
巴丹吉林沙漠东缘天然梭梭种群结构与动态特征
2023-06-28 03:37蒙仲举党晓宏赵飞燕王德慧柴享贤
干旱区研究 2023年6期
毛 毛, 蒙仲举, 党晓宏, 赵飞燕, 王德慧, 柴享贤
(1.内蒙古农业大学沙漠治理学院,内蒙古 呼和浩特 010018;2.内蒙古兴安职业技术学院农牧系,内蒙古兴安盟137400;3.孪井滩生态移民示范区工业和科技局,内蒙古 阿拉善盟 750312)
种群是指同一时间生活在一定自然区域内,同种生物的所有个体,是进化的基本单位[1]。种群结构与动态常用来反映同一时间生活在一定自然区域内种群的变化规律,是种群个体与受外界共同影响的结果,也是种群生态学的重要研究课题[2]。种群结构与动态能够使种群生存现状数据化,评估种群发展动态,可以理解种群对周围资源环境的适应性、其生长特性以及生存潜力,有助于理解同一种群个体间,以及种间相互关系及整个群落的演替规律[3-5]。近年来,由于气候变化与人类过度干扰,干旱发生的频率也越来越高[6],在特别干旱的区域内,已经在生态系统结构上造成了植物种群的退化甚至死亡,严重影响了生态系统的服务价值功能,因此在生态学上研究植物种群对干旱的适应以及响应已经成为热点。
而本身耐干旱植物例如梭梭(Haloxylon ammodendron)、杨柴(Hedysarum laeve)、花棒(Hedysarum scoparium)等,因其本身的耐旱性能,更成为重点关注研究的植物[7]。这其中,作为沙漠先锋植物的梭梭,其分布广泛,不仅耐干旱,且抗逆性强,具有极高的生态经济价值[8]。梭梭是藜科梭梭属[9]灌木,耐寒抗旱,适应风蚀沙埋,在沙漠治理中发挥着不可替代的作用[10]。位于巴丹吉林沙漠东缘的塔木素,现有天然梭梭21.5×104hm2[11],位于沙漠与绿洲的交错区域[12],其植物种群的退化与扩张,会影响到绿洲的扩张或者衰退,因此,明晰其种群结构和动态是探究该研究区天然梭梭林能否实现自我更新的关键,对探究该区域群落的演替具有重要意义。
目前,对于梭梭的研究集中于种群特征[13],种群结构和空间分布格局[14-15],以及种群数量动态[16-18],民勤红石荒漠区的梭梭分为3 种类型,但是研究其种群分布格局发现,无论是哪一种梭梭,均对荒漠干旱有较强的适应性[13];同时对甘家湖梭梭的研究结论也相似,说明梭梭处于不同的生长阶段会根据环境做出不同的生长响应[14-15]。综上所述,目前对梭梭的研究大多集中于梭梭的空间分布[18-22],对该研究区天然梭梭林种群结构与动态特征的研究鲜见报道。
对梭梭种群数量的动态研究可以突破目前对于梭梭整体科学管理的瓶颈,对梭梭种群生长更新的策略,后续的形成发展以及对环境的稳定性都有较为重要的意义,同时,也可以在一定程度上对巴丹吉林沙漠东缘的绿洲与沙漠的动态变化特征加以掌握,以此对环境恶化区域提前进行干预。因此,本文以分布于巴丹吉林沙漠东缘的天然梭梭林为研究对象,对其种群动态发展趋势进行探究,进而为塔木素地区梭梭种群保护与当地生态可持续恢复提供理论依据和数据支撑。
1 研究区概况与方法
1.1 研究区概况
研究区位于内蒙古阿拉善盟阿拉善右旗塔木素苏木格日勒图嘎查的朝恒扎干(40°33.06′~40°35.53′N,103°25.16′103°~27.55′E),海 拔 为1255.8~1264.3 m[11]。研究区属暖温带荒漠干旱区,为典型的干燥大陆性气候。年均降水量53.4 mm,年蒸发量>3500 mm,年均温8.4 ℃,地下水位低于500 m,局部低于1000 m[23],自然植被稀少,以降水为主要供给类型。主要植物有梭梭、白刺(Nitraria tangutorum)、红砂(Reaumuria songarica)、霸王(Sarcozygium xanthoxylon)等。
1.2 研究方法
1.2.1 样地设置 2020 年9 月,以巴丹吉林沙漠东缘的天然梭梭林为研究区域(图1),研究区总面积923 m2,采用大样地调查法,选取梭梭分布密集的区域作为实验样地,采用手持全球定位系统记录样地4个端点处的坐标,以左下角作为起始点,以横向和纵向每隔200 m 分别设置样线,形成48 个200 m×200 m的相邻网格。对样线上出现的每一株梭梭进行测量,记录其基径和相对坐标。

图1 试验样地示意图Fig.1 Geographical position of the test area
1.2.2 梭梭种群径级结构划分 测量中发现梭梭地上茎株增长不规则,其株龄很难用年轮来表示[24]。因此,参考李艳丽等[25]的方法,采用径级代替龄级。参考蔡新斌等[16]和吕朝燕等[26]的划分标准,将其划分为17个龄级,每个龄级之间相差2。同时,将17个龄级分为4个阶段,即幼龄阶段(Ⅰ~Ⅲ)、中龄阶段(Ⅳ~Ⅵ)、成龄阶段(Ⅶ~Ⅸ)、老龄阶段(Ⅹ~ⅩⅦ)。
1.2.3 静态生命表编制 由于所研究对象为天然梭梭林,本文采用匀滑(Smooth Out)技术[27-28]处理,对梭梭林各龄级的个体数进行统计,按照静态生命表编制方法进行编制。
1.2.4 梭梭种群存活曲线 本文将存活曲线分为以下3种类型[29]:Deevey-Ⅰ型、Deevey-Ⅱ型、Deevey-Ⅲ型,来描述生存规律。参考Hett 等[30]的数学模型,分别检验种群存活曲线类型。
1.2.5 种群结构与动态量化方法 梭梭种群动态结构分析参考陈晓德[31]的研究方法:
式中:Vn为动态变化指数;Vpi和分别为忽略和考虑外部干扰情况下种群的数量动态变化指数;Sn为种群个体数;Sn+1为n+1 龄级种群个体数;k为龄级总数量。
1.2.6 时间序列预测模型 本文选用一次平均推移法[32]进行模拟和预测。公式如下:
式中:n为龄级;t为预测时间;Xk为k龄级的种群存活数;Mn为种群存活数预测值。
1.3 数据分析
本文采用Excel 2020、Origin 2019 和ArcGIS 进行分析和作图。
2 结果与分析
2.1 梭梭种群年龄结构
本研究共调查梭梭样本1151 株,最大基径37 cm。由图2 可知,研究区该种群龄级结构呈“J”型,属增长型。Ⅰ~Ⅲ龄级个体数之和占比56.21%,表明梭梭幼龄个体数量多,具有较强的增长潜力。Ⅳ和Ⅴ龄级分别占种群个体总数的10.08%和9.73%,较前一龄级降低趋势明显,说明幼龄到中龄过渡困难;Ⅵ龄级以后梭梭个体数量逐步降低,且Ⅴ~Ⅵ龄级降低趋势明显,说明仅有少量个体能够过渡到成龄阶段,成长为成龄个体后生命力增强。老龄仅占个体总数的3.04%。
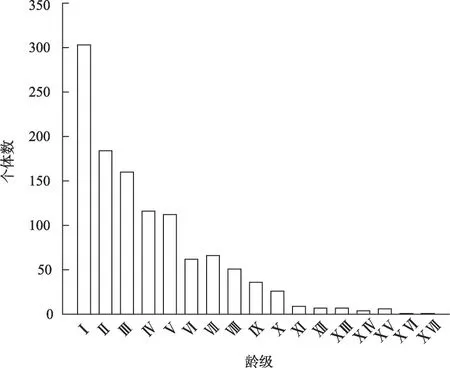
图2 梭梭种群径级结构Fig.2 Age structure of Haloxylon ammodendron population
2.2 梭梭种群静态生命表
由表1 可知,龄级越大梭梭数量越少。该种群在第Ⅳ龄级时ex出现峰值,qx和Kx进入低谷,说明幼龄整体发展较好。从第Ⅳ龄级开始,qx和Kx逐渐增大,ex逐渐减小,表明梭梭开始衰老,第Ⅷ龄级开始出现不稳定变化。从Ⅺ龄级开始ex持续下降,到ⅩⅥ和ⅩⅦ龄级达最小值,反之qx和Kx最大,说明少数存活下来的梭梭个体正处于退化阶段。
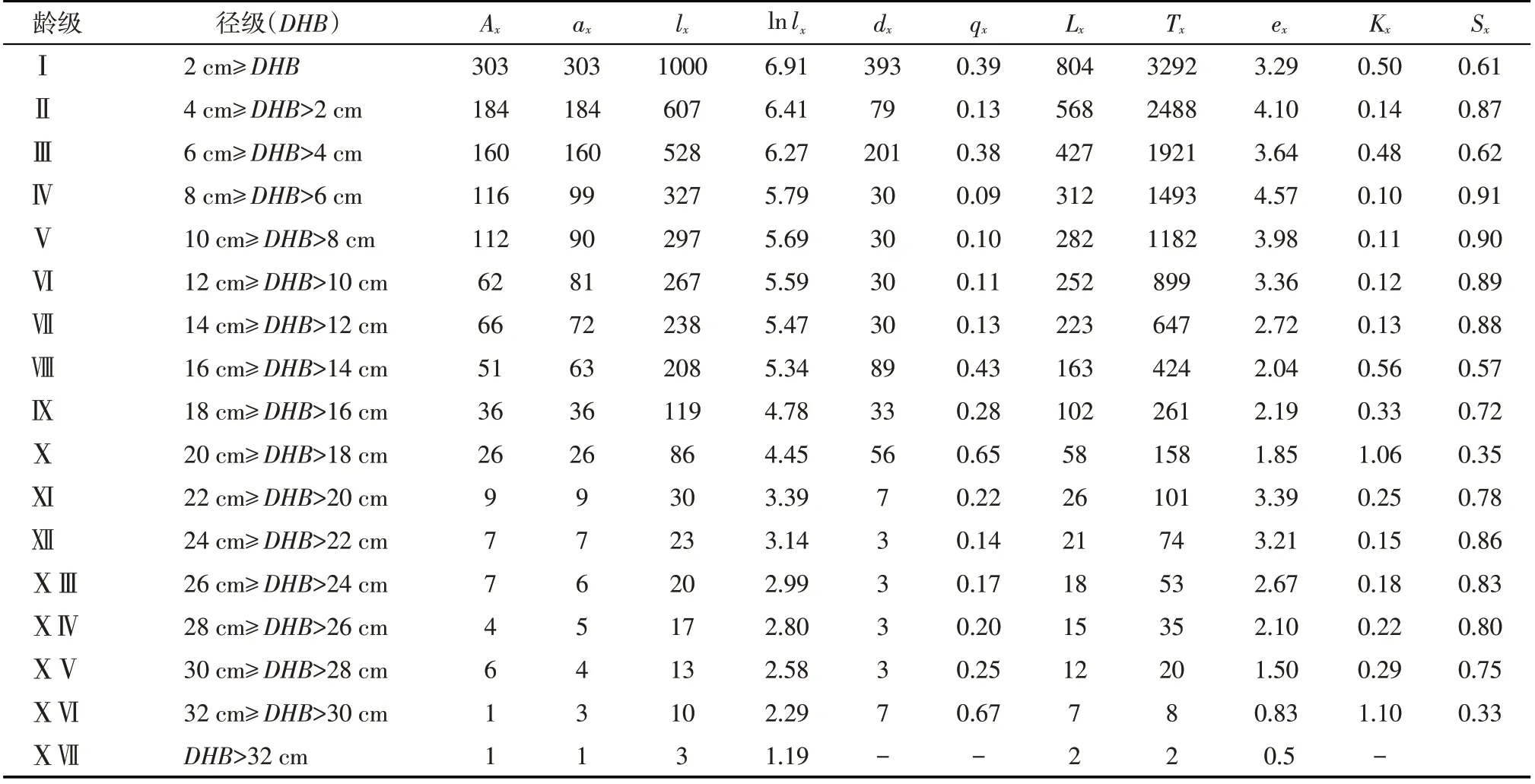
表1 梭梭种群静态生命表Tab.1 Static life table of Haloxylon ammodendron population
2.3 梭梭种群存活曲线、死亡曲线和消失率曲线
存活曲线能够反映种群个体在各个龄级的存活状况。梭梭种群存活曲线如图3 所示,呈下降趋势,参考Deevey[29]基于数学模型[30]检验存活曲线类型,如表2所示,梭梭种群的直线函数的决定系数R2(0.9592)大于幂函数R2(0.6321)和指数函数R2(0.8949),表明该存活曲线符合Deevey-Ⅱ型(对角线型),这与梭梭种群存活曲线变化幅度较小,各龄级间个体数量减少速率相差不大结果相一致,与梭梭种群年龄结构和生命表的分析结果接近。

表2 梭梭种群存活曲线的检验模型Tab.2 Statistical test model for survival curves of Haloxylon ammodendron population
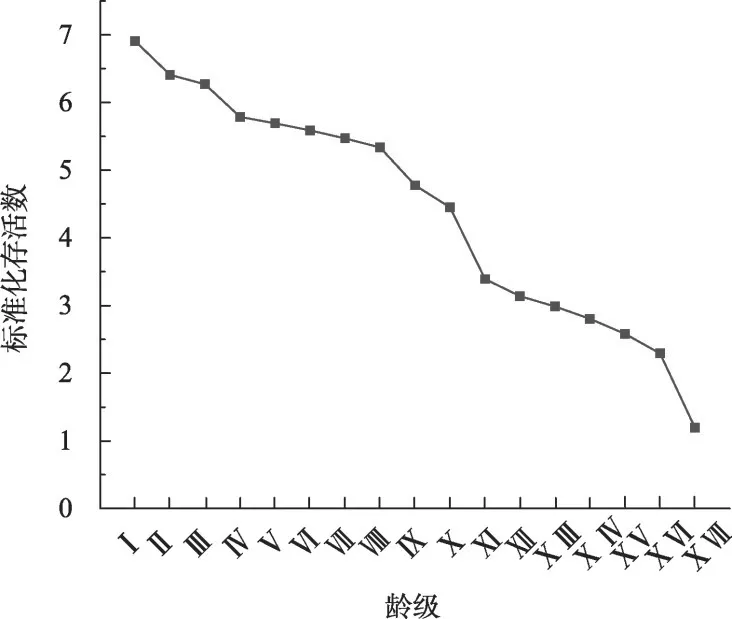
图3 梭梭种群存活曲线Fig.3 Survial curves of Haloxylon ammodendron population
梭梭种群的消失率和死亡率曲线变化趋势一致(图4),二者均呈现“降-增-降-增”的动态变化趋势。

图4 梭梭种群死亡率qx和消失率Kx曲线Fig.4 Mortality rate and vanishing rate curves of Haloxylon ammodendron population
2.4 梭梭种群动态指数
如表3 所示,种群VⅠ~VⅤ、VⅦ~VⅪ、VⅩⅢ、VⅩⅤ、VⅩⅦ大于零,VⅫ和VⅩⅥ等于零,VⅥ和VⅩⅣ小于零,表明种群动态变化为“增长-衰退-增长-稳定-增长-衰退-增长-稳定-增长”。忽略外部干扰时,梭梭种群动态指数,表明梭梭种群动态变化为增长型;考虑外部干扰时,趋近于0,说明梭梭种群增长趋势不明显且动态变化不稳定。

表3 梭梭种群动态变化指数Tab.3 Dynamic indices of Haloxylon ammodendron population
2.5 梭梭种群时间序列预测
由表4 可知,梭梭种群各龄级从Ⅱ~Ⅹ龄级,个体数量均呈现增加趋势。
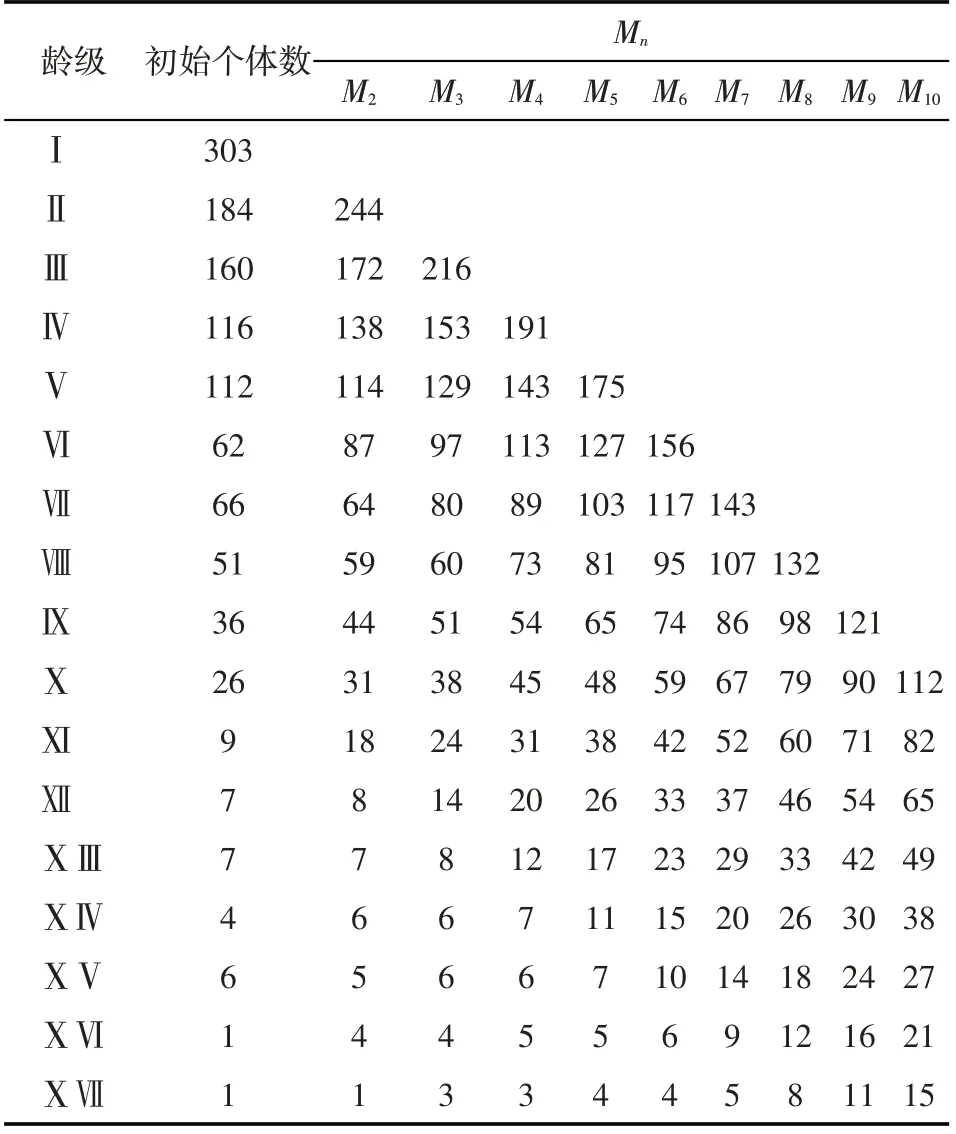
表4 梭梭种群结构与动态时间序列预测Tab.4 The prediction number of Haloxylon ammodendron population by using time sequence analysis
3 讨论
种群结构与动态特征不仅是种群个体发展的表现,同时也受外部因素的综合作用[33]。巴丹吉林沙漠中恶劣的自然条件、稀缺的水资源、动物取食以及人为因素等的强烈干扰,均是导致梭梭种群变化的重要因素。
本研究发现,巴丹吉林沙漠东缘天然梭梭种群的龄级结构属于增长型,幼苗个体数量充足,这与孙利鹏等[10]和宋于洋[34]的研究结果一致,说明梭梭种群天然更新较好且具有较强的增长潜力,随着个体增大,对资源的需求增大,当不能满足其生长时,加剧竞争,自疏作用明显。梭梭种群静态生命表也表现出与年龄结构一致的变化。这可能是由于梭梭对资源和空间持续增加的需求所致,当超过环境承载力后引发自疏和他疏作用[35]。初期,幼苗个体对资源需求较小,呈现出不明显的竞争,随着梭梭个体对资源的需求加大,梭梭种内竞争也在加剧[36]。中期,梭梭个体已有很强的生命力,受环境制约的可能性较小,少量的死亡率和消失率可能是受到外部干扰,骆驼取食或者人为采挖野生肉苁蓉所致[26]。后期,少数存活下来的梭梭开始生理衰老。梭梭种群存活曲线与李进军等[19]和杨瑞红[18]的研究结果一致,都表明种群龄级越大数量越少。梭梭种群的消失率和死亡率曲线均呈现“降-增-降增”的变化趋势,这是由于梭梭成年后对环境的适应能力增强,在某一阶段实行封育措施后,放牧对梭梭取食的影响减少,因此老龄梭梭发展较好,该阶段后梭梭接近其生理衰老年龄,死亡率缓慢增加[37]。这与本研究中梭梭种群静态生命表和梭梭存活曲线的分析结果相一致。梭梭种群动态变化指数Vpi>V′pi>0,也表现为增长型[31],说明该研究区内梭梭林天然更新良好,未发生大面积毁林现象。此外,时间序列预测分析结果也表明,梭梭种群各龄级在未来2、3、4、5、6、7、8、9、10 个龄级时间后个体数量持续增加。由此可见,该种群稳定,未来一段时间内,不考虑外界破坏其生存环境的前提下,梭梭种群天然更新稳定并能够继续保持增长[33]。
梭梭种群的发展趋势是基于其生态环境不发生变化得出的,一旦种群遭受外部干扰,其发展趋势难以预测,造成的种群破坏程度也难以想象。塔木素天然梭梭林因地处偏远,人烟稀少,且较早的实行封育禁牧措施,未遭受大面积人为破坏,但是在极端干旱的气候条件和少量外部因素(本研究指骆驼取食和人工采挖野生肉苁蓉)影响下,要使种群长期稳定发展必须制定相关保护对策。
4 结论
通过对巴丹吉林沙漠天然梭梭林的种群结构以及动态特征分析,得出以下结论:
(1)梭梭种群年龄结构属于“J”型,梭梭幼苗数量较多但成活率低,针对这一现象建议采取人工疏苗,并在极端干旱的自然环境条件下适当补充水分,水分作为植物生长的限制性因子直接决定该研究区梭梭的成活。
(2)梭梭种群存活曲线属于Deevey-Ⅱ型,龄级越大数量越少,各龄级死亡率相差不大,结合梭梭种群动态变化指数可知,该种群为稳定增长型种群,因此,建立天然梭梭林自然保护区,最大程度减少外部干扰,使得种群能够实现自我更新。
(3)时间序列预测分析结果也表明,该种群个体数量随时间推移而增加,进一步说明在该研究区设立自然保护区是必要的。
猜你喜欢
湖南生态科学学报(2022年4期)2022-11-11
河南农业大学学报(2020年2期)2020-05-22
山西林业科技(2019年1期)2019-05-27
绿色科技(2016年23期)2017-03-15
科教导刊·电子版(2016年25期)2016-11-16
江苏农业科学(2016年5期)2016-07-23
干旱气象(2016年2期)2016-06-01
文物鉴定与鉴赏(2015年9期)2015-12-07
广西农学报(2015年4期)2015-09-11
东北林业大学学报(2014年8期)2014-08-02
