
日本落叶松遗传变异分析及优良家系无性系选择
2023-08-08 01:14王芳马茂王佳兴张含国
森林工程 2023年4期
王芳 马茂 王佳兴 张含国


摘 要:为探索日本落叶松遗传变异规律及选择优良家系无性系,以内蒙古自治区赤峰市旺业甸实验林场的59个日本落叶松无性系测定林和81个日本落叶松家系子代试验林为研究对象,测定其树高,对其进行方差分析和变异分析。结果表明,日本落叶松无性系和家系树高性状存在丰富的变异,变异系数范围分别为17.84%~20.67%和24.68%~31.19%。方差分析表明,每一年,无性系间和家系间均存在显著差异,重复力和遗传力范围分别为0.482~0.494和0.793~0.819,属于中等重复力和高遗传力。以20%的入选率为标准,最终选择出12个优良无性系和16个优良家系,无性系现实增益和家系现实增益范围分别为5.73%~16.48%和7.59%~15.35%。
关键词:日本落叶松;树高;家系;无性系;变异分析
中图分类号:S791.223 文献标识码:A 文章编号:1006-8023(2023)04-0048-10
Analysis of Genetic Variation and Selection of Superior
Families and Clones of Larix kaempferi
WANG Fang1, MA Mao1, WANG Jiaxing2, ZHANG Hanguo2*
(1.Wangyedian Experimental Forestry Farm of Karqin Qi, Chifeng 024423, China;
2.State Key Laboratory of Tree Genetics and Breeding, Northeast Forestry University, Harbin 150040, China)
Abstract:In order to explore the genetic variation pattern of Larix kaempferi and to select superior family and clones, 59 clones and 81 families of Larix kaempferi in Wangyedian Experimental Forestry Farm, Chifeng City, Inner Mongolia Autonomous Region, were taken as research objects, and their tree heights were measured, and their ANOVA and variance analysis were conducted. The results showed that there was abundant variation of tree height traits in Larix kaempferi clones and families, with coefficients of variation ranging from 17.84%-20.67% and 24.68%-31.19%, respectively. ANOVA showed the difference was highly significant between clones and families every year, with a range in 0.482-0.494 and 0.793-0.819 of repeatability and heritability, respectively, which were moderate repeatability and high heritability. Using a 20% entry rate, 12 superior clones and 16 superior families were finally selected, with realistic gains of clones and realistic gains of families ranging were 5.73% to 16.48% and 7.59% to 15.35%, respectively.
Keywords:Larix kaempferi; tree height; family; clone; analysis of variance
收稿日期:2022-09-17
基金項目:国家重点研发计划项目研究任务(2022YFD2201001)
第一作者简介:王芳,高级工程师。研究方向为林木遗传育种。E-mail: 651387751@qq.com
通信作者:张含国,博士,教授。研究方向为林木遗传育种。E-mail: hanguozhang1@ sina.com
引文格式:王芳,马茂,王佳兴,等.日本落叶松遗传变异分析及优良家系无性系选择[J].森林工程,2023,39(4):48-57.
WANG F, MA M, WANG J X, et al.Analysis of genetic variation and selection of superior families and clones of Larix kaempferi[J]. Forest Engineering, 2023, 39(4):48-57.
0 引言
日本落叶松(Larix kaempferi)分布在日本本州中部地区和关东地区[1]。日本落叶松树干端直,姿态优美,叶色翠绿,适应范围广,生长初期较快,抗病性较强,木材力学性能较高,有较好的耐腐性,可做建筑材料和工业用材的原料,并可从其木材中提取松节油、酒精和纤维素等化学物品,用途很广[2-3]。日本落叶松早在20世纪中期就引入我国,东北、华北、西北乃至华中、西南地区都有栽植,其中山东、辽宁引种最早,已形成大面积人工林[4]。
变异系数大小可反映群体的变异程度,是衡量性状遗传变异潜力的有效指标[5-6]。罗芊芊等[7]在对马尾松无性系进行研究时发现,马尾松无性系生长和形质性状存在丰富的遗传差异,各性状变异系数变幅为12.44%~36.36%;肖德卿等[8]在对红豆树优树家系进行研究时发现,3年生红豆树生长和形质性状在家系间的遗传差异均达到极显著水平(P<0.01),两地点生长和形质性状的变异系数变幅为23.3%~65.1%。
增益是衡量选择效果的最重要的参数之一,反映了下一代比亲本可能增加的收获量[9]。聂林芽等[10]对杉木半同胞家系进行研究得出结论,优良家系地径遗传增益为10.17%~16.46%,家系增产效果相对显著;黄云鹏等[11]对42个4年生的福建山樱花嫁接无性系进行研究,最终选出速生福建山樱花无性系4个,树高、胸径、冠幅遗传增益均值分别为11.12%、13.35%、9.52%。
在林木良种选育过程中,优良家系选择至关重要,在选择过程中既要考虑广泛的遗传基础,又应考虑选择的遗传稳定性[12]。葛清锋等[13]以17年生日本落叶松半同胞家系为研究对象,选择出优良家系并对其早期选择结果进行验证;潘艳艳等[14]以67个日本落叶松全同胞家系为材料进行分析选择出优良家系;张正刚等[15]以36个日本落叶松自由授粉家系为研究对象进行研究,最终选择出8个优良家系。
无性系与传统的实生苗相比,具有目的性强、集约经营和生产周期短的特点,优良遗传材料的取得和大规模的无性增殖利用是无性系林业的发展基础之一[16]。杜彦昌等[17]以小陇山林区18个日本落叶松无性系为研究对象,选择出4个优良适生无性系;杜超群等[18]以11年生日本落叶松混系超级苗无性系为研究对象,开展优良无性系初步选择,最终选择出5个优良无性系。
为了获得高产稳产、遗传增益高的良种,对无性系和家系进行评价尤为重要[19]。本研究以内蒙古自治区赤峰市旺业甸林场的59个日本落叶松无性系测定林和81个日本落叶松家系子代测定林为研究对象,测量其树高,对其树高性状进行变异分析和方差分析,选择出优良家系和无性系,为日后日本落叶松优良家系无性系的大面积推广以及营造速生林提供优良材料。
1 材料与方法
1.1 试验地条件情况
旺业甸实验林场位于内蒙古自治区赤峰市喀喇沁旗西南部,地处燕山山脉北麓七老图山支脉,地理位置东经118°09′~118°30′,北纬41°21′~41°39′,海拔800~1 890 m。经营区属暖温带半干旱地区,为明显的大陆性季风气候。年降水量400~600 mm,年平均气温4.2 ℃,无霜期117 d,土壤以典型的棕色森林土为主。
1.2 试验材料
日本落叶松无性系测定林位于旺业甸实验林场美林营林区前敖包沟61林班45小班,试验材料来自日本落叶松种子园,在种子园分无性系采集穗条并嫁接育苗。2013年4月嫁接育苗,2014年4月造林,参试无性系共59个(表1),单行2株小区5次重复,株行距1.5 m×2 m,随机排列,整地方式为穴状。日本落叶松家系子代测定林位于旺业甸实验林场古山营林区南岔林班21小班,自由授粉家系子代测定种子来自日本落叶松种子园。2015年4月分家系进行田间试验育苗,2017年4月造林,参试家系共81个(表1),单行10株小区6次重复,株行距1.5 m×2 m。
1.3 試验方法
2019—2021年秋季当日本落叶松停止生长后,用塔尺对家系和无性系测定林当年树高进行测量。数据分析采用Excel和Spss 22.0统计分析软件进行分析。
以家系小区平均值为单元进行方差分析,方差分析采用线性模型
Yijk=μ+Bi+Fj+BFij+eijk
式中:Yijk为第i个区组第j个家系第k个观测值;μ为群体平均效应;Bi为第i个区组效应;Fj为第j家系的效应;BFij为第i家系和第j区组的互作效应;eijk为机误。
变异系数(CV)计算公式
CV=S/X×100%。
式中:S表示表型标准差;X表示树高性状平均值。
家系遗传力计算公式
h2=(MSf-MSe)/MSf=1-1/F。
式中:MSf表示家系均方;MSe表示误差均方;F表示方差分析中的F值。
家系遗传增益、现实增益计算公式[20]
ΔG=(Xi-X)Δh2/X×100%。
G=(Xi-X)/X×100%。
式中:ΔG为遗传增益;G为现实增益;Xi为入选家系平均值;X为家系平均值;h2为家系遗传力。
无性系重复力计算公式
R=1-1/F。
无性系遗传增益、现实增益计算公式
ΔG=RW/X×100%。
G=W/X×100%。
式中:W为选择差;R为无性系重复力。
2 结果与分析
2.1 日本落叶松无性系树高变异分析及初步选择
2.1.1 日本落叶松无性系树高变异分析
对7—9年生日本落叶松无性系进行变异分析,结果见表2。日本落叶松树高性状存在丰富的变异,并且随着树龄的增长,变异系数逐渐降低。变异系数范围在17.84%~20.67%,平均变异系数为19.05%。
对不同年度的日本落叶松各无性系进行变异分析,并列出变异系数较大前5位和较小后5位的无性系。
7年生日本落叶松无性系树高平均变异系数为20.67%,其中变异系数较高的5个无性系为81、D2、87、73081和38,平均变异系数为33.82%,比总体平均变异系数高13.15%;变异系数较低的5个无性系为73118、93、RT1、2019和S2,平均变异系数为9.29%,比总体平均变异系数低11.38%。
8年生日本落叶松无性系树高平均变异系数为18.67%,其中变异系数较高的5个无性系为95、81、D2、38和73096,平均变异系数为31.27%,比总体平均变异系数高12.60%;变异系数较低的5个无性系为41、88、94、93和RT1,平均变异系数为8.55%,比总体平均变异系数低10.12%。
9年生日本落叶松无性系树高平均变异系数为17.84%,其中变异系数较高的5个无性系为81、73113、73108、86和38,平均变异系数为27.83%,比总体平均变异系数高9.99%;变异系数较低的5个无性系为76、RB、S9、93和41,平均变异系数为9.24%,比总体平均变异系数低8.60%。
可见,日本落叶松无性系存在着丰富的遗传变异,除9年生变异较小的5个无性系平均变异系数高于8年生外,其他变异系数均随着树龄的变化而降低。3个年度共有的变异大的家系有81和38,变异小的家系有93。
2.1.2 日本无性系树高方差分析及无性系初步选择
对日本落叶松无性系各年度树高进行方差分析,结果见表3。3个年度在无性系间均表现出差异极显著,3个年度重复力分别为0.494、0.482和0.489,属于中等重复力水平,可以进行无性系选择。
入选率为10%时,遗传增益分别为7.02%、6.4%和6.72%;入选率为20%时,遗传增益分别为5.92%、5.04%和5%。通过估算遗传增益可以看出,由于总体变异系数逐年减小,遗传增益随着林龄增长而略有降低,但林龄越大,选择结果会越准确。
通过树高均值和无性系变异系数,以20%入选率为标准,对9年生日本落叶松无性系进行选择,最终选择73109、73121、D81、2019、D61、76、72、73099、82、S4、79和44号12个优良无性系,结果见表4。现实增益范围为5.73%~16.48%。
2.2 日本落叶松家系变异分析及初步选择
2.2.1 日本落叶松家系树高变异分析
对5—7年生的日本落叶松家系进行变异分析,结果见表5。与无性系相似,家系树高性状也存在丰富的变异。并且随着树龄的增长,变异系数也逐渐降低。变异系数范围为24.68%~31.19%,平均变异系数为28.16%。
对不同年度日本落叶松各家系进行变异分析,并列出变异系数较高前5和较低后5位的家系。
5年生日本落叶松家系树高平均变异系数为31.19%,其中变异系数较高的5个家系为99、44、87、88和S2,平均变异系数为37.63%,比总体平均变异系数高6.44%;变异系数较低的5个家系分别为73098、73108、76、D81和41,平均变异系数为21.44%,比总体平均变异系数低9.75%。
6年生日本落叶松家系树高平均变异系数为28.61%,其中变异系数较高的5个家系为44、87、99、88和73094,平均变异系数为34.97%,比总体平均变异系数高6.36%;变异系数较低的5个家系分别为73098、76、73108、D81和41,平均变异系数为18.08%,比总体平均变异系数低10.53%。
7年生日本落叶松家系树高平均变异系数为24.68%,其中变异系数较高的5个家系为44、87、99、73101和D41,平均变异系数为30.82%,比总体平均变异系数高6.14%;变异系数较低的5个家系分别为RT2、D81、82、S4和41,平均变异系数为15.12%,比总体平均变异系数低9.56%。
可见,日本落叶松家系存在着丰富的遗传变异,并且随着树龄的增长,变异系数逐年降低。3个年度共有的变异大的家系有44、87和99;共有的变异小的家系有41和D81。
2.2.2 日本落叶松家系树高方差分析及家系初步选择
对日本落叶松家系树高进行方差分析,结果见表6。3个年份在家系以及区组家系互作间均差异极显著。3个年度年树高性状遗传力分别为0.819、0.793和0.804,受到较强遗传控制,适合在家系水平进行遗传改良。
入选率为10%时,遗传增益分别为13.75%、11.09%和9.66%;入选率为20%时,遗传增益分别为11.74%、9.43%和8.51%。与无性系相似,总体遗传增益系数逐年减小。林龄越大,选择结果越准确。
通过树高均值和家系变异系数,以20%入选率为标准,对7年生日本落叶松家系进行选择,最终选择82、77、D82、73098、D81、D61、76、73105、RT2、74、73109、S9、81、D12、94和73108号16个优良家系,结果见表7。现实增益范围为7.59%~15.35%。
2.3 无性系与家系对比分析
2.3.1 变异对比分析
对57个具有相同系号的日本落叶松家系和无性系变异进行对比分析,结果见表8。由表8发现同一年龄下家系平均变异系数要大于无性系平均变异系数,符合有性繁殖变异较大的理论,7年生家系变异系数比无性系变异系数高2.72%。
对7年生各家系和无性系进行变异分析,结果发现排名前5的无性系变异系数大于排名前5的家系变异系数,而排名后5的无性系变异系数要小于排名后5的家系变异系数,说明家系、无性系变异较大或较小系号是不同的。排名前5的无性系平均变异系数比家系高3.22%,排名后5的无性系平均变异系数比家系低5.68%。
2.3.2入选系号及增益对比分析
对7年生日本落叶松相同家系和无性系进行方差分析,结果见表9。树高性状在家系和无性系间均差异极显著。同一年度家系遗传力大于无性系重复力。
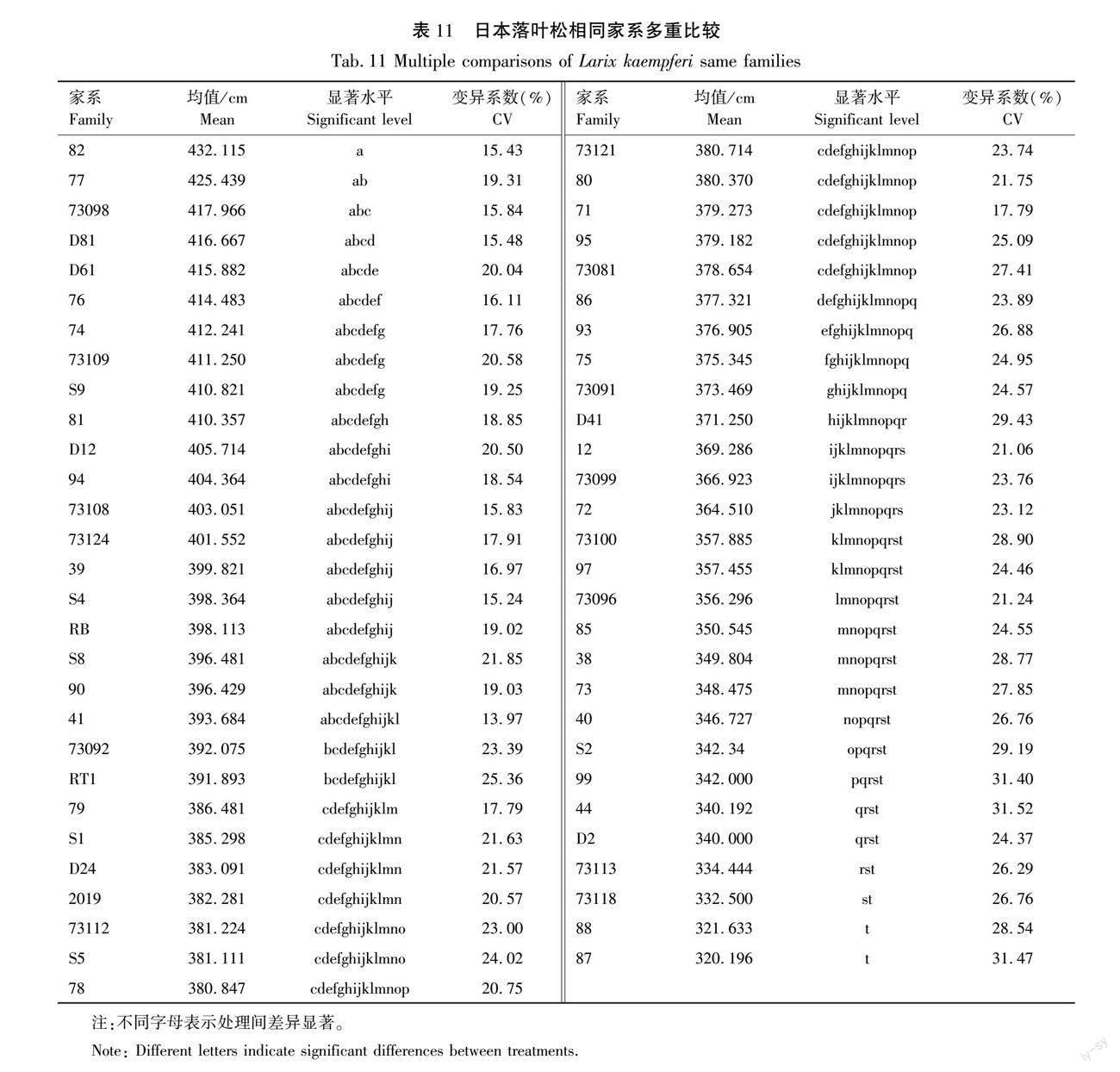
通过树高均值和变异系数,以20%入选率为标准,对日本落葉松无性系和家系进行选择,最终选择出73121、2019、73109、D81、D61、S2、D24、72、82、S4和76号11个优良无性系和82、77、73098、D81、D61、76、74、73109、S9、81和94号11个优良家系,结果见表10和表11。无性系现实增益为7.62%~16.09%;家系现实增益范围为6.30%~13.60%。
從结果上看,选出的优良家系和优良无性系相同系号有82、73109、D81、D61号4个系号,说明这4个系号有性繁殖和无性繁殖均能获得较大增益。
3 结论和讨论
林木的遗传变异是遗传改良的前提,是遗传信息的重要表征。丰富的遗传变异决定了物种在育种过程中的改良潜力,掌握林木的遗传变异规律,是育种策略制定的基础[21-23]。本研究结果显示,日本落叶松无性系和家系树高性状存在丰富的变异,7—9年生无性系变异系数范围为17.84%~20.67%,平均变异系数为19.05%;5—7年生家系变异系数范围为24.68%~31.19%,平均变异系数为28.16%。研究发现,无论是无性系还是家系,树龄越大,变异系数越小,这与马茂等[24]对日本落叶松无性系和缪小飞等[25]对杂种落叶松家系的研究结果一致。同一年龄相同系号的日本落叶松家系和无性系变异对比分析,结果显示家系平均变异系数要大于无性系平均变异系数,而且家系、无性系变异较大或较小系号是不同的。
在林木育种研究中,方差分析是评估变异幅度的重要方法[26]。方差分析结果显示,3个年份树高在无性系间和家系间差异极显著。遗传力和重复力是衡量某一性状在不同时间、不同地点其表型特征是否可以持续稳定遗传的判定指标,遗传力(重复力)越高,说明受外界环境影响越小[27,28]。3个年度树高性状无性系重复力范围为0.482~0.494,属于中等重复力,表现较为稳定。3个年度家系遗传力范围为0.793~0.819,属于高遗传力,受到较强遗传控制。相同材料同一年度家系遗传力大于无性系重复力。
日本落叶松无性系入选率为20%时,3个年度树高性状遗传增益分别为5.92%、5.04%和5%。家系入选率为20%时,3个年度遗传增益分别为11.74%、9.43%和8.51%。由于试验材料的遗传差异随着年龄增长会发生变化,因此选择树龄较大的材料,选择的结果会相对准确一些。
通过树高平均值和变异系数,以20%入选率为标准,最终选择出12个优良无性系和16个优良家系,其中D81、D61、76、73109、76、82号为无性系和家系共有系号,表明这6个系号有性繁殖和无性繁殖均能获得较大增益。通过对比共有家系和无性系,最终选择出包括S2、D24、71号在内的15个无性系和16个家系。
【参 考 文 献】
[1]中国科学院中国植物志编辑委员会.中国植物志[M].北京:科学出版社,1978.
Editorial Committee of China Plant Journal, Chinese Academy of Sciences. China plant journal[M]. Beijing: Science Press, 1978.
[2]LAI M, SUN X M, CHEN D S, et al. Age-related trends in genetic parameters for Larix kaempferi and their implications for early selection[J]. BMC Genetics, 2014, 15(1):1-8.
[3]FUKATSU E, TSUBOMURA M, FUJISAWA Y, et al. Genetic improvement of wood density and radial growth in Larix kaempferi: result from a diallel mating test[J]. Annals of Forest Science, 2013, 70:451-459.
[4]滕秀兰,王福森,李树森,等.日本落叶松品种引进及育苗造林技术探析[J].防护林科技,2018(1):91-92.
TENG X L, WANG F S, LI S S, et.al. Introduction of Larix kaempferi species and seedling planting technology[J]. Protection Forest Science and Technology, 2018(1):91-92.
[5]LEPOITTEVIN C, ROUSSEAU J P, GUILLEMIN A, et al. Genetic parameters of growth, straightness sand wood chemistry traits in Pinus pinaster[J]. Annals of Forest Science, 2011, 68(4):873-884.
[6]高玉红,魏萌,李凯迪,等.不同红河橙居群果实表型的变异[J].经济林研究,2021,39(4):88-96.
GAO Y H, WEI M, LI K D, et al. Phenotypic variation of Citrus hongheensis fruit from different provenances[J]. Non-wood Forest Research, 2021, 39(4): 88-96.
[7]罗芊芊,肖德卿,徐洪辉,等.马尾松无性系生长和形质性状变异分析和优选[J].东北林业大学学报,2022,50(9):40-44.
LUO Q Q, XIAO D Q, XU H H, et al.Variation analysis and optimization of growth and form-quality traits of Pinus massoniana clones[J]. Journal of Northeast Forestry University, 2022, 50(9):40-44.
[8]肖德卿,邓章文,罗芊芊,等.幼龄红豆树生长和形质性状家系变异分析[J].林业科学研究,2021,34(3):152-157.
XIAO D Q, DENG Z W, LUO Q Q, et al.Growth and form-quality traits diversity analysis of young Ormosia hosiei family[J]. Forest Research, 2021, 34(3):152-157.
[9]RUOTSALAINEN S, LINDGREN D. Predicting genetic gain of backward and forward selection in forest tree breeding[J]. Silvae Genetica,1998, 47(1):42-50.
[10]聂林芽,曾素平,厉月桥,等.杉木半同胞家系生长性状遗传分析及优良家系选择[J].林业科技通讯,2022(10):3-9.
NIE L Y, ZENG S P, LI Y Q, et al. Genetic analysis of growth traits and selection of excellent families in half sib families of Cunninghamia lanceolata[J]. Forest Science and Technology, 2022(10):3-9.
[11]黄云鹏,丁珌,潘惠忠,等.福建山樱花无性系生长性状遗传变异分析及选择[J].福建林业科技,2022,49(2):7-10,25.
HUANG Y P, DING M, PAN H Z, et al. Genetic variation of growth trait and superior clone selection of Cerasus campanulata[J]. Joural of Fujian Forestry Science and Technology, 2022, 49(2):7-10, 25.
[12]潘艳艳.日本落叶松种子园亲本及其子代变异研究[D].哈尔滨:东北林业大学,2019.
PAN Y Y. Study on genetic variation of parents and progenies in Larix kaempferi seed orchard[D]. Harbin: Northeast Forestry University, 2019.
[13]葛清锋,曾美,侯义梅.日本落叶松半同胞家系生长变异研究[J].湖北林业科技,2022,51(3):6-9,21.
GE Q F, ZENG M, HOU Y M. Study on growth variation of Larix kaempferi half-sib families[J]. Hubei Forestry Science and Technology, 2022, 51(3):6-9, 21.
[14]潘艳艳,许贵友,董利虎,等.日本落叶松全同胞家系苗期生长性状遗传变异[J].南京林业大学学报(自然科学版),2019,43(2):14-22.
PAN Y Y, XU G Y, DONG L H, et.al. Genetic variations of seedling growth traits among full-sib families of Larix kaempferi[J]. Journal of Nanjing Forestry University (Natural Sciences), 2019, 43(2):14-22.
[15]张正刚,马建伟,靳新春,等.日本落叶松 [J].西北林学院学报,2013,28(4):74-79.
ZHANG Z G, MA J W, JIN X C, et al. Analysis and selection of progeny test forest of the open pollinated family of Larix kaempferi[J]. Journal of Northwest Forestry University, 2013, 28(4):74-79.
[16]DONALDSON L, BURDON R. Clonal variation and repeatability of micro fibrilangle in Pinus radiata[J]. New Zealand Journal of Forestry Science, 1995, 25(2):164-174.
[17]杜彥昌,马建伟,安三平,等.小陇山林区日本落叶松无性系遗传变异及初选[J].林业科技通讯,2018(7):11-15.
DU Y C, MA J W, AN S P, et al. Genetic variation and primary selection of clones of Larix kaempferi in Xiaolong mountain forest area[J]. Forest Science and Technology, 2018(7):11-15.
[18]杜超群,许业洲,孙晓梅,等.鄂西亚高山区日本落叶松无性系生长性状变异分析与早期选择[J].华中农业大学学报,2015,34(3):19-23.
DU C Q, XU Y Z, SUN X M, et al. Variation of growth traits and early selection of Larix kaempferi clones in sub-alpine area of western Hubei Province[J]. Journal of Huazhong Agricultural University, 2015, 34(3):19-23.
[19]李岩,朱嘉瑶,王喜和,等.红松优树无性系及其子代的生长评价与选择研究[J].北京林业大学学报,2021,43(10):38-46.
LI Y, ZHU J Y, WANG X H, et al. Growth evaluation and selection study of elite clones and its offspring families in Pinus koraiensis[J]. Journal of Beijing Forestry University, 2021, 43(10):38-46.
[20]欧阳天林,朱柯帆,邱建勋,等.木荷种子园自由授粉家系生长遗传变异及初选[J/OL].中南林业科技大学学报,2022(9):17-23.
OUYANG T L, ZHU K F, QIU J X, et al. Genetic variation and selection of the free-pollinated families in Schima superba seed orchard[J]. Journal of Central South University of Forestry & Technology, 2022(9):17-23.
[21]王云鵬,张蕊,周志春,等.木荷优树自由授粉家系早期生长性状遗传变异动态规律[J].林业科学,2020,56(9):77-86.
WANG Y P, ZHANG R, ZHOU Z C, et al. Dynamic patterns of genetic variation in early growth traits of the open-pollinated families of Schima superba plus tree[J]. Scientia Silvae Sinicae, 2020, 56(9):77-86.
[22]王玮槐,陈雨桐,党乾顺,等.红松无性系和优树种子性状及营养成分分析[J].森林工程,2019,35(2):11-20,26.
WANG W H, CHEN Y T, DANG Q S, et al.Analysis of characters and nutritional components of Pinus koraiensis seeds on clones and superior trees[J]. Forest Engineering, 2019, 35(2):11-20, 26.
[23]周燕,黄盛怡,杨孟晴,等.赤皮青冈不同家系苗期生长和叶片性状遗传变异[J].西部林业科学,2022,51(5):75-80.
ZHOU Y, HUANG S Y, YANG M Q, et al. Genetic variation of growth and leaf traits of different Cyclobalanopsis gilva families at seedling stage[J]. Journal of West China Forestry Science, 2022, 51(5): 75-80.
[24]马茂,徐开源,张含国.日本落叶松无性系树高变异与初步选择[J].林业科技,2019,44(5):5-7.
MA M, XU K Y, ZAHNG H G. Tree height variation and preliminary selection of clones of Larix kaempferi[J]. Forestry Science & Technology, 2019, 44(5):5-7.
[25]缪小飞,张含国,侯丹,等.杂种落叶松家系遗传变异及多点稳定性[J].东北林业大学学报,2018,46(12):1-8.
MIU X F, ZHANG H G, HOU D, et al. Genetic variation and multi-point stability of families of Hybrid larch[J]. Journal of Northeast Forestry University, 2018, 46(12):1-8.
[26]SAFAVI S A, POURDAD S A, MOHMMAD T, et al. Assessment of genetic variation among safflower (Carthamus tinctorius L.) accessions using agro-morphological traits and molecular markers[J]. Journal of Food Agriculture & Environment, 2010, 8(3):616-625.
[27]刘明国,张欣,董胜君,等.西伯利亚杏优选无性系间数量性状的差异与重复力研究[J].沈阳农业大学学报,2015,46(5):548-554.
LIU M G, ZHANG X, DONG S J, et al. Differences and repeatabilities of quantitative characters of Prunus sibirica superior clones[J]. Journal of Shenyang Agricultural University, 2015, 46(5):548-554.
[28]史加亮,李凤瑞,赵文超,等.陆地棉特异种质主要性状配合力和遗传力分析[J].山东农业科学,2021,53(1):14-19.
SHI J L, LI F R, ZHAO W C, et al. Analysis on combining ability and heritability of main traits for special heteroplasm in upland cotton[J]. Shandong Agricultural Sciences, 2021, 53(1):14-19.
猜你喜欢
中华实用诊断与治疗杂志(2022年1期)2022-08-31
农家科技中旬版(2017年4期)2017-05-11
农民致富之友(2017年4期)2017-04-10
现代农业科技(2017年4期)2017-04-10
绿色科技(2017年1期)2017-03-01
安徽农学通报(2016年12期)2016-07-09
广东海洋大学学报(2015年4期)2016-01-13
听力学及言语疾病杂志(2015年5期)2015-12-24
首都医科大学学报(2015年4期)2015-12-16
福建林业科技(2015年2期)2015-09-16
