
迷迭香酸对堆型艾美耳球虫攻毒肉鸡生长性能、免疫机能及炎症反应的影响
2023-10-16 12:49酉加民杨茜梓宫嘉泰何晨鹏蒋亦轩贺建华伍树松
动物营养学报 2023年9期
酉加民 杨茜梓 宫嘉泰 何晨鹏 蒋亦轩 贺建华 伍树松*
(1.湖南农业大学动物科学技术学院,长沙 410128;2.湖南德诺健康产业集团有限公司,长沙 410100)
家禽生产是畜牧业增长最快的产业之一,为全球提供大量蛋白质。球虫病是家禽生产中最常见的寄生虫病,每年给家禽生产造成超过8亿美元的损失[1]。艾美耳虫属(Eimeria)是球虫病的主要病原体,可引起肠道炎症,其代谢产生的毒素还可引起全身性中毒症状与炎症[2]。在过去的几十年里,通常采用在饲粮中添加抗球虫药物(如氨溴铵、脱奎宁、双青尿尔和盐霉素)的方式来预防和控制球虫,然而长期使用这些药物往往伴随着耐药性的产生[3-4]。在全面“饲料禁抗”时代,开发可防治家禽球虫病的新型绿色饲料添加剂显得尤为迫切。
迷迭香酸(RA)是迷迭香、牛至、百里香等中药材的主要活性成分之一,具有抑菌、抗炎、抗氧化等生物活性[5]。有研究报道,RA可以抑制乙醇诱导的肿瘤坏死因子-α(TNF-α)和白细胞介素-6(IL-6)的mRNA表达,阻止乙醇诱导的山梨醇的增加,进而缓解酒精性肝炎[6]。在大肠杆菌K88(ETEC K88)诱导断奶仔猪模型中,饲粮添加RA可以改善ETEC K88攻毒断奶仔猪结肠炎症反应,缓解断奶仔猪腹泻[7]。在硫酸葡聚糖钠(DDS)诱导的结肠炎模型中,RA通过降低IL-6、白细胞介素-1β(IL-1β)、白细胞介素-22(IL-22)、环氧化酶-2(COX-2)和诱导型一氧化氮合成酶(iNOS)的蛋白表达水平,抑制核因子-κB(NF-κB)的激活,发挥对结肠炎的改善作用[8]。基于生物信息学的分析表明,RA可调控炎症及氧化应激相关基因表达[9]。此外,研究显示,饲粮中添加富含RA的草本植物(迷迭香、百里香、牛至和大蒜)可改善家禽免疫机能和抗氧化能力,降低家禽的发病率和死亡率[10-11]。本研究以堆型艾美耳球虫(Eimeriaacervulina,E.acervulina)攻毒黄羽肉鸡作为模型,探究RA对堆型艾美耳球虫攻毒肉鸡生长性能、免疫机能以及炎症反应的影响,以期为RA在家禽生产中进一步的研究与应用提供参考。
1 材料与方法
1.1 试验动物与分组
选择健康的1日龄黄羽肉鸡360羽,随机分为6个组,每组6个重复,每个重复10羽。对照组和攻毒对照组饲喂基础饲粮,阳性对照组在基础饲粮中添加20 mg/kg地克珠利,3个试验组在基础饲粮中分别添加40、80、160 mg/kg RA(由湖南某公司提供)。攻毒对照组、阳性对照组和试验组雏鸡在14日龄时灌喂孢子化的堆型艾美耳球虫卵悬液1 mL(孢子数量不低于4×104个/mL),对照组灌喂相同体积的生理盐水。试验期为8周(56 d)。
1.2 饲养管理
所有试验肉鸡采用笼养,每笼10羽,通过水壶式饮水器(1~14日龄)或乳头式饮水器(14~56日龄)全天无限制供应,每日07:30和16:30各投喂1次饲粮,自由采食,每2~3 d采用传粪带清粪的方式清粪1次,随着鸡日龄增加清粪频率也随之增加。试验前1周鸡舍采取锅炉增温,维持温度在33 ℃,并随着肉鸡日龄增加逐步降低鸡舍内温度至23 ℃。试验期全程进行常规免疫和消毒程序,保证鸡舍内环境卫生质量。
1.3 试验饲粮
试验用基础饲粮为玉米-豆粕型,根据《鸡饲养标准》(NY/T 33—2004)中肉鸡营养需要,分为0~4周龄与5~8周龄2个阶段配制,基础饲粮组成及营养水平见表1。
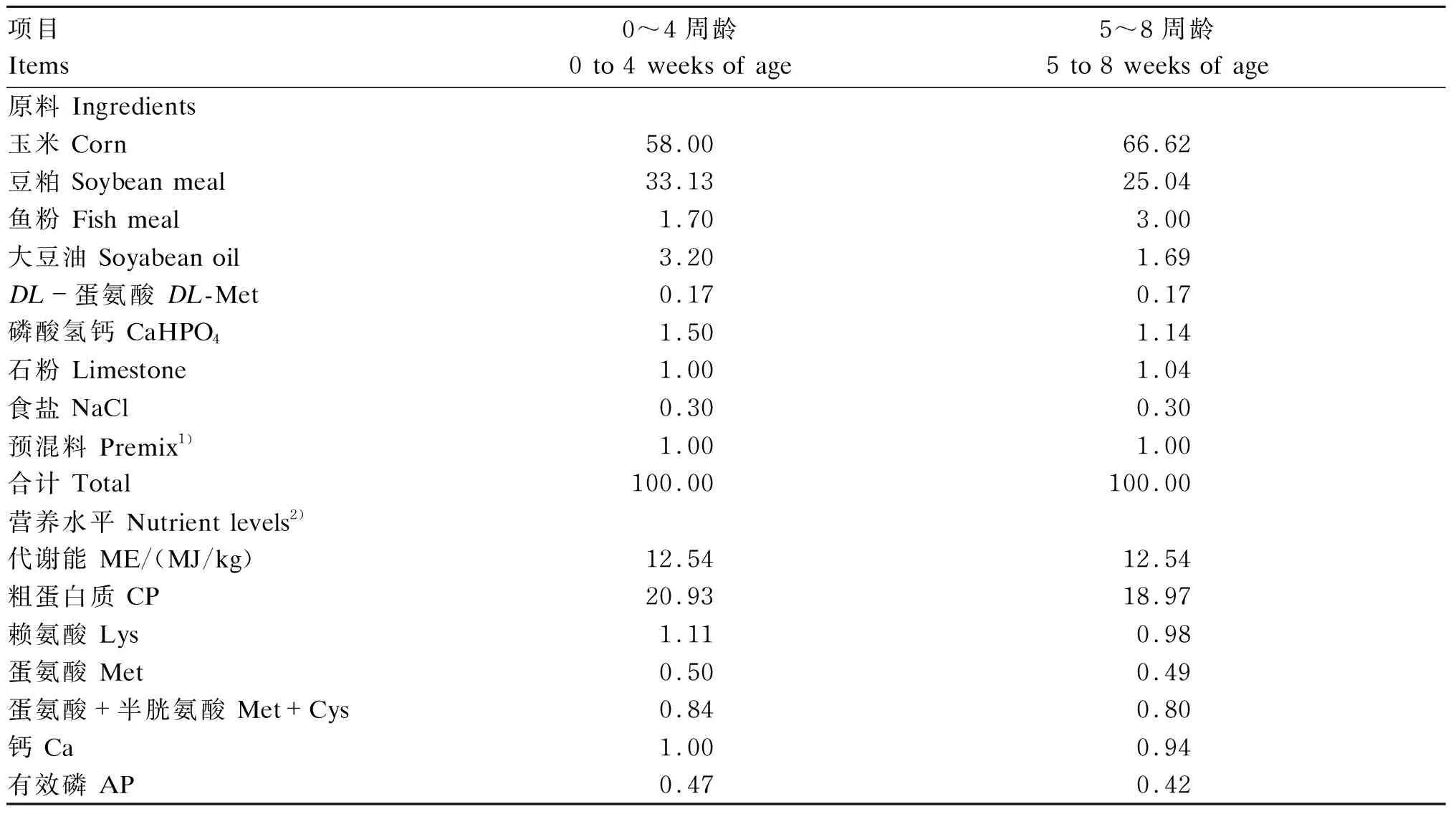
表1 基础饲粮组成及营养水平(风干基础)
1.4 样品采集与指标测定
1.4.1 生长性能与死亡率
以重复为单位记录1、14、21和56日龄鸡体重以及每日的采食量和鸡死亡数,并计算鸡1~14日龄、15~21日龄和22~56日龄3个阶段的平均日采食量、平均日增重、料重比和死亡率。如存在死亡淘汰鸡只,平均日采食量和料重比需要及时进行校对。
1.4.2 球虫计数
在堆型艾美耳球虫攻毒后第4、7、10天采用五点法收集每个重复肉鸡的粪便3~4 g,装于5 mL离心管中,在-20 ℃冰箱保存。用麦克马斯特法进行球虫卵囊计数。取2 g新鲜鸡粪便置于干净的100 mL烧杯内,先加入8 mL饱和食盐溶液,用玻璃棒捣碎混匀,再加入50 mL饱和食盐溶液,混匀后立即用60目筛网过滤,然后立即吸取滤液充满2个计数室,在显微镜载物台上静置3~5 min,在10倍物镜下镜检计数,每个计数室内有100个方格,其体积为1 cm×1 cm×0.15 cm=0.15 mL,分别查完2个计数室100个方格内的卵囊数量,最后按照如下公式计算每克粪便中卵囊数量(OPG):
OPG=[(n1+n2)/(2×0.15)]×60/2。
式中:n1、n2分别为上、下计数室卵囊数量。
1.4.3 血清生化指标
在堆型艾美耳球虫攻毒后第1周(21日龄)每个重复随机抽取1只肉鸡,翅静脉采血5 mL于10 mL离心管中,静置10 min后3 000 r/min离心,取上层血清分装于数个1.5 mL EP管中,-20 ℃保存。采用科华全自动生化仪(卓越450,上海)按说明方法检测血清生化指标。
1.4.4 免疫器官指数
在21、28、56日龄每个重复随机抽取1只肉鸡,称重后处死、解剖,取脾脏、肝脏、法氏囊、胸腺称重,并计算肝脏、脾脏、法氏囊、胸腺指数。
肝脏指数(%)=[肝脏重(g)/活体重(g)]×100;
脾脏指数(%)=[脾脏重(g)/活体重(g)]×100;
法氏囊指数(%)=[法氏囊重(g)/
活体重(g)]×100;
胸腺指数(%)=[胸腺重(g)/活体重(g)]×100。
1.4.5 血清、肝脏炎性因子与免疫学指标
采用上海优选生物公司生产的酶联免疫吸附测定(ELISA)试剂盒,按照试剂盒说明检测血清和肝脏中TNF-α、IL-1β、IL-6、补体3(C3)、补体4(C4)、免疫球蛋白A(IgA)、免疫球蛋白G(IgG)、免疫球蛋白M(IgM)含量。
1.5 数据处理与分析
试验数据初步整理剔出离群值后,采用SPSS 23.0 软件进行单因素方差分析,并采用Duncan氏法进行组间多重比较,P<0.05为差异显著,P<0.01为差异极显著。试验数据采用平均值±标准差表示。
2 结 果
2.1 RA对堆型艾美耳球虫攻毒肉鸡粪便卵囊数量的影响
由图1可知,堆型艾美耳球虫攻毒后第4、7、10天,攻毒对照组肉鸡OPG均较对照组显著上升(P<0.05);而与攻毒对照组相比,添加40(攻毒后第10天除外)、80、160 mg/kg RA或20 mg/kg地克珠利均显著降低了肉鸡的OPG(P<0.05)。
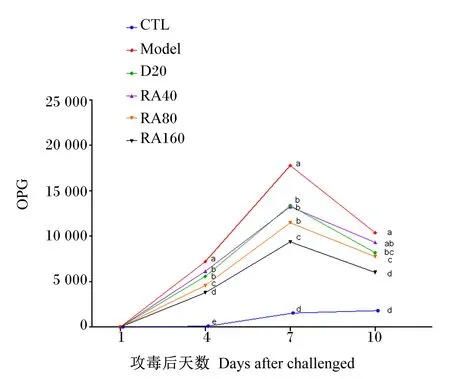
CTL:对照组;Model:攻毒对照组;D20:阳性对照组;RA40:40 mg/kg迷迭香酸组;RA80:80 mg/kg迷迭香酸组;RA160:160 mg/kg迷迭香酸组;OPG:每克粪便中卵囊数量。同一时间数据点标注不同字母表示差异显著(P<0.05)。
2.2 RA对堆型艾美耳球虫攻毒肉鸡生长性能与死亡率的影响
由表2可知,各组间肉鸡1~14日龄平均日增重、平均日采食量和料重比均无显著差异(P>0.05)。与对照组相比,堆型艾美耳球虫攻毒显著降低了肉鸡15~21日龄平均日增重(P<0.05);与攻毒对照组相比,添加160 mg/kg RA显著提高了堆型艾美耳球虫攻毒肉鸡15~21日龄平均日增重(P<0.05);各组间肉鸡15~21日龄平均日采食量和料重比均无显著差异(P>0.05)。各组间肉鸡22~56日龄平均日增重、平均日采食量和料重比均无显著差异(P>0.05)。
由表2可知,与对照组相比,攻毒对照组15~21日龄肉鸡死亡率上升了3.52%;与攻毒对照组相比,阳性对照组、40 mg/kg RA组、80 mg/kg RA组和160 mg/kg RA组肉鸡死亡率分别降低了3.52%、3.52%、5.18%、3.52%。与对照组相比,攻毒对照组22~56日龄肉鸡死亡率上升了6.58%;与攻毒对照组相比,阳性对照组、40 mg/kg RA组、80 mg/kg RA组和160 mg/kg RA组肉鸡死亡率分别降低了4.84%、5.06%、7.15%、10.85%。
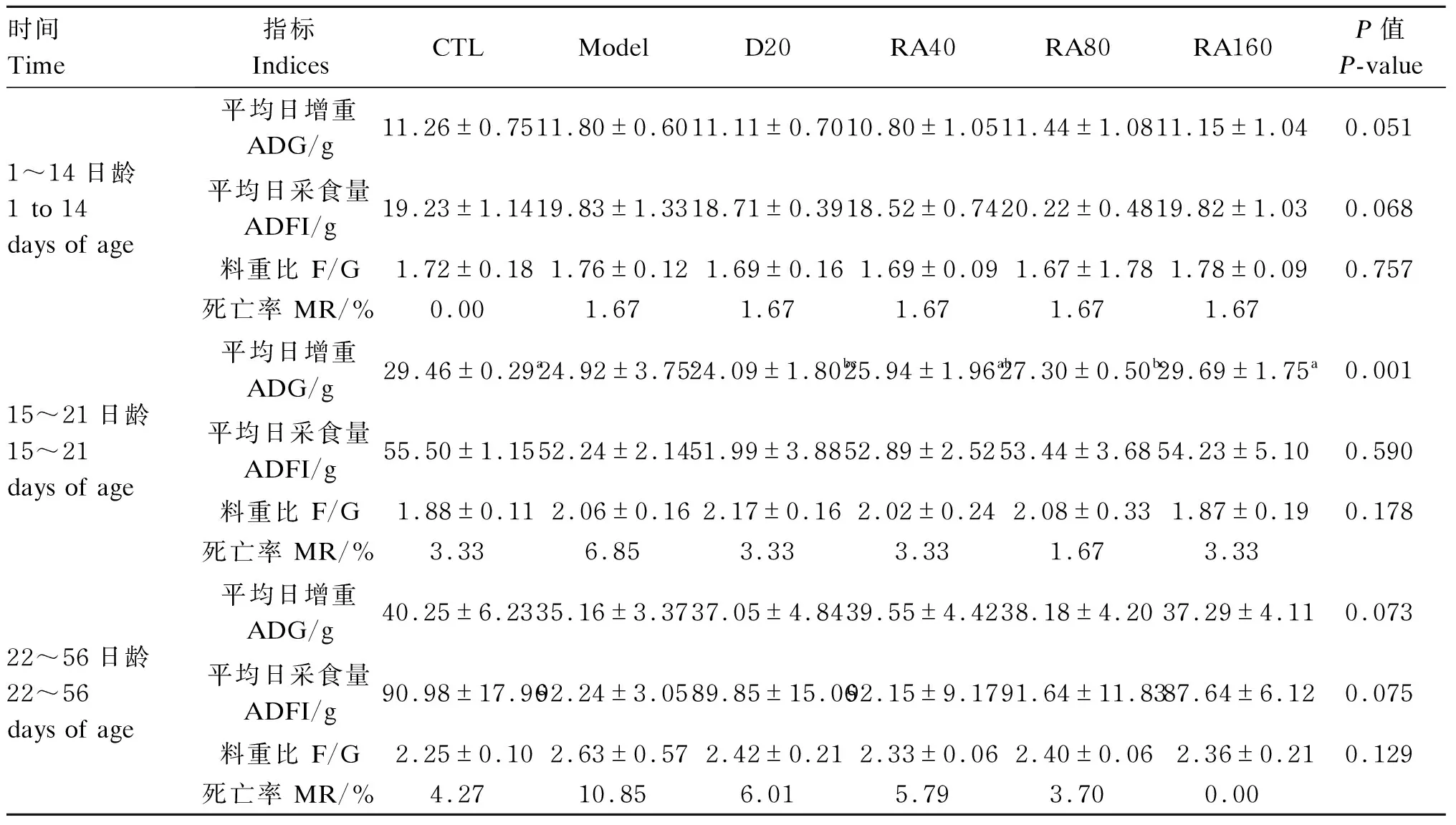
表2 RA对堆型艾美耳球虫攻毒肉鸡生长性能和死亡率的影响
2.3 RA对堆型艾美耳球虫攻毒肉鸡免疫器官指数的影响
由表3可知,与对照组相比,堆型艾美耳球虫攻毒显著提高了肉鸡21日龄肝脏指数和脾脏指数(P<0.05),但对法氏囊指数与胸腺指数无显著影响(P>0.05);与攻毒对照组相比,添加160 mg/kg RA显著降低了堆型艾美耳球虫攻毒肉鸡21日龄肝脏指数(P<0.05)。
由表4可知,与对照组相比,堆型艾美耳球虫攻毒对肉鸡28日龄法氏囊指数、肝脏指数、脾脏指数、胸腺指数均无显著影响(P>0.05);与攻毒对照组相比,添加40、80、160 mg/kg RA或20 mg/kg地克珠利对堆型艾美耳球虫攻毒肉鸡28日龄法氏囊指数、肝脏指数、脾脏指数、胸腺指数均无显著影响(P>0.05)。
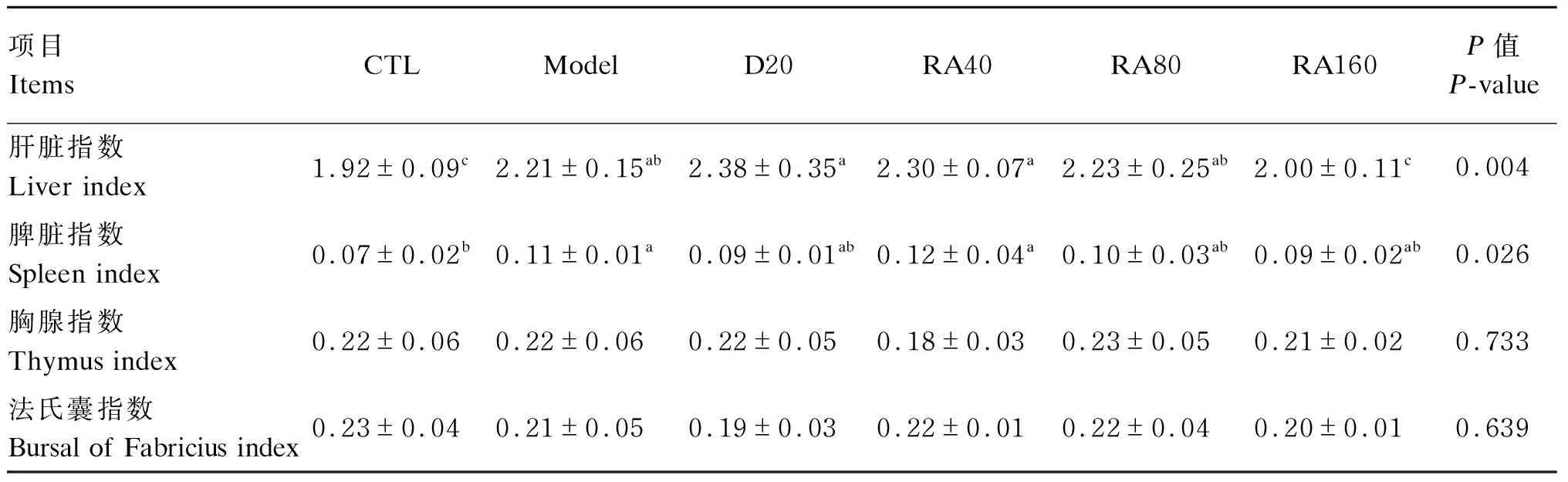
表3 RA对堆型艾美耳球虫攻毒肉鸡21日龄免疫器官指数的影响
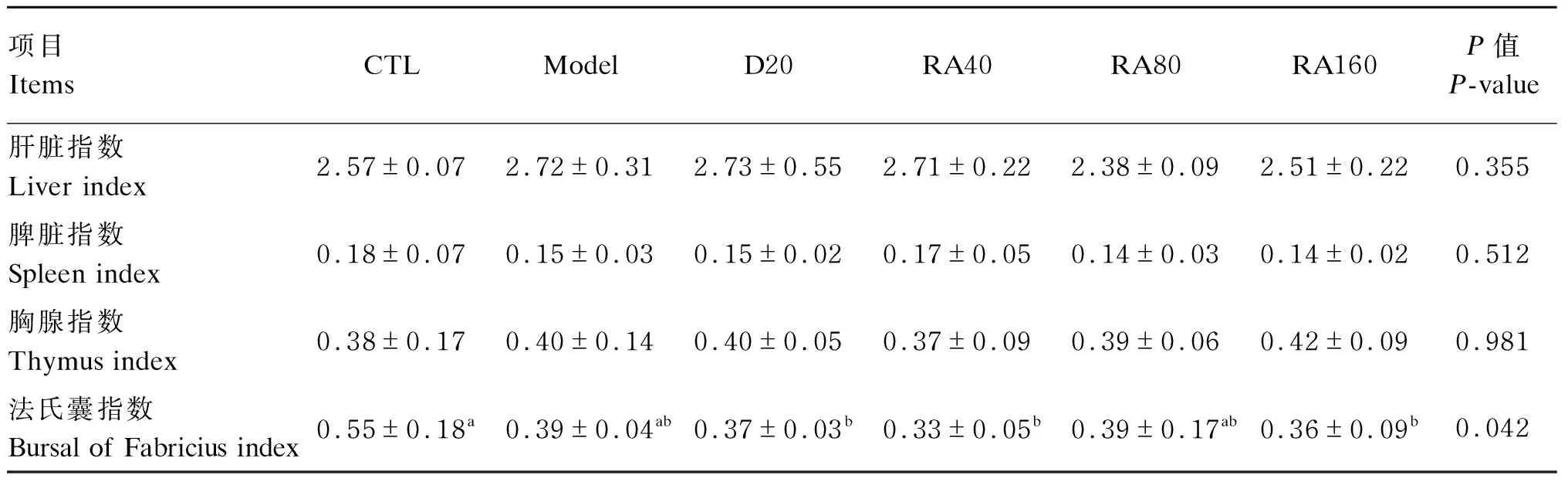
表4 RA对堆型艾美耳球虫攻毒肉鸡28日龄免疫器官指数的影响
由表5可知,与对照组相比,堆型艾美耳球虫攻毒对56日龄肉鸡肝脏指数、脾脏指数、胸腺指数法氏囊指数无显著影响(P>0.05);与攻毒对照组相比,添加160 mg/kg RA显著增加了56日龄肉鸡法氏囊指数(P>0.05)。
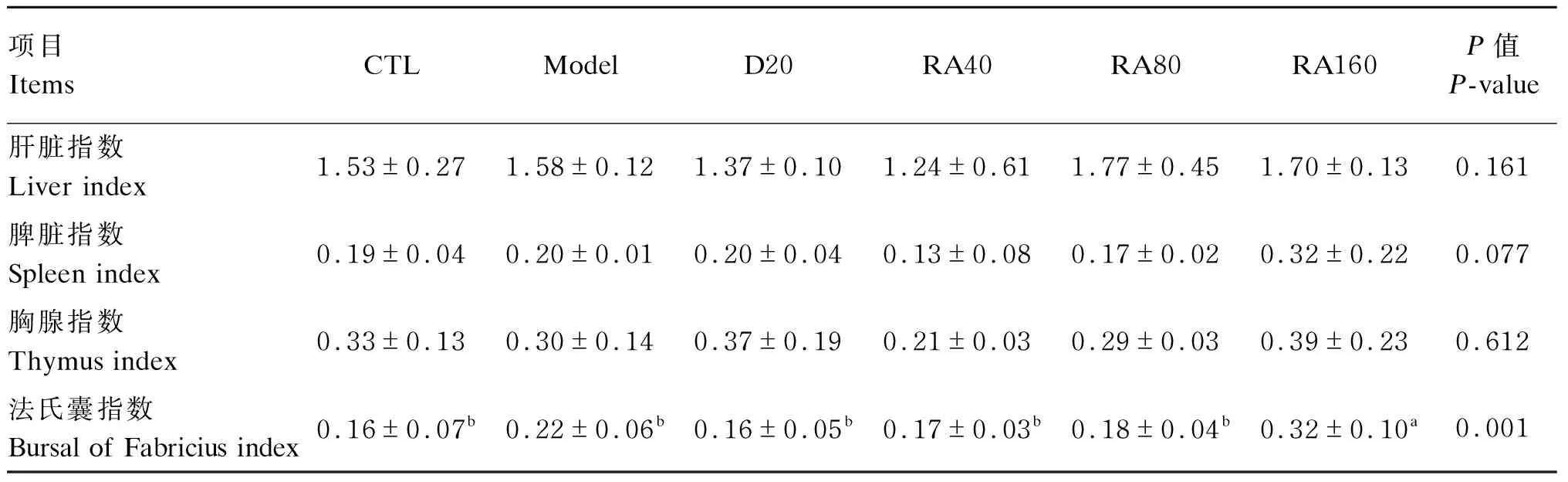
表5 RA对堆型艾美耳球虫攻毒肉鸡56日龄免疫器官指数的影响
2.4 RA对堆型艾美耳球虫攻毒肉鸡血清与肝脏免疫学指标的影响
由表6可知,与对照组相比,堆型艾美耳球虫攻毒显著提高了肉鸡21日龄血清中C3、C4含量(P<0.05),显著降低了21日龄肉鸡血清IgM含量(P<0.05),但对血清中IgA、IgG含量无显著影响(P>0.05);与攻毒对照组相比,添加40、80、160 mg/kg RA或20 mg/kg地克珠利均显著降低了堆型艾美耳球虫攻毒肉鸡21日龄血清中C3、C4含量(P<0.05),添加80、160 mg/kg RA显著提高了血清中IgA含量(P<0.05),添加40、160 mg/kg RA显著降低了血清中IgG含量(P<0.05),添加160 mg/kg RA或20 mg/kg地克珠利显著提高了血清中IgM含量(P<0.05)。
与对照组相比,堆型艾美耳球虫攻毒显著提高了肉鸡21日龄肝脏中C3、C4含量(P<0.05),显著降低了肝脏中IgA含量(P<0.05),但对肝脏中IgG和IgM含量无显著影响(P>0.05);与攻毒对照组相比,添加160 mg/kg RA或20 mg/kg地克珠利显著降低了肝脏中C3含量(P<0.05),添加40、80、160 mg/kg RA或20 mg/kg地克珠利均显著降低了肝脏中C4含量(P<0.05),添加80 mg/kg RA显著提高了肝脏中IgA含量(P<0.05),添加160 mg/kg RA显著提高了肝脏中IgG含量(P<0.05),添加40、80、160 mg/kg RA或20 mg/kg地克珠利均显著提高了肝脏中IgM含量(P<0.05)。
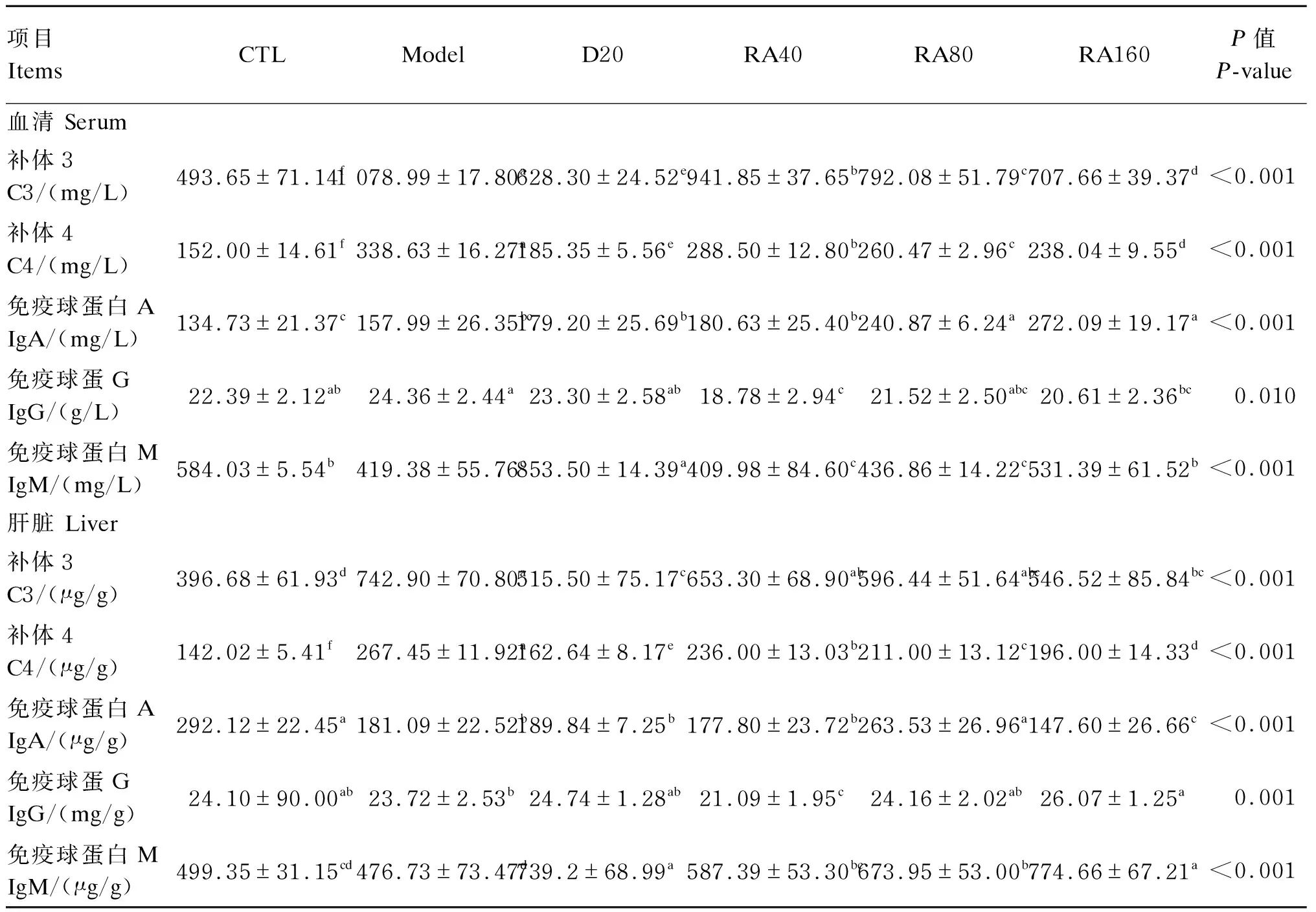
表6 RA对堆型艾美耳球虫攻毒肉鸡21日龄血清与肝脏免疫学指标的影响
2.5 RA对堆型艾美耳球虫攻毒肉鸡血清与肝脏炎性因子含量的影响
由表7可知,与对照组相比,堆型艾美耳球虫攻毒显著提高了21日龄肉鸡血清与肝脏IL-1β、IL-6、TNF-α含量(P<0.05);与攻毒对照组相比,添加20 mg/kg地克珠利或40、80、160 mg/kg RA均显著降低了21日龄肉鸡血清与肝脏IL-1β、IL-6、TNF-α含量(P<0.05)。
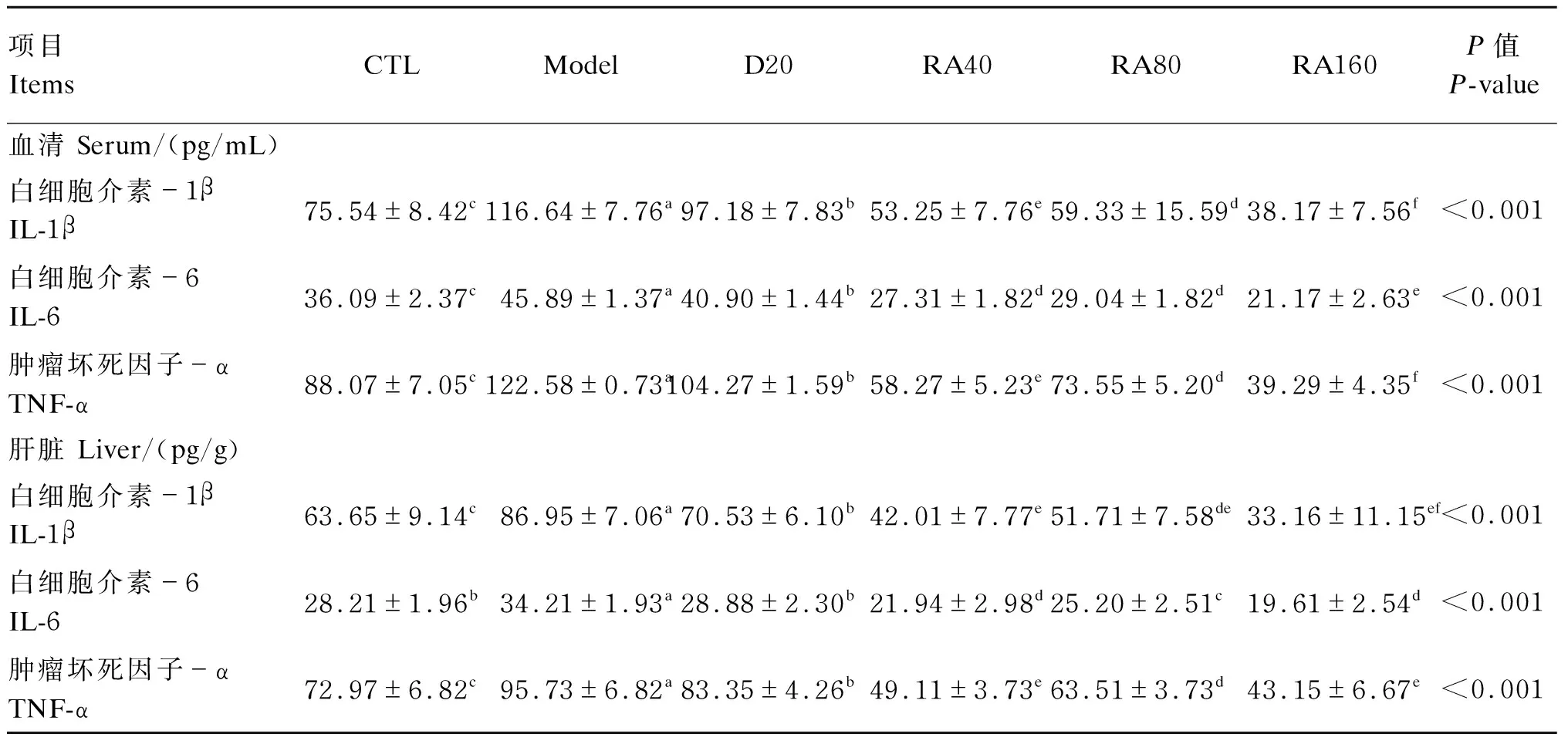
表7 RA对堆型艾美耳球虫攻毒肉鸡21日龄血清与肝脏炎性因子含量的影响
2.6 RA对堆型艾美耳球虫攻毒肉鸡血清生化指标的影响
由表8可知,与对照组相比,堆型艾美耳球虫攻毒显著提高了肉鸡21日龄血清中谷丙转氨酶(ALT)活性(P<0.05),对其他血清生化指标无显著影响(P>0.05);与攻毒对照组相比,添加20 mg/kg地克珠利或40、80、160 mg/kg RA显著降低了堆型艾美耳球虫攻毒肉鸡21日龄血清中ALT活性(P<0.05),添加40、80 mg/kg RA显著降低了血清中尿素氮(UN)含量(P<0.05),添加40 mg/kg RA显著降低了血清中总胆固醇(TC)含量(P<0.05)。
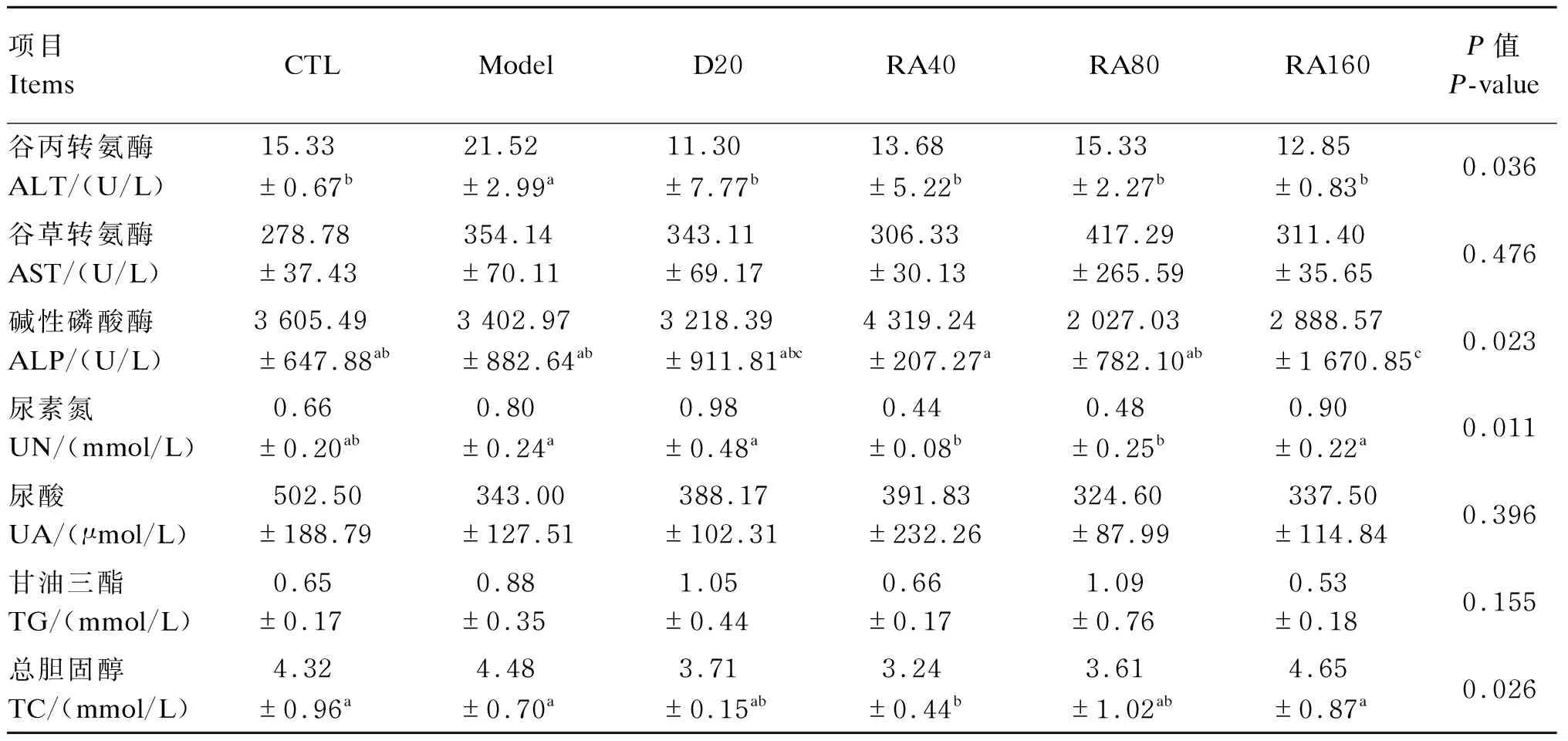
表8 RA对堆型艾美耳球虫攻毒肉鸡21日龄血清生化指标的影响
3 讨 论
3.1 堆型艾美耳球虫攻毒对肉鸡生长性能与OPG的影响
球虫病的发生与发展十分复杂,通常是由多种不同种类艾美尔球虫共同感染的结果。不同的艾美尔球虫致病性与典型症状不尽相同[12],例如,毒害艾美尔球虫与柔嫩艾美尔球虫毒性最大,导致肠绒毛完全破坏,产生出血和死亡;布氏艾美耳球虫和巨型艾美尔球虫导致肠壁肿胀、瘀点和上皮细胞松动;而堆型艾美耳球虫和变位艾美尔球虫感染的典型症状是导致水分流失和营养吸收减少,进而导致日增重降低、料重比增加[13-15]。本试验中,堆型艾美耳球虫攻毒使肉鸡平均日增重显著降低,死亡率增加,与堆型艾美耳球虫感染的典型症状一致,且从堆型艾美耳球虫攻毒肉鸡的粪便检测到大量堆型艾美耳球虫虫卵,表明本研究成功建立了肉鸡的堆型艾美耳球虫攻毒模型。
3.2 RA对堆型艾美耳球虫攻毒肉鸡生长性能的影响
Bozkurt等[16]研究表明,在饲粮中添加12或24 mg/kg牛至精油可以提高堆型艾美耳球虫攻毒肉鸡的体增重。Alp等[17]研究表明,在饲粮中添加300或600 mg/kg牛至精油可以缓解柔嫩艾美尔球虫与堆型艾美耳球虫攻毒引起的平均日增重下降,并降低攻毒肉鸡的料重比。Fotea等[18]研究表明,当迷迭香精油添加量为0.5%时,可提高育肥期肉鸡的生长性能和饲料利用率。本试验中,在饲粮中添加160 mg/kg RA显著提高了堆型艾美耳球虫攻毒肉鸡15~21日龄平均日增重,这与前人的研究结果一致,说明饲粮中添加RA对堆型艾美耳球虫攻毒导致的肉鸡生长性能下降有一定的缓解效果。
3.3 RA对堆型艾美耳球虫攻毒肉鸡OPG的影响
在生产中,通常采用OPG衡量球虫感染严重程度[19-21]。在进行堆型艾美耳球虫攻毒的第4天,各攻毒组的粪便中均检测到球虫的排出,在第7天OPG达到高峰,随后OPG缓慢下降。这说明堆型艾美耳球虫进入肉鸡消化道7 d内快速地繁殖导致数量上巨大增长,使得通过粪便排出的球虫卵囊数量增加。Arczewska-Wosek等[21]研究表明,在饲粮中添加1 000 mg/kg植物精油混合物(迷迭香、百里香、紫雏菊、丹参、牛至)可有效减少巨型艾美尔球虫、柔嫩艾美尔球虫攻毒肉鸡OPG。本试验中,饲粮中添加40、80、160 mg/kg RA或20 mg/kg地克珠利均减少了攻毒4、7、10 d肉鸡粪便OPG。体外研究表明,4 mg/mL富含RA的植物精油可以破坏85%~90%的艾美尔球虫卵囊[22]。这可能是饲粮中添加RA显著降低肉鸡OPG的原因之一。
3.4 RA对堆型艾美耳球虫攻毒肉鸡免疫器官指数的影响
免疫器官指数常用来评价鸡的免疫状态[23]。脾脏是禽类最大的外周免疫器官,参与机体的细胞免疫与体液免疫。法氏囊是禽类特有的中枢免疫器官,主导机体的体液免疫。胸腺为T淋巴细胞分化、发育和成熟提供场所。肝脏是体内最大的网状内皮细胞吞噬系统,对抗原的过滤、清除、吞噬和调节和免疫反应的作用。在早期研究中,去除法氏囊的鸡对继发性艾美球虫感染产生保护性免疫的能力没有显著影响[24],而切除胸腺的大鼠对原发性和继发性球虫感染的抵抗力均显著降低[25]。有研究报道,球虫感染会损害肉鸡肝脏组织结构[26]。宋鸿雁等[27]的研究表明,斯氏艾美耳球虫感染导致新西兰兔肝脏严重受损,且血清中ALT、谷草转氨酶(AST)、乳酸脱氢酶(LDH)、ALP活性与总胆红素(T-BIL)含量极显著增高。本试验中,堆型艾美耳球虫攻毒导致肉鸡脾脏指数、肝脏指数显著增加,但对法氏囊指数、胸腺指数无显著影响,这与前人的研究结果基本一致。进一步对血清生化指标进行分析发现,血清中ALT活性在堆型艾美耳球虫攻毒后显著增加,而血清中ALT活性升高是肝脏炎症或肝细胞受损的表现[28]。前人研究发现,在对乙酰氨基酚诱导的小鼠急性肝损伤模型中,小鼠血清ALT活性显著升高,RA可以降低血清ALT活性,缓解对乙酰氨基酚诱导的小鼠急性肝损伤[29]。本试验中,饲粮中添加160 mg/kg RA显著降低了堆型艾美耳球虫攻毒肉鸡21日龄的肝脏指数与血清中ALT活性,说明RA对肉鸡肝脏损伤有一定的修复作用。
3.5 RA对堆型艾美耳球虫攻毒肉鸡血液与肝脏免疫学指标的影响
前人研究表明堆型艾美耳球虫攻毒可使鸡血清补体含量升高[30]。本试验结果表明,堆型艾美耳球虫攻毒显著提高了肉鸡血清、肝脏中C3、C4含量。体外研究表明,RA可以特异性连接到C3b的a9-链上,抑制补体的激活[31]。本试验中,饲粮添加RA降低了堆型艾美耳球虫攻毒肉鸡肝脏中C3、C4含量。研究显示,鸡感染艾美尔球虫后,会产生大量的特异性IgA、IgG、IgM,这些抗体主要分布在血清、胆汁和肠道分泌物中;雏鸡初次感染毒害艾美耳球虫后,血清中IgA、IgG、IgM含量显著升高[32]。本试验中,堆型艾美耳球虫攻毒提高了肉鸡血清中IgA、IgM含量,降低了肝脏中IgA、IgM含量。台瑞青[33]的研究表明,在饲粮中添加200 mg/kg RA可以提高育肥猪血清中IgM含量。Sanbongi等[34]研究发现,RA可显著抑制过敏原特异性IgG。而Farouk等[35]给感染大肠杆菌的肉鸡饲喂添加了迷迭香的饲粮后,血清中IgG含量增加。Al Sheyab等[36]研究表明,迷迭香提取物增加了小鼠血清中IgM和IgG含量。本试验中,RA提高了堆型艾美耳球虫攻毒肉鸡血清中IgM含量,而使IgG含量下降,这与Sanbongi等[34]的研究结果一致。
3.6 RA对堆型艾美耳球虫攻毒肉鸡血清与肝脏炎性细胞因子含量的影响
TNF-α作为炎症中主要的细胞因子,是在细菌、病毒、免疫复合物、补体因子、细胞因子等刺激下产生,能够激活适应性免疫系统、其他促炎细胞因子(IL-1β和IL-6)和生长因子的转录,以及活化单核细胞和巨噬细胞[37]。鸡外周血白细胞来源的巨噬细胞在受到柔嫩艾美耳球虫孢子和裂殖子刺激时,TNF-α的产生呈现剂量与时间依赖性[38-39]。IL-1β是参与初始炎症反应的重要细胞因子,它诱导趋化因子的产生,从而促进炎症细胞在炎症部位的募集[40],且在感染球虫的肉鸡体内检测到它的表达上调[41]。IL-6负责B细胞最终成熟并产生抗体,并在感染堆型艾美耳球虫、巨大艾美尔和柔嫩艾美尔球虫的鸡的上皮内淋巴细胞中表达增强,这表明IL-6在适应性免疫中发挥作用[42]。这些研究表明炎性细胞因子在球虫病的发病机制中扮演重要角色。本试验中,进行堆型艾美耳球虫攻毒后,肉鸡血清、肝脏中TNF-α、IL-1β和IL-6含量显著上升,说明堆型艾美耳球虫球虫攻毒激活了肉鸡适应性免疫系统并引起肝脏炎症。研究表明,RA显著降低大鼠肝脏中TNF-α、IL-1β的mRNA表达水平,缓解缺血再灌注引起的肝脏急性炎症[43]。最近的研究发现,RA与百里香酚复配能够降低脂多糖攻毒小鼠空肠黏膜中IL-6含量[44]。在本试验中,在饲粮中添加RA显著降低了堆型艾美耳球虫攻毒肉鸡血清和肝脏中TNF-α、IL-1β、IL-6含量,说明RA缓解了堆型艾美耳球虫攻毒引起的炎症与肝脏损伤。信号转导和转录激活因子3(STAT3)和NF-κB协同调控多个靶基因,并协同调控相关细胞因子和趋化因子,STAT3直接与p65和p50相互作用,促进NF-κB募集到STAT3启动子,反之亦然[45]。研究表明,RA在DDS诱导的慢性结肠炎小鼠模型中可以抑制NF-κB和STAT3核转位[8]。因此,RA可能是通过抑制NF-κB和STAT3核转位,下调下游的炎性细胞因子(TNF-α、IL-1β、IL-6),进而缓解由堆型艾美耳球虫感染引起的肝脏炎症。值得注意的是,在一项早期的研究中发现,抑制肿瘤坏死因子使得苏赛斯鸡在感染球虫时体重下降减缓[42],因此,球虫攻毒后,机体发生炎症反应,导致基础代谢增快和能量消化增加,这可能是攻毒对照组15~21日龄肉鸡平均日增重显著降低的原因之一。
4 结 论
综上所述,在本试验条件下,饲粮添加160 mg/kg RA可提高堆型艾美耳球虫攻毒1周后肉鸡的平均日增重和血清中免疫球蛋白(IgA、IgM)含量,降低肝脏指数和血清、肝脏中补体(C3、C4)和炎症细胞因子(IL-1β、IL-6、TNF-α)含量,其效果优于20 mg/kg地克珠利或40~80 mg/kg RA。以上结果表明,饲粮添加160 mg/kg RA可有效缓解堆型艾美耳球虫引起的肉鸡生长缓慢、免疫失衡与肝脏炎症。
猜你喜欢
广东农业科学(2022年5期)2022-07-21
辐射防护(2021年6期)2021-12-06
中国动物保健(2021年8期)2021-11-24
河南畜牧兽医(2020年23期)2020-01-10
畜牧与饲料科学(2016年9期)2016-11-01
湖南畜牧兽医(2016年3期)2016-06-05
电站辅机(2016年1期)2016-03-08
核科学与工程(2015年3期)2015-09-26
中国预防兽医学报(2015年10期)2015-03-09
自动化博览(2014年9期)2014-02-28
