
基于同质园栽培的21种淫羊藿属植物花部特征的分类学意义
2023-09-21 01:09易雪倩舒为杰杨晓琛
西北植物学报 2023年8期
易雪倩,何 花,曾 罗,舒为杰,杨晓琛
(吉首大学 生物资源与环境科学学院 湖南吉首 416000)
花是被子植物有性繁殖的重要器官,花部特征被认为是对不同传粉媒介的适应,是植物与环境相互作用,长期进化的产物[1],多样性极为丰富且相对稳定[2]。因此,花部形态特征常被用作被子植物分类和系统演化的重要依据[3]。
依据形态学对植物的分类研究多以叶形态[4]、花部形态[5]、花粉形态[6]、果实或种子形态[7-8]等性状作为分类依据,利用上述性状的种间差异将物种进行区分。这类研究的采样主要基于野外自然居群[9-10]或同质园栽培居群[11-12]。基于野外采样所观测性状的差异是建立在植物所在生境与遗传物质共同作用的基础之上,会导致形态差异的来源不明确。然而,同质园实验(common garden experiment)是将不同生境居群的个体或不同物种引种栽培在环境因素相对一致的同一园地内[13-14],可以排除环境因素对表型变异的影响,从而单独考察遗传因素的影响;如果形态差异经同质园栽培后仍然存在,则可以认定这种差异来源于遗传分化[15]。
淫羊藿属(Epimedium)为小檗科(Berberidaceae)最大的草本属,全世界约68种,其中85%以上的物种为中国特有[16-17]。该属植物最原始种类出现在东亚,中国为该属植物的多样性中心和地理分布中心,具有显著资源优势[18-19]。淫羊藿具有增强肾功能、强健筋骨、祛风除湿的功效,在中国入药历史悠久[20]。
Stearn[18]根据叶和花的形态、地理分布和染色体的c带特征,将淫羊藿分为2个亚属、4个组和4个系,是目前较为广泛认可的淫羊藿属分类学专著,但是该属属下分类和种的范围确定十分困难。
因此,分类学家们一直致力于该属植物各类性状的分析研究,以求更好解决其分类鉴定问题。花粉形态[21]、核型分析[22]、化学成分[23]都曾用于淫羊藿属分类学研究,但结果均不理想,不能很好地区分淫羊藿属内物种。而基于ITS序列[24-25]、atpB-rbcL序列[26]、matK序列[27]以及简化基因组序列[28]的系统发育研究也不能完全将淫羊藿属植物进行分类。
先前淫羊藿属基于形态的分类主要以花大小、花瓣长短、花序梗有无以及花茎上叶的形态、数目为依据,缺乏对萼片、雌雄蕊等详细花部性状分类意义的探讨。
因此,为了明确淫羊藿属物种之间的分类关系,本研究以同质园栽培的21种淫羊藿为研究材料,通过观测这些物种的花部特征等性状,旨在回答以下问题:(1)基于同质园栽培下的淫羊藿属植物的花部特征在该属中是否具有分类学意义?(2)哪些特征在分类中起主要作用?
对上述问题的回答不仅能为淫羊藿属植物的分类、系统演化关系和药材鉴定提供基础数据,还能为淫羊藿种质资源的保护与开发利用提供参考依据。
1 材料和方法
1.1 研究地点与材料
本研究所有观测均在湖南省湘西州吉首市的吉首大学淫羊藿属植物种质资源圃(23°17′92″N,104°80′36″E,海拔195 m)中进行。21种淫羊藿于2014-2015年从野外原生境移栽至资源圃,各物种在资源圃中长势良好,每年均能正常开花结实。研究物种的具体信息见表1。
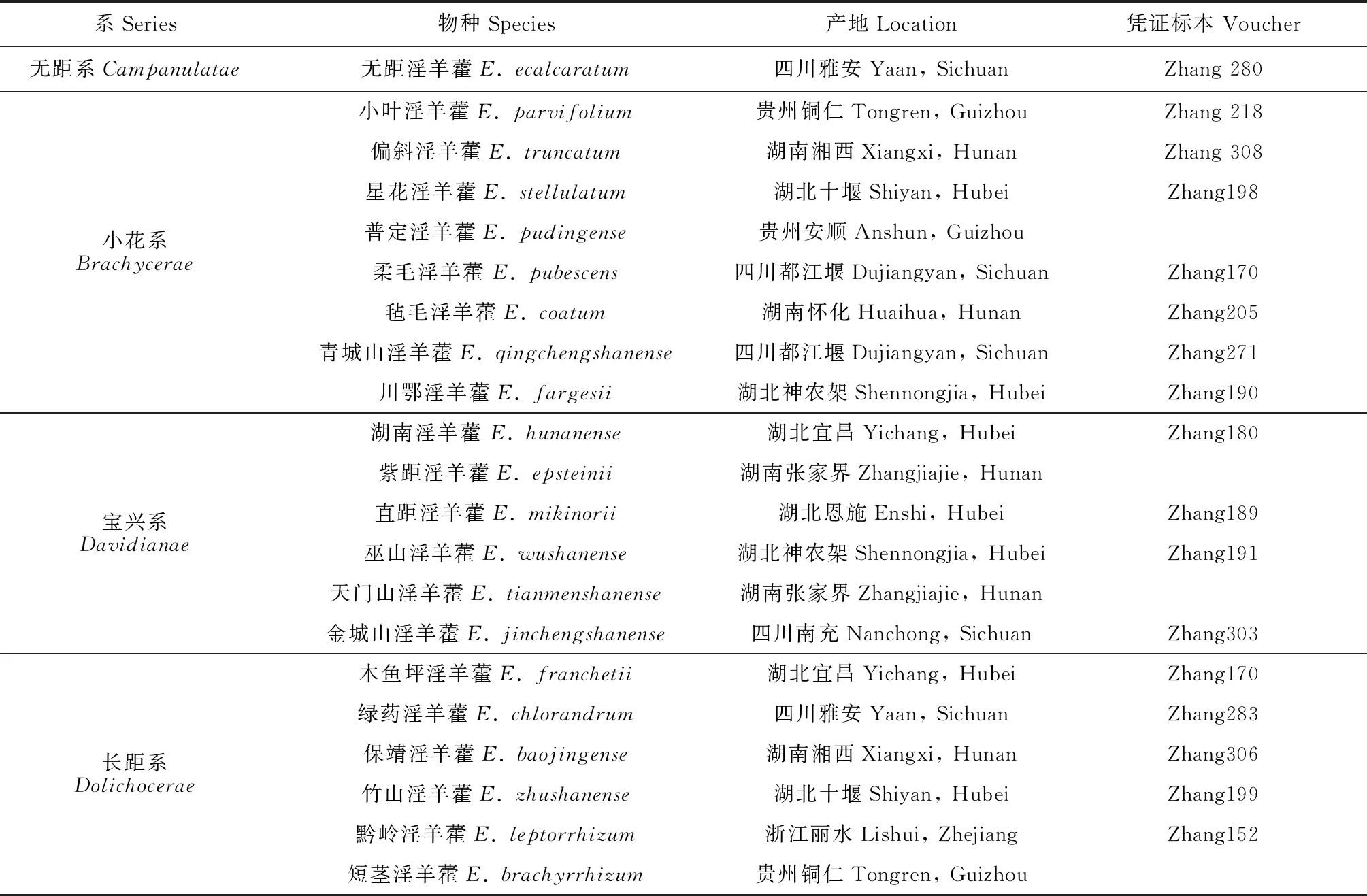
表1 试验物种及其产地信息[18,28]
1.2 花部特征观测
于各物种的盛花期,在花序的不同部位随机标记20朵即将开放的花,记录这些花开放及凋谢的时间,计算出物种单花寿命(以花冠打开为花开放标志,以花冠脱落为花凋谢标志)。
每物种从花序的不同部位随机采集20朵完全开放的花,对22个性状指标进行观测:用精度为0.01 mm的数显游标卡尺测量花梗长、花直径、内萼片长宽、蜜距长、蜜距口径、雌雄蕊长度、花药长、花柱长、柱头和蜜距口的高度差(以柱头伸出花冠记为“+”,柱头隐藏于花冠记为“-”)、花药与柱头的高度差等特征(部分观测部位说明见图1);同时观察以下6个质量性状:花序类型、有无蜜距、花瓣颜色、内萼片颜色、内萼片与花瓣相对长度和花粉颜色;在体式显微镜(Motic SMZ-171, MOTIC Inc., China)下对子房进行解剖统计胚珠数;在光学显微镜(Lecia D6200, LECIA Inc., Gremany)下统计各物种花粉粒数量并测量水合花粉粒直径。

A. 小花类群的花;B. 大花类群的花。1. 蜜距口径;2. 蜜距长;3. 内萼片长;4. 内萼片宽;5. 柱头和蜜距口的高度差;6. 雄蕊长;7. 花药与柱头的高度差;8. 花药长。
1.3 数据分析
数据分析与比较均在SPSS 26.0 (Statistical Product and Service Solutions, IBM Inc., New York, USA)中进行。测量得到的花部特征数值通过描述统计进行初步分析;而后用广义线性模型(generalized linear models)中正态分布(normal distribution)恒等关联函数(identity-link function)分析比较21个物种种间花部特征的差异。
最后,对22个花部特征性状进行聚类分析:数量性状以原始数据的平均数进行直接赋值,质量性状根据每一性状的不同类型进行编码(表2)。
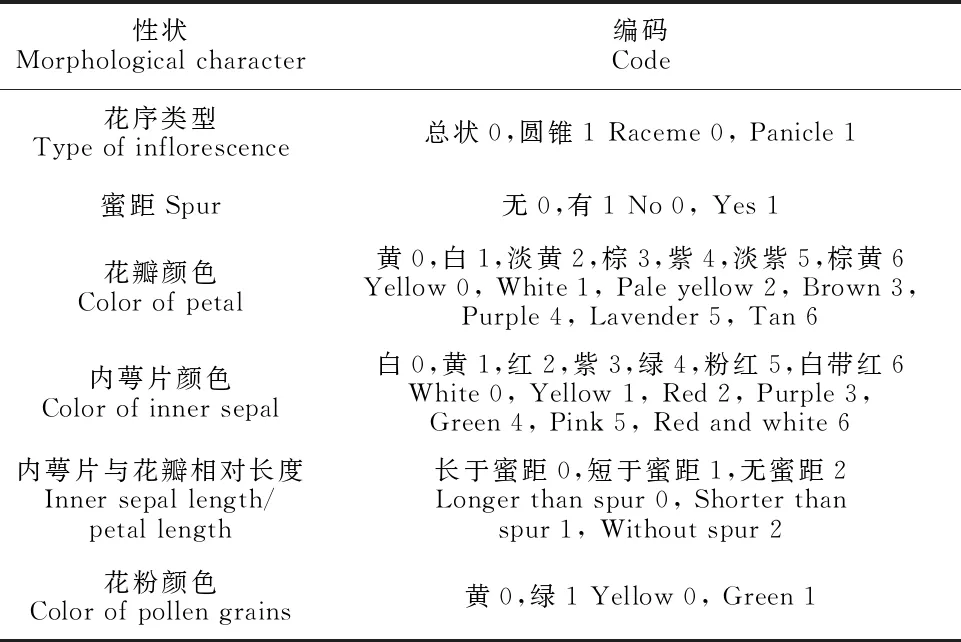
表2 淫羊藿属植物花部特征质量性状编码
聚类分析时首先要数据标准化,消除不同量纲对数据分析产生的影响,再对标准化数据计算分类单位之间的平均欧式距离系数,然后采用UPGMA法进行聚类分析,最后作出分类单位聚类结果的树形图,即亲缘关系表征图。主成分分析要在标准化的基础上计算分类单位之间的相关系数,然后求出相关系数的特征值和特征向量,最后得到特征性状在前6主成分中的负荷量。
本研究依据所观察到的花部形态特征编写了21种淫羊藿属植物分种检索表。由于实验资源圃中无距淫羊藿、天门山淫羊藿、巫山淫羊藿、偏斜淫羊藿、小叶淫羊藿的居群数量较少,在亲自测得实验数据的基础之上,参考了中国植物志(http://www.iplant.cn/frps)、中国数字植物标本馆(https://www.cvh.ac.cn/)以及相关文献中对以上物种的特征描述。基于此,也对其他物种的特征值进行了校正,确保检索表的准确性[29-32]。
2 结果与分析
2.1 花部特征
同质园栽培的21种淫羊藿的花寿命均存在显著差异(P<0.01):单花寿命2~7 d,平均(3.6±0.22) d。其中直距淫羊藿花寿命最短,仅(2.10±0.10) d,湖南淫羊藿花寿命最长:(6.40±0.40) d。淫羊藿花两性,圆锥花序或总状花序,花直径5.14~43.46 mm,内轮萼片花瓣状,花瓣4,特化成蜜距(无距淫羊藿除外)(图2),花瓣颜色和内萼片颜色多变(图3、表3)。

A.黔岭淫羊藿;B.短茎淫羊藿;C.湖南淫羊藿;D.竹山淫羊藿;E.直距淫羊藿;F.巫山淫羊藿;G.紫距淫羊藿;H.绿药淫羊藿;I.木鱼坪淫羊藿;J.保靖淫羊藿;K.金城山淫羊藿;L.无距淫羊藿;M.青城山淫羊藿;N.川鄂淫羊藿;O.柔毛淫羊藿;P.星花淫羊藿;Q.小叶淫羊藿;R.毡毛淫羊藿;S.偏斜淫羊藿;T.普定淫羊藿;U.天门山淫羊藿。
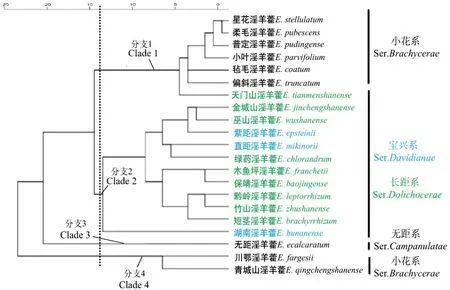
横向距离表示差异的大小,树状结构则表示分类情况,右侧系依据Stearn[18]的分类系统。
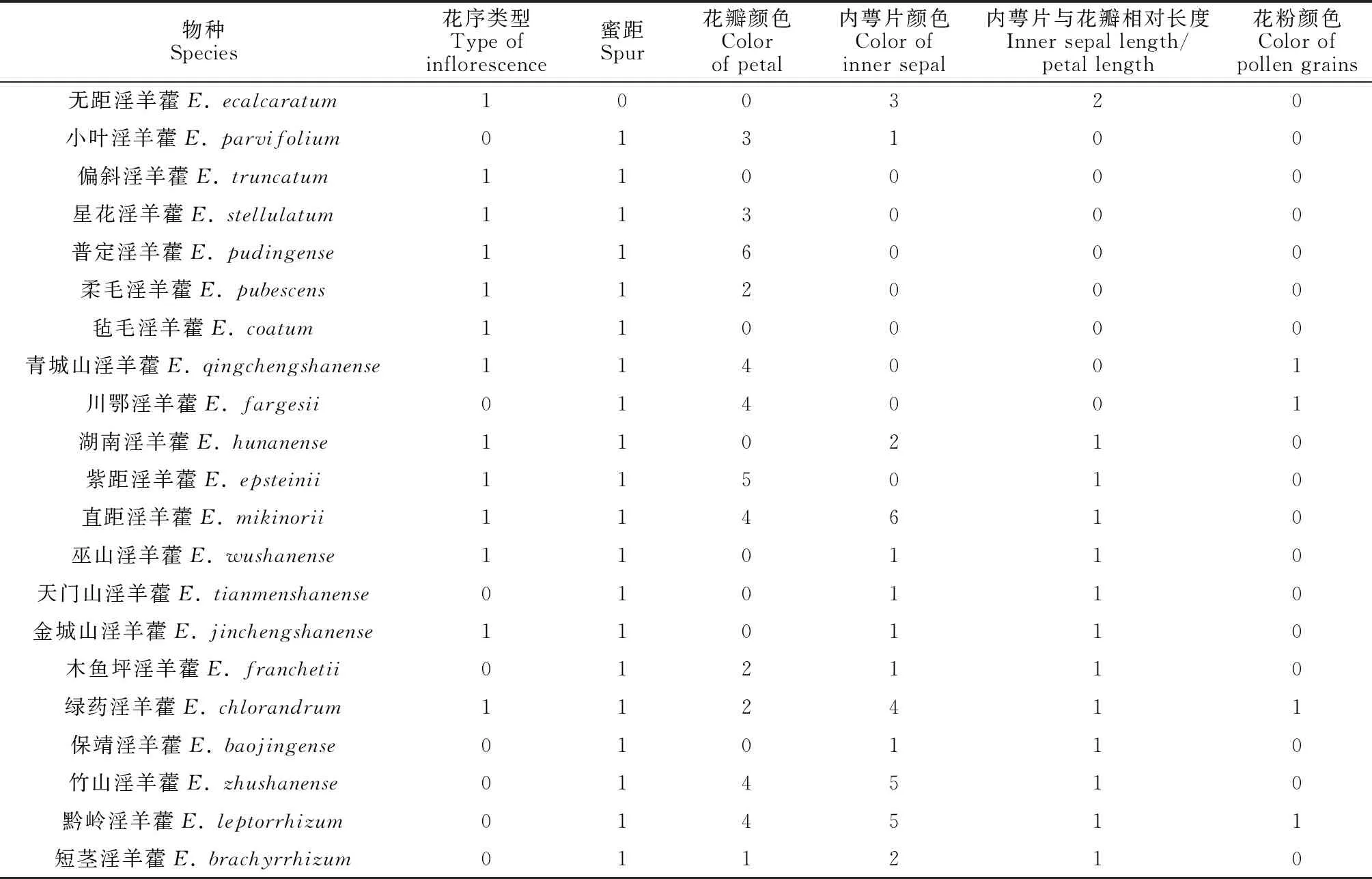
表3 21种淫羊藿质量性状编码
本研究中21种淫羊藿根据内萼片与花瓣的相对长度[33]可分成大花类型10种(花瓣远大于内轮萼片,花直径一般大于1 cm);小花类型11种(花瓣小于内轮萼片,花直径一般小于1 cm)。物种间各花部特征均存在显著差异(P<0.01,表4a,b)。其中,花直径、内萼片长宽、蜜距长、雌蕊长、柱头和蜜距口的高度差、胚珠数、花粉量、花粉直径等特征种间差异较大。
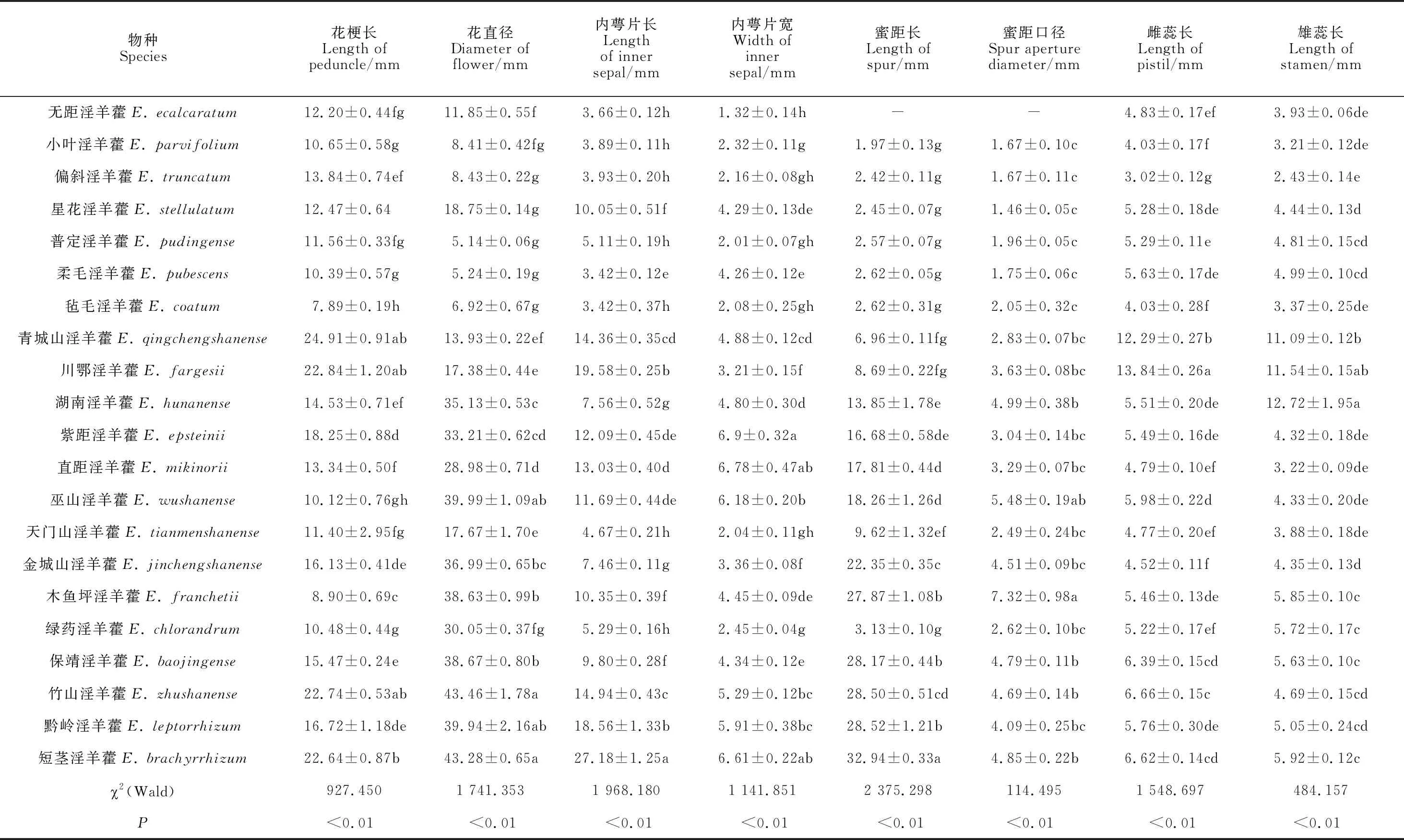
表4 21种淫羊藿花部特征测量参数(a)
2.2 主成分分析
在第1主成分中,花直径、内萼片长、花药长、胚珠数和花粉直径等性状负载值的绝对值都在0.80以上,信息负荷量最大,在本属分类中起主要作用(表5)。在第2主成分中,柱头和蜜距口的高度差和内萼片与花瓣相对长度等性状负载值的绝对值都在0.70以上,信息负荷量较大,对分类较为重要。在第3至第6主成分中,花瓣颜色、有无蜜距和花序类型等性状负载值的绝对值在0.59以上,在分类中也有一定的作用。主成分分析结果显示:前3个主成分累积贡献率为66.969%,直到第6个主成分累积贡献率达到85.631%,增值速度适中。
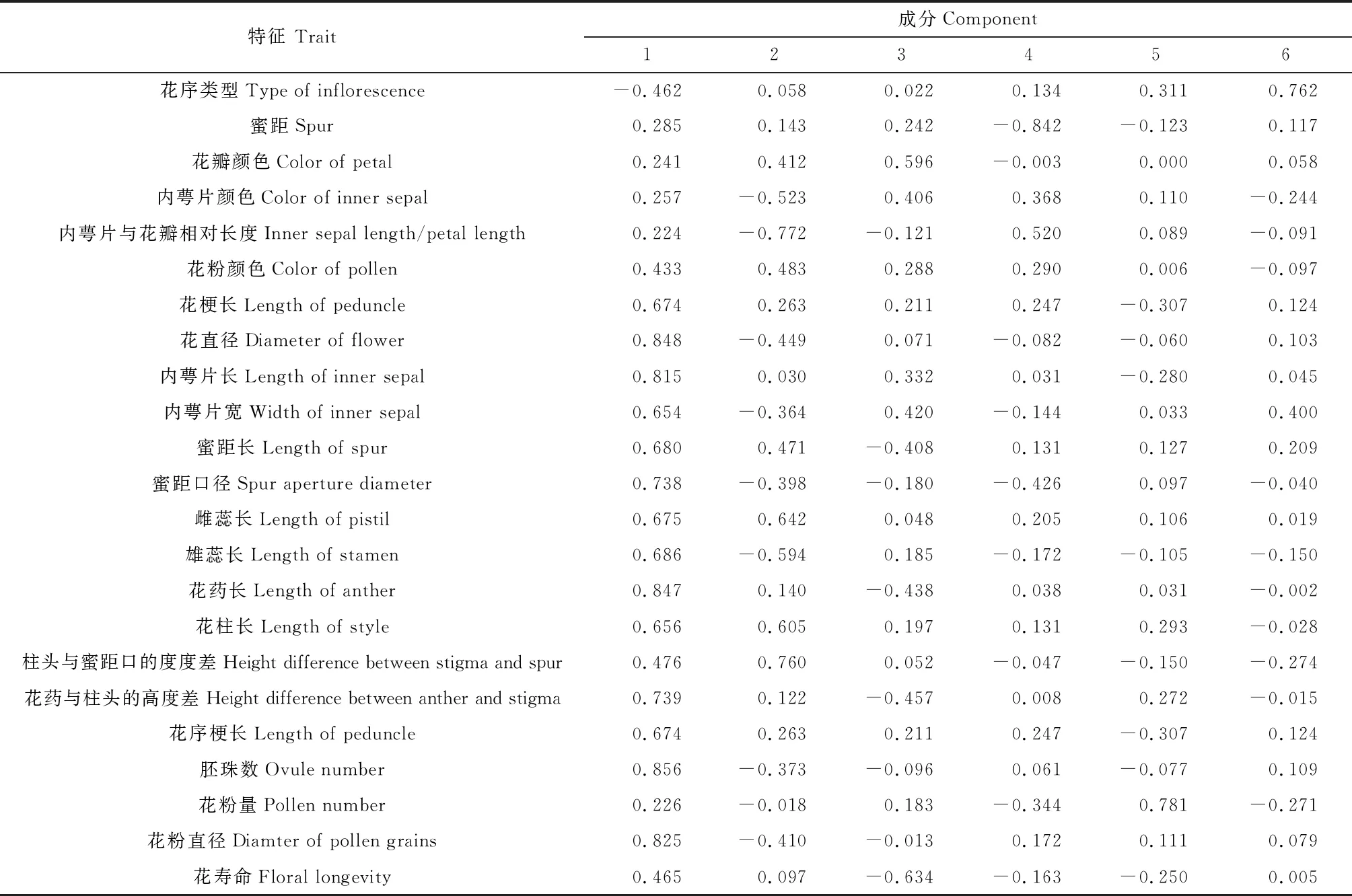
表5 淫羊藿属植物形态性状主成分负荷值
2.3 聚类分析
21种淫羊藿的系统聚类图表明在遗传距离为14的时候,21种淫羊藿根据有性繁殖特征分类,分为4支(图3)。分支 1星花淫羊藿、柔毛淫羊藿、普定淫羊藿、小叶淫羊藿、毡毛淫羊藿、偏斜淫羊藿聚为一支,主要特征为花小,花瓣短于内轮萼片,囊状距或兜状距,花直径一般小于1 cm。分支 2直距淫羊藿、绿药淫羊藿、木鱼坪淫羊藿、保靖淫羊藿、金城山淫羊藿、黔岭淫羊藿、竹山淫羊藿、天门山淫羊藿、紫距淫羊藿、巫山淫羊藿、湖南淫羊藿、短茎淫羊藿聚为一支,均为大花,其主要特征花瓣远长于内轮萼片,长距状,花直径一般大于1 cm。分支 3无距淫羊藿单独为一支,花小,花瓣扁平,花瓣没有特化形成蜜距,花直径(11.85±0.55) mm。分支4川鄂淫羊藿、青城山淫羊藿聚为一支,花丝伸长明显,可达10 mm,花蜜距呈钻状,内轮萼片长于花瓣,花直径14~18 mm。

淫羊藿属21种植物花部特征分种检索表
3 讨 论
物种间花部形态的变异是导致被子植物多样性的关键因素[34-35],因此花部特征常被用作被子植物分类和系统演化的重要依据[36],具有相近亲缘关系的属、种往往具有相似的花部形态特征[37]。在包含较多观赏和药用植物的云实亚科(Caesalpinioideae)中,花托、花萼、花冠、雄蕊、雌蕊等花部特征的性状是重要的分类依据[38];对蓼科头状蓼组(Sect.Cephalophilon)的19个分类群和与冰岛蓼属(Koenigia)相关的4个分类群的花形态进行比较,发现花特征的大小范围与分类群有关,花被片、雄蕊、花柱等特征是关键的分类依据[39]。同质园条件下21种淫羊藿的花寿命及花部特征均存在显著差异,尤其是花瓣、内萼片的形态及颜色。在前三主成分中,花直径、内萼片长、花药长、胚珠数和花粉直径等性状在本属分类中起主要作用。从聚类分析结果来看,在遗传距离为14时,21种淫羊藿属植物基于花部特征构建的聚类树形成稳定的4个分支,且具有较好的区分度。这表明淫羊藿属植物花部特征具有分类学意义,能通过花部特征将物种进行有效区分。
Stearn[18]将淫羊藿分为2亚属4组4系,其中中国组(Sect.Diphyllon)中种类最多,均来自中国,此组组下分为4个系:无距系(Ser.Campanulatae)、宝兴系(Ser.Davidianae)、长距系(Ser.Dolichocerae)和小花系(Ser.Brachycerae)。本实验的淫羊藿材料均来自中国组(Sect.Diphyllon)。基于同质园栽培的21种淫羊藿花部特征的聚类分析结果基本符合Stearn基于花形态学对中国组淫羊藿的分类,但不完全支持其对中国组4系的划分。本研究中分支 1星花淫羊藿、柔毛淫羊藿等花形态较小的物种聚为一支(属于Brachycerae系),而在Stearn的分类系统中属于Dolichocerae系的天门山淫羊藿在本试验与Brachycerae系物种聚为一支。天门山淫羊藿经调查发现是淫羊藿属植物大花和小花类群的一个中间过渡形态,形态变异巨大,可能是保靖淫羊藿的特殊生态类型[40];以往对淫羊藿属植物的分类通常基于腊叶标本进行描述,但该属植物花形态较为立体,制成标本后形态变形严重;此外标本数量有限,对物种性状的描述并不能全面概括。分支2中出现了Stearn系统中Davidianae系和Dolichocerae系的物种嵌套,该分枝的物种均属大花类群,具长距,且花瓣长于内萼片。但本研究结果与李超等[41]和张成[28]基于形态特征构建的淫羊藿属下类群的分类关系的结果一致,因此Stearn划分的Davidianae系和Dolichocerae系中部分物种分类位置有待商榷。分支4中川鄂淫羊藿、青城山淫羊藿聚为一支,没有与Brachycerae系的其他物种聚在一起。尽管上述2种淫羊藿具有内萼片长于花瓣的与小花类群相似的特征,但其花直径明显大于典型的小花类群物种介于大花类群和小花类群之间,且花丝明显伸长,长度可达10 mm。这与张成[28]基于形态特征构建的聚类关系结果一致,建议以后的分类学处理考虑将这一支成立为新系。
除了通过传统形态学分类外,基于分子手段的系统发育分析也致力于解决淫羊藿属的分类问题。基于ITS2序列对中国组15种淫羊藿的研究表明:该序列较为保守,趋异值低,因此ITS2不适合单独作为条形码用于该属系统发育分析[25]。Zhang等[26]通过对38种淫羊藿植物的ITS和叶绿体atpB-rbcL 2个序列的研究,认为虽然中国组类群(Sect.Diphyllon)与其他地理区域类群(Sect.Macroceras、Polyphyllon和Epimedium)有效区分,但在中国组的各系之间相互嵌套。类似的结果也出现于近年来的多项研究中,不论是基于核糖体DNA序列[24]、单核苷酸多态性(SNPs)数据[28],还是质体基因组数据[42],亦或是基于扩增片段长度多态性[43]等分子标记构建的系统发育树,均能强烈支持中国组(Sect.Diphyllon)和大花组(Sect.Macroceras)的姊妹关系,但不能完全支持Stearn对中国组4系的划分[18],特别是宝兴系(Ser.Davidianae)、长距系(Ser.Dolichocerae)的物种嵌套在各分支中,这与本研究基于同质园栽培的21种淫羊藿花部特征的分类结果一致。出现这一现象可能与淫羊藿属中国组分布区域的辐射型进化有关,其祖先可能经历了快速的物种形成[42]。
猜你喜欢
大自然探索(2023年9期)2023-10-09
江西中医药(2022年8期)2022-08-22
广西植物(2020年13期)2020-09-12
中国中医药现代远程教育(2018年22期)2018-02-09
湖南林业科技(2017年6期)2018-01-30
小学生导刊(2017年22期)2017-07-19
学苑创造·A版(2016年10期)2016-11-19
学生天地(2016年16期)2016-05-17
花卉(2015年11期)2015-09-14
陕西中医(2015年11期)2015-03-22
