
短枝木麻黄半同胞家系苗期遗传变异及选择
2023-09-20 11:23徐惠昌聂森余锦林尤龙辉叶功富张航晓程分生苏亲桂
热带作物学报 2023年8期
关键词:遗传变异
徐惠昌 聂森 余锦林 尤龙辉 叶功富 张航晓 程分生 苏亲桂
关键词:短枝木麻黄;半同胞家系;遗传变异;苗期选择
短枝木麻黄生长迅速,具有良好的抗风沙、耐旱及耐盐碱等特性,是我国东南沿海沙地最重要的生态防护树种之一[1-2]。20世纪80年代营造的木麻黄防护林,因林分逐渐衰退更新,而被短枝木麻黄优良无性系惠1、闽平2号、木麻黄粤501等无性系所替代,但是由于无性系品种来源单一、种质资源贫乏、遗传基础狭窄,导致林分抗性下降、易感病虫害等问题逐渐凸显,木麻黄防护林生态防护功能不断下降[3-4]。因此,木麻黃实生种子苗所具有的生长期长、根系发达、抗性强等优点得到相关研究者、林业生产和管理单位重新审视和重视。
“有性选育,无性利用”是木麻黄遗传改良的指导方针[5-6]。我国自20世纪80年代中期,以中澳、中法等国际合作项目为契机,先后引进短枝木麻黄不同地区国际种源73个[7],并于福建、广东和海南等地建立了多个种源试验点,从而揭开了国内短枝木麻黄种源试验的序幕,并对其遗传变异规律进行了较为全面、系统的研究,选育出了多用途的优良种源[2]。但是,关于短枝木麻黄优良家系遗传选育的研究却较为滞后。傅玉狮[8]于1996年,通过对34个短枝木麻黄半同胞家系的11年生子代测定林进行分析,结果表明其生长和抗性性状遗传变异丰富,并筛选出8个速生丰产的优良家系,其在一定程度上丰富了短枝木麻黄优良种质资源。然而,此后关于短枝木麻黄家系遗传改良育种工作的研究鲜有报道。
亚群体层次(如家系层次)遗传改良相较于群体层次,可以消除后者对有益配子不平衡的负面效应,并避免加性基因方差的下降,提高子代育种值,林业发达国家在采用现代苗木生产技术的同时,也正从种源层次的苗木培育逐步转为家系层次育种,以充分发挥亚群体层次遗传改良的潜在效益[9]。鉴于良种选育和苗木培育对发掘变异、研究变异、利用变异的要求,我国林木育种工作者对红皮云杉[10]、云南松[11]等树种进行了大量研究,并证实在苗期进行遗传改良选择,在提高林木良种选育效率、加速育种进程的同时,还可获得较高的增益。故本研究以40个短枝木麻黄半同胞家系1年生幼苗为研究对象,结合前人对短枝木麻黄的相关研究[2,12],观测分析其苗期苗高、地径、干重、根冠比、侧枝数、侧枝角及侧枝长等生长与分枝性状的遗传变异,探寻短枝木麻黄家系层次的苗期遗传变异规律,以期为后续木麻黄科树种更好地开展家系层次遗传改良工作提供参考依据。
1材料与方法
1.1材料
1.1.1试验地点试验地位于福建省惠安赤湖国有防护林场后山工业区(118°54′29″E,24°54′45″N),平均海拔约12m。属南亚热带海洋性季风气候,干湿季明显,夏季多台风,秋冬盛行东北风,该区域年均气温为20.7℃,年均降雨量1029.0mm,年均蒸发量达2056.7mm。试验地设在林场场部苗圃,基质土壤类型为沙壤土,肥力较低,日照充足。
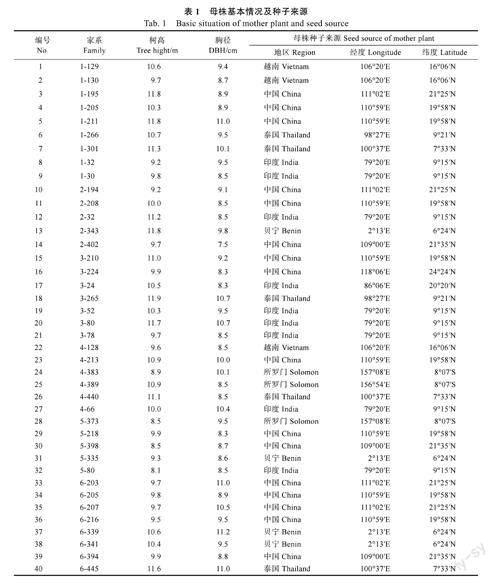
1.1.2试验材料2019年8月,对短枝木麻黄国际种源试验林(5年生)的表型性状进行观测分析,筛选出树高、胸径生长势及健康状况良好的优良母株进行采种、保存。2021年3月取出种子,于林场苗圃进行播种育苗,共计培育40个短枝木麻黄半同胞家系(表1)。2021年5月,当幼苗长至10cm左右时,按照完全随机区组设计,采用双列10株小区排列方式,将幼苗移栽至塑料营养钵(规格为8cm×15cm),一株一钵,基质为沙壤土,共4个区组(重复)。适时进行浇水、除草等常规育苗管理。
1.2试验指标观测
2022年3月对短枝木麻黄40个半同胞家系幼苗的生长和分枝性状进行测定。测定指标包括苗高(SH)、地径(GD)、干重(WD)、根冠比(R/S)、侧枝数(LBN)、侧枝角(LBA)、侧枝长(LBL)等7个指标。其中苗高、地径分别采用卷尺和电子游标卡尺进行测量;侧枝角和侧枝长则采用量角器和直尺进行测量。根据测量结果,计算各区组每个家系平均苗高和平均地径;各区组每个家系再随机选取4株幼苗,清理根部泥土后,分根、茎、小枝带回实验室,置于鼓风干燥箱中,105℃烘至恒重并称重,计算根冠比=地下部分干重/地上部分干重。
1.3数据处理
采用Excel2020和SPSS25.0软件进行数据整理和统计分析;使用R语言程序包“HalfsibMS”计算遗传参数[13];利用Origin2021进行聚类分析和增益柱状图的绘制。
1.3.1生长与分枝性状遗传力、遗传变异系数、相关系数和增益的数学模型[14-15]
1.3.2短枝木麻黄优良家系的选择通过聚类分析和布雷金多性状综合评价法开展优良家系的综合选择。布雷金多性状综合评价法数学模型为[16]:
2结果与分析
2.1生长与分枝性状变异
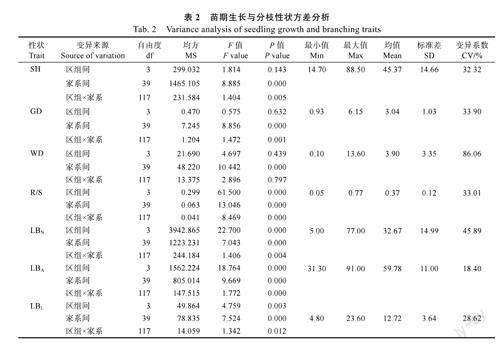
由表2可知,7个苗期性状变异系数为18.40%~86.06%,表明参试群体生长与分枝性状分化较明显。方差分析结果则显示,40个短枝木麻黄半同胞家系间苗期各生长和分枝性状差异均极显著(P<0.01,下同),且方差来源以家系为主。
2.2生长与分枝性状的遗传参数估算
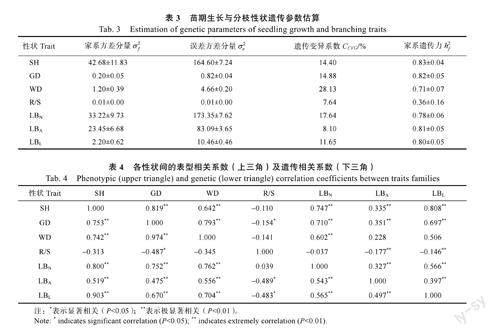
如表3所示,参试群体生长与分枝性状的遗传变异系数为7.64%~28.13%,其中根冠比和侧枝角的遗传变异系数均低于10%。在遗传力方面,各表型性状的家系遗传力变化幅度为0.36~0.83。
2.3生长与分枝性状的表型和遗传相关分析
对短枝木麻黄半同胞家系苗期生长与分枝性状,进行表型和遗传相关分析,结果(表4)表明,根冠比、侧枝角与其他表型性状指标相关性不明显或呈弱相关关系。苗高、地径、干重、侧枝数及侧枝长相互间表型和遗传相关性较高,且均达到极显著水平。
2.4优良家系选择
由于根冠比、侧枝角的遗传变异系数较低,因此,仅以苗高、地径、干重、侧枝数和侧枝长这5个性状作为评价指标,对参试家系进行聚类分析,划分类群(图1),再计算各类群的遗传增益和现实增益。
图1显示,当欧式距离达到15时,可将参试家系划分为4个类群:其中第Ⅰ类群包含1-129、2-402、2-32等10个家系;第Ⅱ类群则包含1-211、3-80、1-32和3-224等4个家系;第Ⅲ类群包含1-130、1-30、2-194等15个家系;第Ⅳ类群则包含1-205、4-128、2-343等11個家系。图2显示,第Ⅰ、Ⅱ类群家系苗期生长和分枝性状的现实增益和遗传增益均为负值,因此将其进一步归为苗期慢生家系;第Ⅲ类群家系除干重的现实增益和遗传增益高于10%,其余4个性状的现实增益为2.01%~7.86%,遗传增益为1.57%~6.44%,将其归为苗期较速生家系;第Ⅳ类群家系与参试群体相比,其侧枝长的现实增益和遗传增益分别为6.28%和5.02%,而苗高、地径、干重和侧枝数的现实增益和遗传增益均超过10%,可归为苗期速生家系。
以苗高、地径、干重、侧枝数及侧枝长为评价指标,采用布雷金多性状综合评价法对参试家系进行评价(表5)。按10%的入选率进行筛选,共有4个家系入选,分别为3-265、5-373、1-266和2-343家系,其苗高、地径、干重、侧枝数和侧枝长平均值分别为54.59cm、3.61mm、5.48g、39.04和15.10cm,现实增益分别为21.14%、19.10%、41.83%、19.97%和18.88%,遗传增益分别为17.55%、15.66%、29.70%、15.57%和15.10%。
3讨论
遗传和变异是林木遗传改良的重要基础[17]。遗传力表示亲本将某性状遗传给子代的能力,遗传力愈大,愈能稳定遗传给子代[18-19]。本研究中,苗高、地径、根冠比、侧枝数、侧枝角及侧枝长等性状的家系遗传力为0.36~0.83,在家系水平上,受到中度或强度遗传控制[20],显著高于PRASAD等[21]在印度安得拉邦对短枝木麻黄家系苗期生长性状遗传力的估算结果。这可能是因为短枝木麻黄花期以3—5月为主,而受西南暖湿气流与冷空气交绥作用的影响,供试样地此时降雨较为频繁,在一定程度上阻碍了花粉传播,同一母株的种子材料中可能包含较大比例的全同胞子代,间接导致参试家系的遗传力有所提高[22-24]。此外,方差分析结果显示,各性状在家系间差异极显著,变异系数为18.40%~86.06%,这与胡盼等[12]对幼苗时期不同种源的短枝木麻黄表型性状差异评价结果及陈珍等[25]对25个短枝木麻黄无性系进行苗期选择研究的结果相似,表明短枝木麻黄生长和分枝性状分化明显,变异较丰富,具有较高的遗传改良潜力。
遗传变异系数反映了性状遗传变异的相对大小,其数值越大,选取获得优良遗传型的潜力越大[26]。本研究中,参试群体根冠比和侧枝角的遗传变异系数较低,仅为7.64%和8.10%,表明这2个性状的选择潜力较低。相关系数代表了性状间的关联程度,可用来确定目的性状之间的相关关系[27-28]。本研究发现,侧枝数、侧枝长与苗高、地径、干重间的表型相关性和遗传相关性均达到了极显著水平,这与胡盼等[12]的研究结果相似,说明侧枝数和侧枝长可作为短枝木麻黄优良半同胞家系筛选的参考因子。
遗传增益是人工选择群体所取得改良效果的重要评价指标,受到遗传力、变异系数和选择强度的直接影响[24]。本研究以苗高、地径、干重、侧枝数和侧枝长作为评价指标,结合聚类分析和多性状综合评价法,以10%入选率,筛选出4个优良家系,增益明显,选择效果较为显著。然而,隶属于较速生类群的1-266入选其中,可能是因为该家系平均侧枝长较大,因此,在苗期选择时,也可适当考虑第Ⅲ类群的较速生家系。入选优良家系为后续短枝木麻黄遗传改良和林分营建提供了较好的种质材料。但受基因与环境的共同作用,林木在不同生长发育阶段,其遗传和表型参数是动态变化的[29],故今后还需结合造林试验进行多年观测,进一步探究短枝木麻黄半同胞家系的变异及其稳定性,以便更加科学地指导木麻黄育种和营林生产实践。
猜你喜欢
中国农业科学(2022年14期)2022-07-26
园林科技(2021年2期)2022-01-19
海南医学(2020年1期)2020-01-18
电子制作(2019年24期)2019-02-23
森林工程(2018年3期)2018-06-26
广东农业科学(2016年7期)2016-10-08
广西林业科学(2016年3期)2016-03-16
广西林业科学(2016年3期)2016-03-16
中国康复理论与实践(2015年10期)2015-12-24
动物医学进展(2015年10期)2015-12-07
