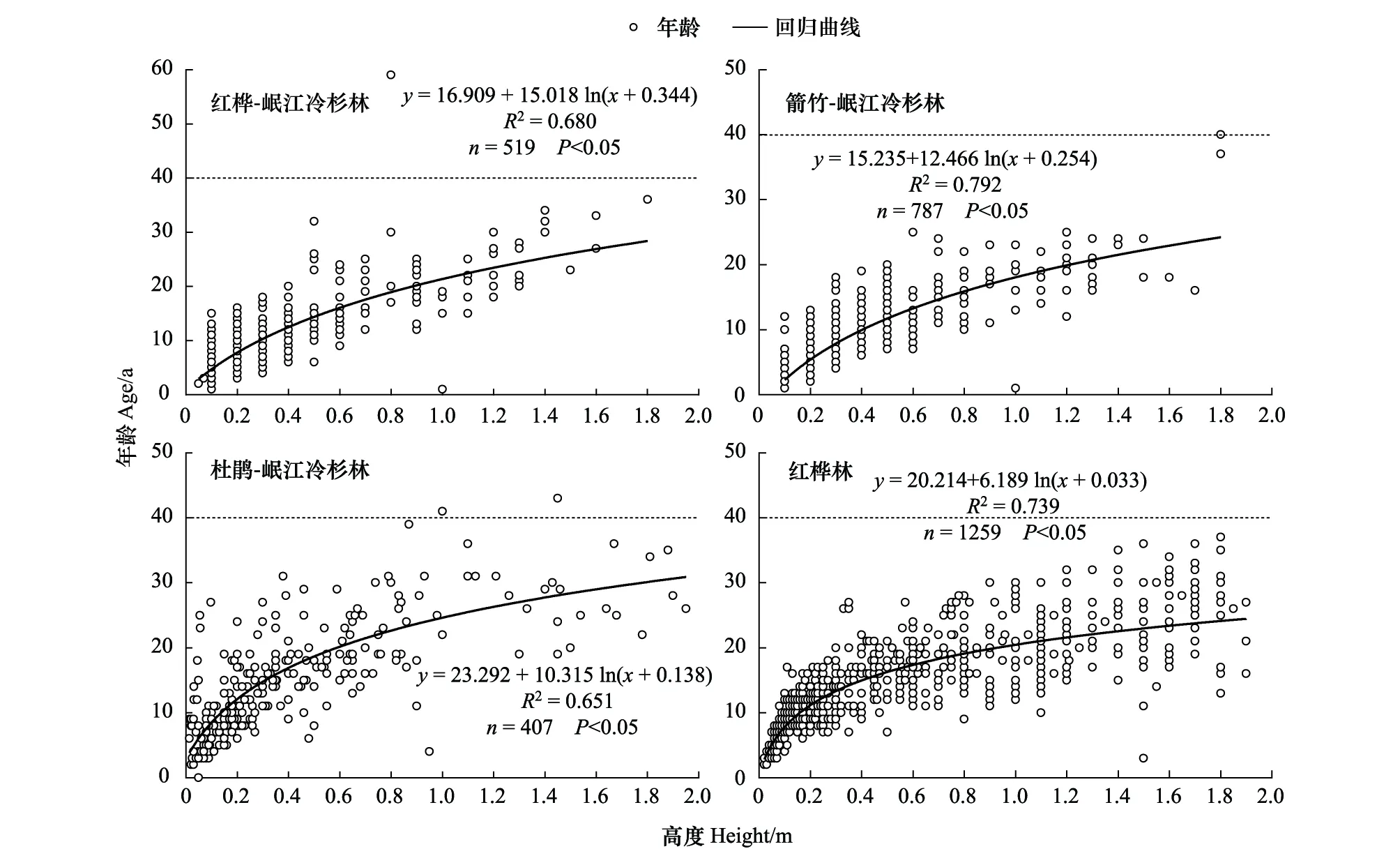
图2 不同林型中岷江冷杉幼龄植株高度与年龄的回归方程Fig.2 Regression equations between tree age and height of young Minjiang fir in the four forest types
基于岷江冷杉幼龄植株的高度和年龄数据,建立了其高度-年龄的回归模型(图2)。4种林型中幼龄植株的高度和年龄拟合模型均为对数模型,拟合参数均达到显著水平(P<0.05),表明拟合效果较好。在2972株岷江冷杉幼龄植株中,仅有3株高度小于 2 m的幼龄植株年龄超过40 年,其余99.9%的幼龄植株年龄均在40年以内(见图2虚线部分)。
2.2 4种林型中岷江冷杉的更新质量
在4种林型中,不同生长阶段岷江冷的杉密度均表现为幼苗>幼树>母树(表2)。杜鹃-岷江冷杉林中岷江冷杉幼龄植株的密度均显著大于其他三种林型,母树密度与箭竹-岷江冷杉林间无显著差异(P<0.05)。红桦林中岷江冷杉幼龄植株密度仅次于杜鹃-岷江冷杉林,但其母树密度却为最小,与红桦-岷江冷杉林中的母树密度无显著差异(P<0.05)。4种林型间箭竹-岷江冷杉林的粗密度最小(4.8),红桦-岷江冷杉林的数值(10.3)为箭竹-岷江冷杉林的2.1倍,红桦林的粗密度(24.8)为箭竹-岷江冷杉林的5.2倍,杜鹃-岷江冷杉林的粗密度值最大(26.1),为箭竹-岷江冷杉林的5.5倍。
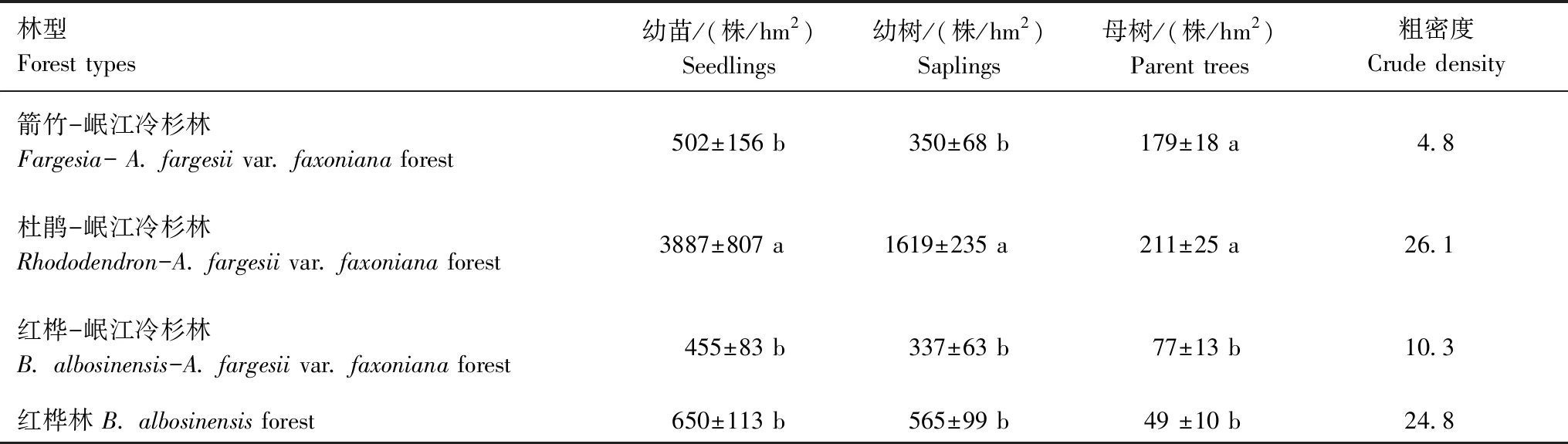
表2 4种林型中的粗密度更新指标Table 2 Crude density in the four types of forest
2.3 4种林型中岷江冷杉幼龄植株的生存过程
总体上,不同林型间岷江冷杉幼龄植株的年龄结构相似,每个林型的实际存活总数(Ax)中超过56%的幼龄个体均分布在第Ⅰ—Ⅲ龄级(1—15年)间(表3)。4种林型中,岷江冷杉幼龄植株的标准化存活数(lx)随龄级增大而趋于减少,死亡率(qx)与消失率(Kx)随着龄级增大的波动较大。

表3 4种林型岷江冷杉幼龄植株的静态生命表Table 3 Stationary life tables of the young Minjiang fir in the four forest types
各林型中岷江冷杉幼龄植株的存活数量随龄级变化的总趋势表现为:随龄级增加其存活数量趋于下降,存活数量排序依次为:杜鹃-岷江冷杉林>红桦林>红桦-岷江冷杉林>箭竹-岷江冷杉林(图3)。各林型中幼龄植株的死亡率随龄级增大的变化剧烈(图3),反映了岷江冷杉种子大小年和林冠更替周期波动的更新特点。
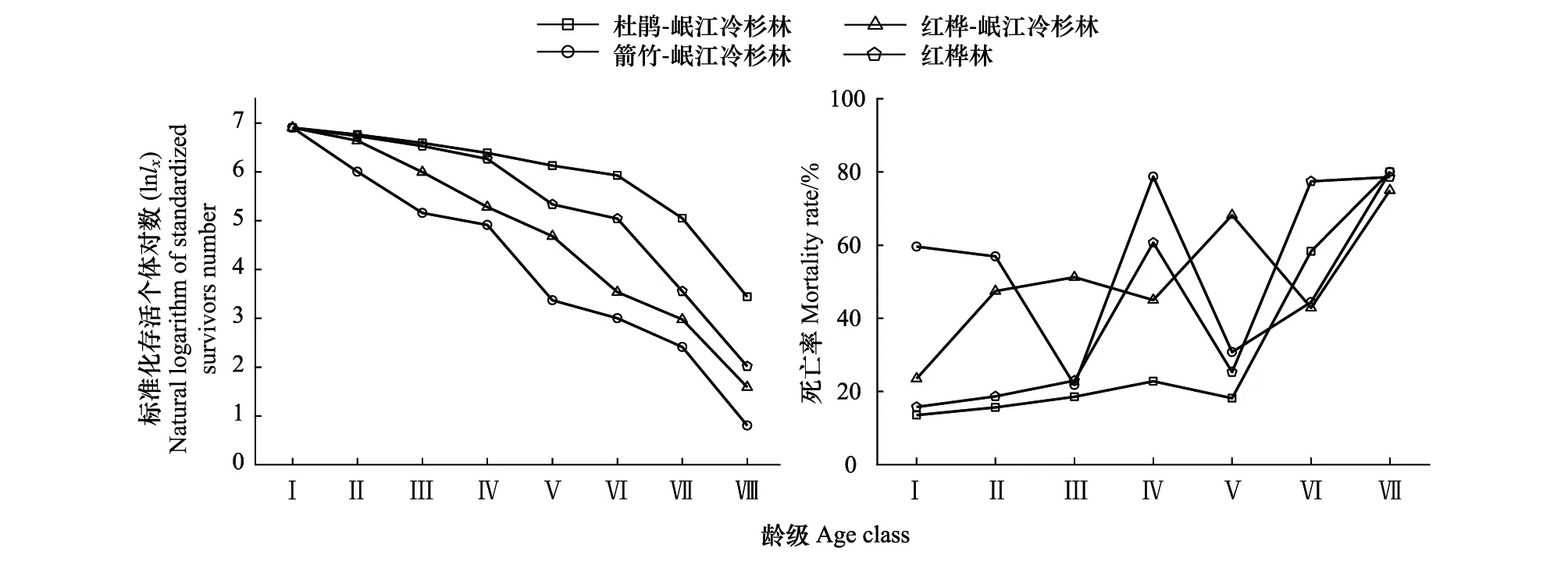
图3 4种林型中岷江冷杉幼龄植株的存活曲线和死亡率曲线Fig.3 Survival curve and mortality curve of the young Minjiang fir in the four forest typesⅠ—Ⅷ:第1—8龄级
2.4 4种林型中岷江冷杉幼龄植株的数量动态
随着龄级的增加,杜鹃-岷江冷杉林和红桦林中岷江冷杉幼龄植株的龄级结构动态量化指数Vn在第Ⅰ龄级均为负值(-37.23%和-48.71%),而箭竹-岷江冷杉林和红桦-岷江冷杉林中的Vn指数却分别在第Ⅰ龄级和第Ⅱ龄级取得最大的正值(59.60%和36.27%)。除第Ⅰ龄级,在杜鹃-岷江冷杉林和红桦林中,Vn指数随着龄级增加的波动较大,且这两种林型中幼龄植株在第Ⅲ、第Ⅴ龄级和第Ⅶ龄级的Vn指数小于前一龄级。箭竹-岷江冷杉林中幼龄植株第Ⅲ龄级和第Ⅴ龄级的Vn指数小于前一龄级,且其第Ⅵ龄级和第Ⅶ龄级为零。红桦-岷江冷杉林幼龄植株的Vn指数在第Ⅱ龄级出现峰值后,随着龄级的增加,Vn指数逐渐减小,但均维持在正值(图4)。
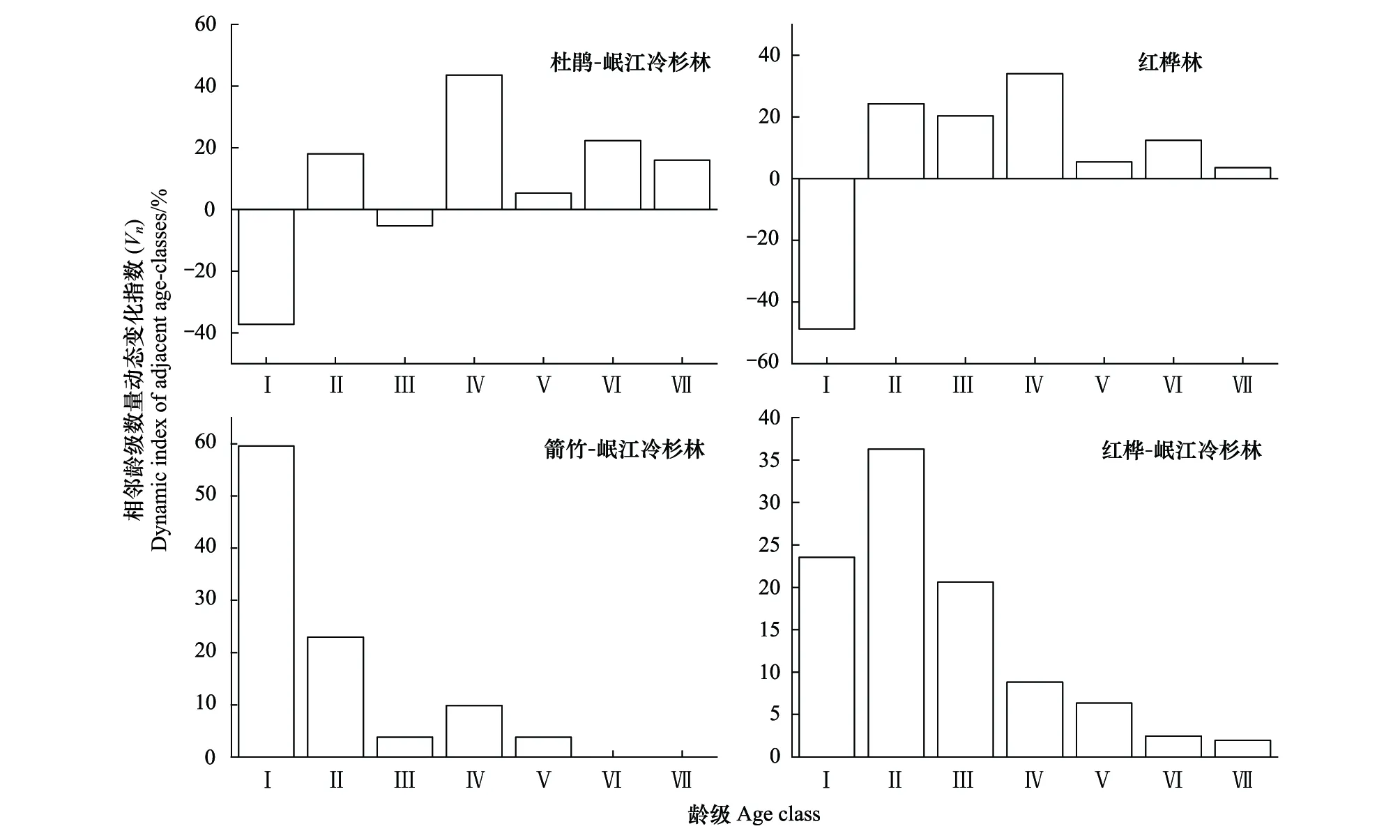
图4 4种林型中岷江冷杉幼龄植株相邻龄级数量变化的动态指数(Vn)Fig.4 Dynamic index (Vn) of quantitative changes of adjacent age-classes of the young Minjiang fir in the four forest types
4种林型中表征岷江冷杉幼龄植株相邻龄级种群结构动态的指数Vn变化趋势各异(图4),结合幼龄植株死亡率在不同林型和不同龄级间的变化趋势(图3),可以综合分析出幼龄植株存活率急剧下降的龄级阶段。原始林中,杜鹃-岷江冷杉林中岷江冷杉幼龄植株的死亡率在第Ⅰ—Ⅴ龄级之间均在22%以下,较为平稳,但从第Ⅴ龄级到第Ⅵ期间,死亡率随龄级开始急剧上升,直至第Ⅶ龄级的80%(图3);此外,种群龄级结构动态量化指数(Vn指数)值从在第Ⅳ龄级的43.62%降低为第Ⅴ龄级的5.32%(图4)。因此,我们推断第Ⅴ龄级(第21—25年,树高0.66—1.04 m)为该林型中岷江冷杉幼龄植株存活的瓶颈期。箭竹-岷江冷杉林中幼龄植株的死亡率在第Ⅲ龄级前逐渐下降,但从第Ⅲ龄级到第Ⅳ龄级期间,死亡率开始从第Ⅲ龄级急剧上升至第Ⅳ龄级的78%,该死亡率仅次于第Ⅶ龄级80%的死亡率峰值(图3);此外,种群龄级结构动态量化指数(Vn指数)值从第Ⅱ龄级的22.99%降为第Ⅲ龄级的3.79%(图4)。因此,我们推断岷江冷杉幼龄植株存活的瓶颈期为第Ⅲ龄级(第11—15年,树高0.46—0.73 m)。
次生林中,红桦-岷江冷杉林中岷江冷杉幼龄植株的死亡率在第Ⅳ龄级前的增长趋势较平缓,从第Ⅳ龄级开始其死亡率急剧上升至68%,在第Ⅴ—Ⅶ龄级的死亡率呈现出较大的波动(42%—75%,图3);此外,种群龄级结构动态量化指数(Vn指数)值从第Ⅲ龄级的20.59%降为第Ⅳ龄级的8.82%(图4)。因此,我们推断第Ⅳ龄级(第16—20年,树高0.60—0.89 m)是红桦-岷江冷杉林中幼龄植株存活的瓶颈期。红桦林中岷江冷杉幼龄植株的死亡率在第Ⅲ龄级前较低且变化平缓,从第Ⅲ龄级开始其死亡率急剧上升至60%,且第Ⅳ—Ⅶ龄级的死亡率呈现出较大的波动趋势(25%—78%,图3);此外,种群龄级结构动态量化指数(Vn指数)值从第Ⅱ龄级的24%降低为第Ⅲ龄级的20%(图4)。因此,我们推断第Ⅲ龄级(第11—15年,树高0.19—0.40 m)为红桦林中的岷江冷杉幼龄植株存活的瓶颈期。
依据岷江冷杉幼龄植株年龄结构动态的Vpi和Vpi′指数模型,在无外部干扰时,岷江冷杉幼龄植株的种群年龄结构数量变化动态指数Vpi在箭竹-岷江冷杉林中为最高(40.43%),杜鹃-岷江冷杉林中为最低(9.73%),中间依次为红桦-岷江冷杉林(24.51%)和红桦林(11.37%) (图5)。当有外部干扰时,4种林型中岷江冷杉幼龄植株的种群年龄结构指数Vpi′随林型的变化呈现出与Vpi相同的变化规律,但Vpi′值分别锐减至2.53%、0.41%、0.77%和0.47%;Vpi和Vpi′均为正值(图5),种群均为增长型。其中,箭竹-岷江冷杉林林中岷江冷杉幼龄植株的种群结构始终有最大的增长潜力,而杜鹃-岷江冷杉林林中岷江冷杉幼龄植株的种群结构的增长潜力始终最低。
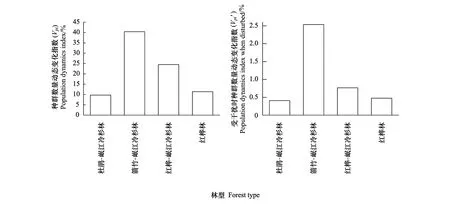
图5 4种林型中岷江冷杉幼龄植株年龄结构的数量变化动态指数(Vpi和Vpi′)Fig.5 Dynamic index (Vpi and Vpi′) of quantitative changes in the age structure of the young Minjiang fir in the four forest types
3 讨论
3.1 杜鹃-岷江冷杉林中岷江冷杉的更新质量和更新潜力
本研究中,杜鹃-岷江冷杉林中岷江冷杉的幼苗和幼树密度均显著高于箭竹-岷江冷杉林、红桦-岷江冷杉林和红桦林,且其更新质量和幼龄植株的存活状态亦优于其余三种林型。有研究亦表明,岷江冷杉更新密度的变化与海拔密切相关,如张远东等[28]证实在靠近林线的海拔3800—4000 m范围内,幼龄植株密度达到峰值。杜鹃-岷江冷杉林中的高密度更新可能与林下光照充足和无箭竹等灌木的制约有关[28]。此外,杜鹃作为高海拔林线区域及其下方的优势树木,与岷江冷杉整体上形成了共存关系。缪宁等[24]在川西鹧鸪山的林线附近发现,凝毛杜鹃(Rhododendronphaeochrysumvar.agglutinatum)大树在小尺度范围内对岷江冷杉幼树具有庇护作用,幼龄岷江冷杉个体的存活和生长可能一定程度上依赖于杜鹃的庇护和遮阴。Germino等[33]在美国Medicine Bow国家森林公园的研究也发现,高山林线环境下杜鹃的郁闭作用促进了银云杉(P.engelmannii)和毛果冷杉(A.lasiocarpa)幼苗的定居,这在高山林线森林群落的构建过程中起着重要的作用。
杜鹃-岷江冷杉林中岷江冷杉幼龄植株相邻龄级数量变化的动态指数(Vn)为负值的龄级数量较其余林型更多,表明该林型中的幼龄植株生存受到干扰的龄级更多,且其种群增长潜力(Vpi和Vpi′)在4种林型中表现最低,这可能是由于该林型处于林线环境,幼龄植株受到极端生境的胁迫和压力较大[34—35]。第Ⅴ龄级(第21—25年,树高0.66—1.04 m)为该林型中岷江冷杉幼龄植株存活的瓶颈期,该阶段幼龄个体死亡率从18%开始急剧上升,直至第Ⅶ龄级的80%,表明从第Ⅴ龄级开始绝大部分幼龄植株难以存活至下一阶段。在幼龄阶段第Ⅵ—Ⅶ龄级,岷江冷杉死亡率急剧上升的阶段现象,可能与数量众多的杜鹃中树对岷江冷杉小树的抑制或者排斥作用有关[24]。
3.2 箭竹-岷江冷杉林中岷江冷杉的更新质量和更新潜力
同为未采伐的原始林,箭竹-岷江冷杉林林中岷江冷杉母树密度与杜鹃-岷江冷杉林中母树密度的差异不显著,说明其母树提供种源的能力大致相当。然而,箭竹-岷江冷杉林中岷江冷杉的粗密度和幼龄植株在各龄级中的存活量却比其他三种林型更低,其更新质量和幼龄植株的存活状态均表现为最差。此外,该林型幼龄植株存活的瓶颈期(第11—15年,树高0.46—0.73 m)也较杜鹃-岷江冷杉林更早出现,这可能是两原始林林下灌木层片优势种的不同导致。研究表明,杜鹃-岷江冷杉林下灌木层优势种杜鹃与岷江冷杉间总体上是促进和庇护的有利关系[24],而箭竹-岷江冷杉林下的箭竹对岷江冷杉幼龄植株通常表现出抑制和阻碍作用[14,16,27]。
箭竹-岷江冷杉林中岷江冷杉幼龄植株在第Ⅰ和第Ⅱ龄级(10年内幼苗)的死亡率明显高于其余三种林型,可能是因为林下箭竹严重制约了第1—10年,即树高0.10—0.45 m阶段幼龄植株的生存和生长。研究表明,箭竹通过地下茎的无性繁殖蔓延扩张,并通过其密集交错的秆枝和根系与岷江冷杉幼苗争夺光照、水分、养分等资源[7,16]。此外,箭竹常形成不易分解的厚凋落物层,层下生境阴湿,不利于岷江冷杉幼苗的存活[17]。然而,箭竹-岷江冷杉林中岷江冷杉幼龄植株的种群增长潜力(Vpi′)大于其余三种林型。有研究表明箭竹对幼龄岷江冷杉的生长并非始终是负面作用,比如,在四川卧龙的原始岷江冷杉暗针叶林内,低密度(10—15株/m2)竹丛的条件下,当其进入幼树阶段后,箭竹对其的抑制作用减弱,能为其提供相对良好的微生境[6]。因此,该林型中岷江冷杉的增长潜力大可能是由于其林下箭竹的平均密度相对较低(0.1株/m2)。
3.3 不同林型间岷江冷杉幼龄植株生存的制约瓶颈阶段差异
本研究中,阔叶林阶段的红桦林中岷江冷杉幼龄植株存活的瓶颈期(第11—15年,树高0.19—0.40 m)较针阔混交林阶段的红桦-岷江冷杉林(第16—20年,树高0.60—0.89 m)更早出现。对岷江冷杉皆伐后次生群落结构和物种多样性演替动态的研究表明,随着次生演替的进行,从桦木阔叶林阶段至桦木-岷江冷杉混交林阶段,林下草本层的盖度由70%逐渐减小至30%左右,而灌木层盖度由70%升至80%的更高水平[36]。因此,这种阔叶林中幼龄植株存活瓶颈期的更早出现可能与该区域红桦阔叶林的林下草本盖度比针阔混交林的草本盖度更大,且草本对岷江冷杉幼苗生长的抑制作用有关[5,22]。此外,两种天然次生林中的岷江冷杉幼龄植株存活的瓶颈期较杜鹃-岷江冷杉原始林(第21—25年,树高0.66—1.04 m)均更早出现。这可能是由于两种次生林中的灌木茂密、草本盖度大[36],导致第Ⅲ龄级以上(10年以上)的幼苗死亡率显著上升,而杜鹃-岷江冷杉林中杜鹃灌丛相对稀疏、林下透光率较高[28],对岷江冷杉幼龄个体早期(年龄<21年,树高<0.66 m)的存活和生长有利。岷江冷杉幼苗的定居和存活相对母树和其他龄级个体独立[26,37],初期定居仅对幼苗周边的微生境条件,如苔藓、凋落物、光照等较为依赖[5];随着岷江冷杉年龄和树高的增加,更高大的草本、灌木(如,箭竹[6,15,17]或杜鹃[24,38])和优势乔木(如,岷江冷杉[37]与红桦[39])则会依次对其存活、生长和空间分布产生直接影响。
4 结论
本研究对比分析了川西亚高山地区4种典型天然林林型中岷江冷杉的更新结构、更新质量和幼龄植株龄级结构的数量动态特征。不同林型中岷江冷杉天然更新幼龄植株的数量丰富,但更新幼龄植株存活至后续生长阶段的比率较低。4种林型间岷江冷杉的更新质量和幼龄植株存活状态表现依次为:杜鹃-岷江冷杉林>红桦林>红桦-岷江冷杉林>箭竹-岷江冷杉林。原始林中,箭竹-岷江冷杉林中幼龄植株存活的瓶颈期(第11—15年,树高0.46—0.73 m)比杜鹃-岷江冷杉林(第21—25年,树高0.66—1.04 m)更早出现。天然次生林中,幼龄植株存活的瓶颈期比杜鹃-岷江冷杉原始林中的幼龄植株更早出现;处于阔叶林阶段的红桦林中幼龄植株存活瓶颈期(第11—15年树高,0.19—0.40 m)比处于针阔混交林阶段的红桦-岷江冷杉林(第16—20年,树高0.60—0.89 m)更早出现。4种典型天然林林型间的更新差异不仅来自于母树密度和灌木层的竞争差异,还与海拔密切相关,并且海拔变化也同时引起灌木种类及其盖度的变化,这些因素交织在一起,对岷江冷杉的更新格局共同产生影响。在对不同林型中岷江冷杉进行人工促进天然更新时,可在次生林中岷江冷杉幼龄个体到达瓶颈期之前及时清除其周边杂草和灌木;在原始林中则采取局部透光伐的抚育措施,以提升25年以上幼龄植株的存活率。
本研究对川西亚高山典型林型中岷江冷杉幼龄植株存活瓶颈期产生的原因进行了初步分析,针对幼龄植株受制约阶段的关键影响因子及其生态学机制还有待于深入研究。在全球气候变化背景下,高海拔针叶林中更新状况的变化趋势如何,也有待进一步揭示,故对不同林型更新动态的持续性监测和评估非常必要。
猜你喜欢
湖南生态科学学报(2022年4期)2022-11-11
西北林学院学报(2021年6期)2021-12-07
世界竹藤通讯(2020年4期)2020-12-23
河南农业大学学报(2020年2期)2020-05-22
山西林业科技(2019年1期)2019-05-27
东北农业大学学报(2018年11期)2018-12-21
南方农业学报(2018年4期)2018-09-10
防护林科技(2017年4期)2017-04-27
安徽农学通报(2016年14期)2016-09-07
西部林业科学(2015年3期)2015-01-17
