
硒代蛋氨酸对青鳉亲本繁育和子代发育的影响
2023-10-16 12:49齐澈力木格尹晓宇于永利康桂英张学富杨景峰
动物营养学报 2023年9期
王 洁 宋 磊 王 欢 齐澈力木格 尹晓宇 于永利,2 康桂英 张学富 董 武* 杨景峰*
(1.内蒙古民族大学动物科技学院,内蒙古自治区毒物监控及毒理学重点实验室,通辽 028000;2.内蒙古民族大学生命科学与食品学院,通辽 028000;3.内蒙古民族大学医学院,通辽 028000)
硒(Se)是生物体功能所必需的元素,在体内发挥重要的生物学功能。在畜牧业中,富硒产品常被用作膳食补充剂,并纳入动物预防保健的功能性饲料中[1]。但当补充的硒浓度超过所需浓度的2倍时,就会产生相应的器官毒性[2]。研究发现,鲤鱼体内的硒代蛋氨酸(Se-Met)主要在卵巢中沉积[3]。斑马鱼胚胎暴露于12.5、25.0、50.0和100.0 μg/L硒120 d,发现亚硒酸盐的存在显著抑制了斑马鱼体内卵母细胞成熟,并增加了卵母细胞凋亡[4]。在含3.7~26.6 μg/(g d)有机硒(Se-Met)饲料组中,观察到鱼体内雌二醇(E2)含量降低,并伴随着产卵率显著下降,造成严重的生殖障碍,这种障碍可能是Se-Met转移到子代,进而对子代产生毒性[5]。Chernick等[6]通过给青鳉(Oryziaslatipes)饲喂Se-Met,导致子代青鳉骨骼发育障碍。大型蚤对Se-Met更为敏感,过量Se-Met处理会导致其生殖停滞甚至死亡[7]。生殖机能与雌二醇含量以及细胞色素P450(CYP)19编码的芳香化酶密切相关[8]。在青鳉体内存在2个CYP19同源基因,其中CYP19a在卵巢中高表达,CYP19b在大脑中高表达[9]。性腺发育受下丘脑-垂体-性腺(HPG)轴调节[10]。大脑中合成并分泌促性腺激素释放激素(GnRH),刺激垂体合成和释放垂体促性腺激素(GTHs),从而刺激性腺中的类固醇生成[11]。同时,卵泡刺激素(FSH)和黄体生成素(LH)对性腺的发育也起到至关重要的作用[12]。研究发现,硒在“水体浮游动植物-鱼-水鸟”食物链中积累,导致水鸟出现筑巢率、繁殖率和胚胎发育率降低等中毒现象[13]。
青鳉属于异性繁殖,其个体卵泡的卵黄发生过程和相关卵母细胞的成熟迅速,便于观察[14-15]。目前关于Se-Met对鱼类生殖发育影响的报道较少。因此,本研究以青鳉为试验动物,用添加Se-Met的饲料饲喂F0代亲本,探讨Se-Met暴露对青鳉性行为学变化、性腺组织学变化及性腺关联基因表达变化的影响,评估Se-Met对青鳉繁殖的影响,并探讨其机制。
1 材料与方法
1.1 仪器与试剂
青鳉养殖系统购买于北京爱生公司。小型鱼行为系统购买于法国VIEWPOINT公司。Se-Met购买于美国Sigma Aldrich公司。
1.2 试验设计和饲养管理
按照楚文庆等[16]的方法和结果制备添加Se-Met的饲料和设置Se-Met暴露浓度,即2、10和50 μg/g Se-Met。首先配制含有1 mg/mL Se-Met的储备溶液(溶剂为超纯水),然后在玻璃培养皿中使用不同浓度的Se-Met稀释液与20 g的干饲料充分混合,并用刮刀彻底混合以确保液体均匀分布。分别制作含2、10和 50 μg/g Se-Met的干饲料(即1 g干饲料中含有2、10和 50 μg Se-Met),用于成年青鳉饲喂。
从繁育群体中随机选择4~5月龄的性成熟雄性和雌性青鳉,将雄性和雌性青鳉分离并观察1周,通过确认可存活的胚胎来确定个体的生殖状态。将选定的青鳉转移到试验缸,并参考自然环境冬季的光照条件保持14 h∶10 h的光照/黑暗时间。在3 L的鱼缸中分别放入雄性(5条/缸)和雌性青鳉(3条/缸),在24 ℃条件下孵育。试验缸内水体使用前曝气并过夜,配制成1‰盐水。把17条青鳉随机分配到4个组,每组3~5条鱼。对照组投喂基础干饲料,试验组分别投喂含2、10和50 μg/g Se-Met的干饲料。试验期7 d。
青鳉由内蒙古民族大学毒物监控及毒理学重点实验室提供,青鳉繁育在养殖系统中进行。成鱼保持在(28±1) ℃的封闭循环水条件下,用人工模拟光源对青鳉的生活周期进行规律性调控(白天光照14 h,晚上黑暗10 h)。水硬度为60 mg/L(以CaCO3计算);水的盐度使用NaCl调节,电导率保持在450~500 μs/cm;pH使用NaHCO3调节,保持在7.0~7.3。每天投喂3次商业干饲料和2次卤虫溶液。
试验之前,青鳉在饲养缸内适应3 d,每天饲喂3次,并补充丰年虾。在适应期后,将青鳉喂养7 d,每天投喂约1%体重的含有或不含Se-Met的干饲料,减少丰年虾的饲喂,确保饲料的摄入。喂养7 d后,使用新的鱼缸并将雌性和雄性青鳉各1条混合饲养。在繁殖期间,鱼类保持相同的饲喂时间并控制饲料用量,卤虫溶液的投喂量恢复正常以促进产卵。每日吸取粪便,以保持水体清洁,并减少缸内废物中积累的Se-Met。观察成鱼的正常活动、喂养状况和应激迹象。
1.3 指标测定
1.3.1 Se-Met在青鳉卵巢和精巢的残留
将投喂10 μg/g Se-Met干饲料7 d的青鳉解剖,取精巢和卵巢,清洗后放1.5 mL Tube管中称重,并添加10 mL优级纯硝酸转移到消解管中。将消解管放置于高温消解仪的加热块上过夜,直到内容物消失变清,然后将消解管取出并在室温下冷却约15 min。冷却后,用超纯水稀释定容消化液至50 mL。采用电感耦合等离子体质谱(ICP-MS)法测定硒浓度。
1.3.2 性腺组织发育
收集青鳉卵巢、精巢和F1代胚胎,并固定于4%多聚甲醛(PFA)溶液中4 ℃过夜,经过梯度浓度酒精脱水、置换酒精(100%酒精+100%二甲苯1次,二甲苯2次,每次10 min)后组织浸蜡包埋。获取3 μm精巢切片、5 μm卵巢切片和5 μm F1代胚胎切片,用于苏木精-伊红(HE)染色,在50 ℃的水浴中展片完全。将载玻片平放在37 ℃展片机上架子上烤干,再放入37 ℃恒温烘干箱烘干。每组3个及以上生物学重复。依据HE染色试剂盒提供方法和步骤进行染色,即常规脱蜡及梯度酒精置换二甲苯,苏木素染色2 min,水洗分化液分化3 min,之后伊红染色液染色21 s,梯度酒精脱水,二甲苯透明,干燥后,封片,并在光学显微镜下镜检观察性腺组织变化,使用Image软件进行不同发育时期卵细胞、精子计数。
1.3.3 求偶行为和游泳距离
青鳉求偶行为测试在2 L的青鳉合缸专用缸中进行。各组随机选取青鳉按雌雄比1∶1配对放入烧杯中过夜,试验于次日光照开始1 min后拍摄记录青鳉的求偶行为,记录时间为20 min。青鳉雄鱼的接近(approaching)、追随(following)、求偶方向(courting orientation)、求偶圆舞(courting round dance)以及交配(copulation)等发生频次被定量为青鳉求偶行为指标[17]。各种求偶行为参数定义如下。接近:雌性青鳉停留或者游得很慢时,可以观察到雄性青鳉通过游泳来缩短与雌性青鳉的距离。追随:通常表现为雄性青鳉跟随雌性青鳉后面游泳,二者游泳速度大致相同,2个个体之间的距离几乎不变,一般不超过5 cm。求偶方向:雄性青鳉在雌性青鳉的下面,嘴部位于雌性青鳉腹部位置,二者以相同的速率很缓慢地游动,并且方向相同,也可以理解为追求取向。求偶圆舞:一般来说,雄性青鳉在雌性青鳉周围快速移动,典型情况下,雄性青鳉在雌性青鳉前面顺时针或者逆时针绘制一个水平或者垂直的圆圈,之后快速恢复到之前的求偶方向,这种舞蹈经常连续重复几次,有时也会出现雄性青鳉只在雌性青鳉面前进行一个小小的转身,这种情况被认定为不完整的求偶舞蹈。交配:当雌性青鳉接受了雄性青鳉的求偶,二者会游至上层,尾鳍紧紧贴合,身躯水平呈60°角,全身及鳍快速而精细地颤抖,缓慢下沉,持续时间为15~30 s,到达高潮。
记录求偶行为之后,使用小型鱼行为系统分别测量各组亮-暗5 min交替下游泳距离,每组测定3个重复。
1.3.4 大脑和性腺中生殖相关基因的表达
收集青鳉大脑和性腺并切取1/2放入2.0 mL离心管中,每组测定3个重复。使用Trizol方法提取总RNA,纯化后使用ND-1000(ThermoFisher Scientifific公司,美国)进行RNA定量。使用250 ng总RNA与高容量cDNA逆转转录试剂盒进行cDNA合成。根据已在NCBI上发表的青鳉基因序列,利用Primer Premier 6.0设计实时荧光定量PCR引物,以18S为看家基因,检测CYP19a、CYP19b、GnRHb、催乳素(PRL)、FSHb、LHb以及生长乳素(SL)的mRNA相对表达水平,引物序列见表1。根据Dong等[18]的方法使用StepOnePlusTMReal-Time PCR System用于PCR扩增。PCR反应条件为:95 ℃ 15 min,95 ℃ 40个周期15 s。采用2-△△Ct方法来计算mRNA相对表达水平。
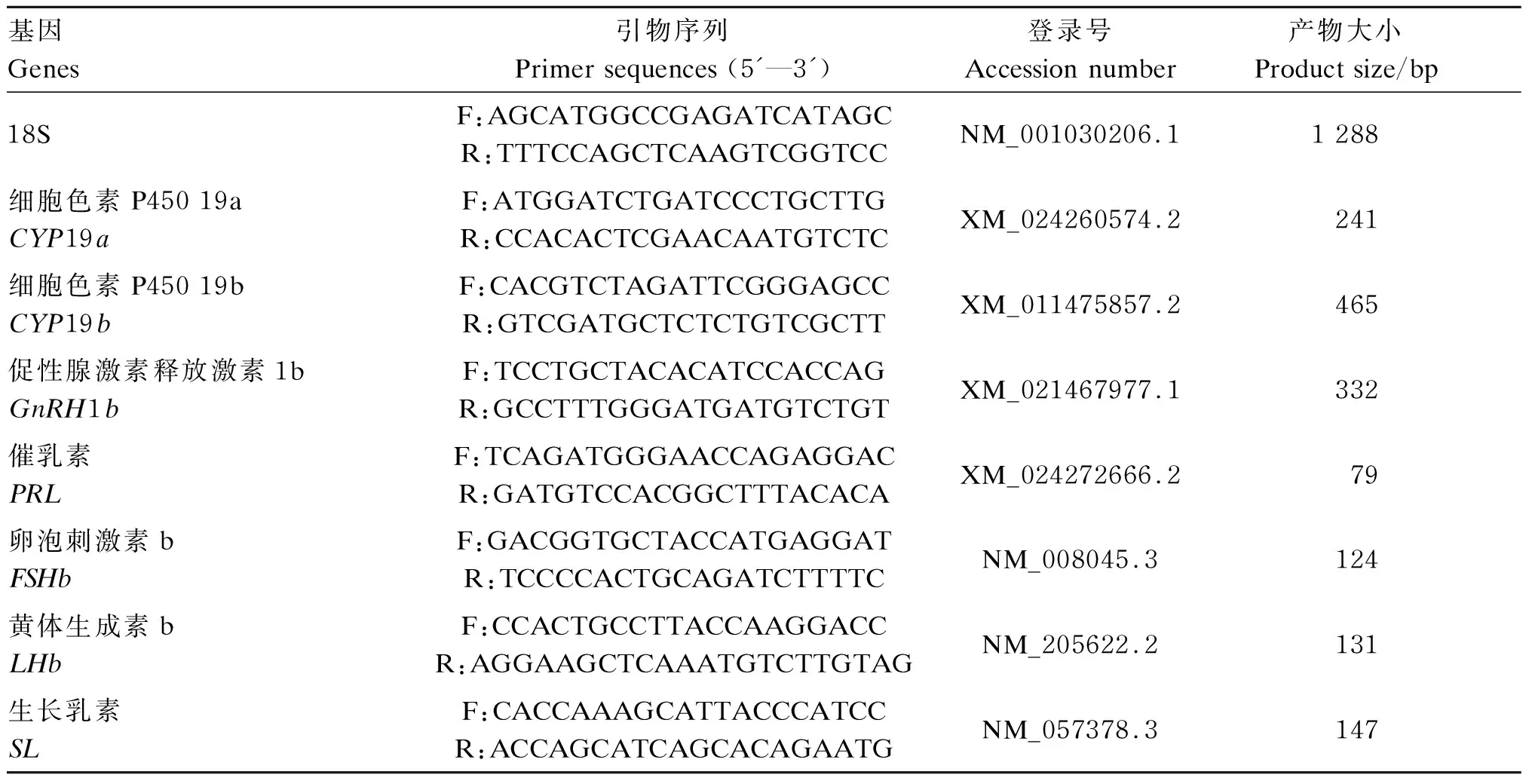
表1 引物序列
1.3.5 F1代青鳉鱼胚胎组织变化
成年青鳉按照雌性和雄性1∶1的比例进行交配产卵,12 h后收集F1代胚胎。在受精后第8天时将青鳉鱼卵置于显微镜下观察其受精和畸形情况,并记录。在第10天时,收集青鳉胚胎进行固定并制作石蜡切片进行HE染色,并进一步在光学显微镜下观察胚胎组织变化。
1.4 统计分析
试验数据统计使用SPSS 20.0分析软件进行单因素方差分析,数据用“平均值±标准误”表示,差异显著性水平设置为P<0.05。
2 结 果
2.1 Se-Met在青鳉卵巢和精巢的残留
由表2可见,对照组卵巢硒浓度为0.008 1 μg/g,10 μg/g Se-Met组卵巢硒浓度为0.016 1 μg/g;10 μg/g Se-Met组卵巢硒浓度是对照组的1.99倍,差异显著(P<0.05)。对照组精巢硒浓度为0.007 8 μg/g,10 μg/g Se-Met组精巢硒浓度为0.007 2 μg/g;10 μg/g Se-Met组精巢硒浓度与对照组无显著差异(P>0.05)。
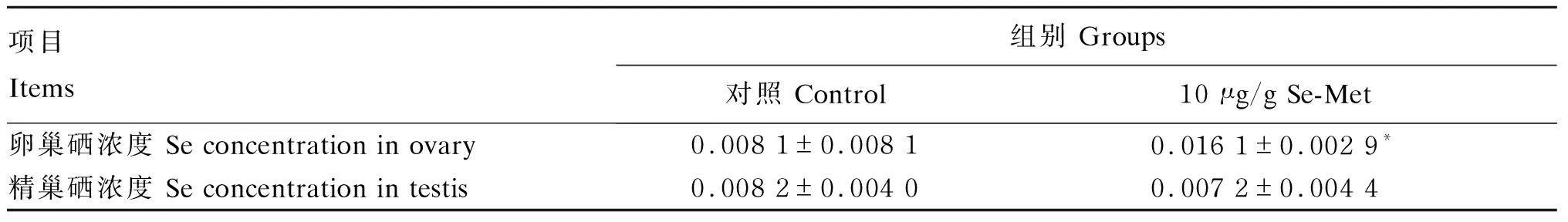
表2 Se-Met在青鳉卵巢和精巢的残留
2.2 Se-Met对青鳉求偶行为及游泳距离的影响
由图1可见,与对照组相比,2 μg/g Se-Met组的接近、追随、求偶方向、求偶圆舞和交配发生频率没有显著差异(P<0.05);10、50 μg/g Se-Met组的接近、求偶方向和求偶圆舞发生频率显著降低(P<0.05),而追随、交配发生频率没有显著差异(P>0.05)。
由表3可见,各组之间雌性和雄性青鳉的游泳距离没有显著差异(P>0.05)。
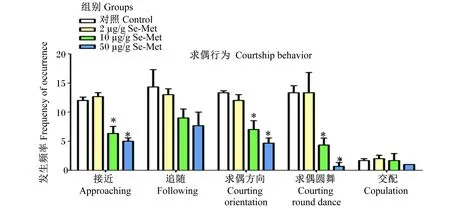
*表示与对照组相比差异显著(P<0.05)。
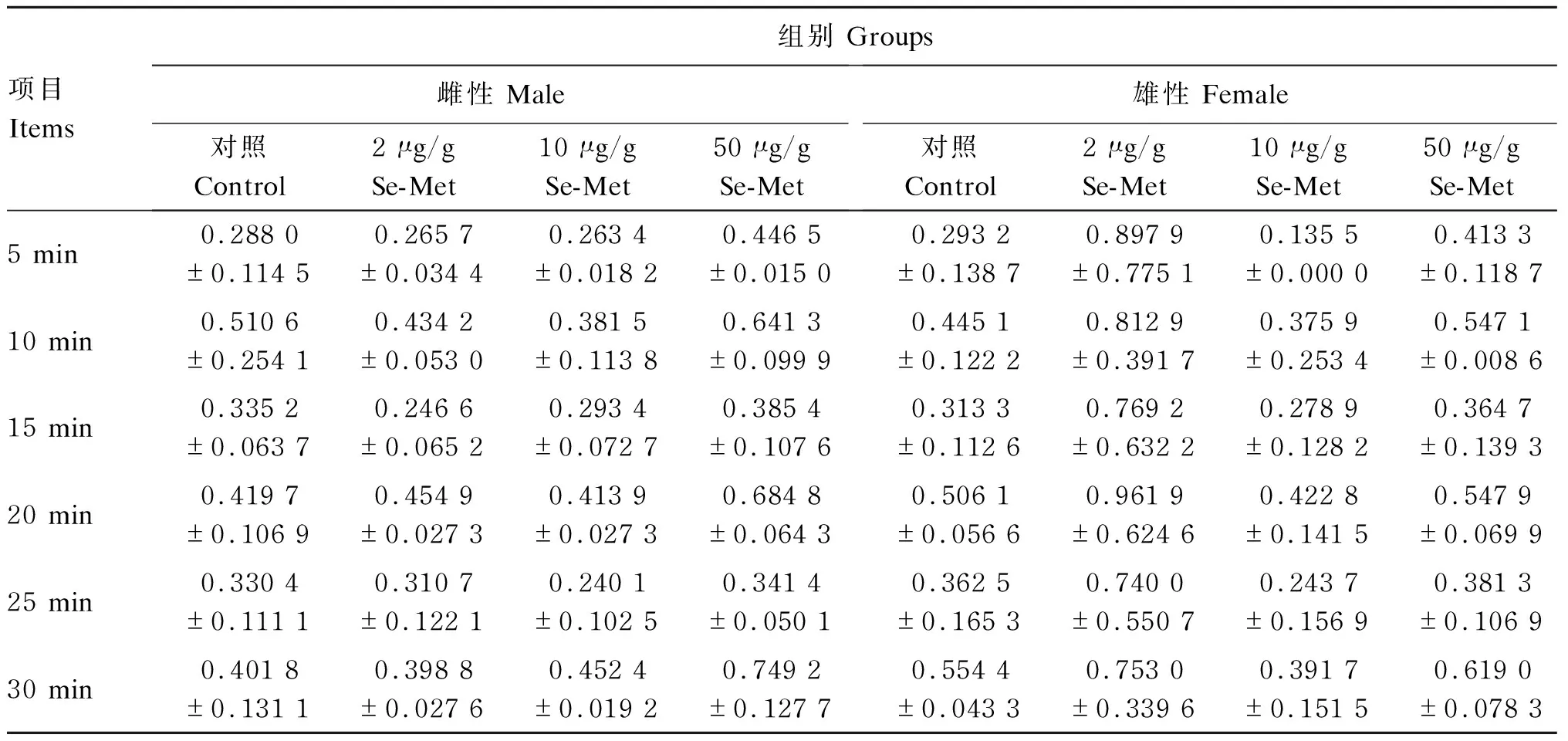
表3 Se-Met对青鳉游泳距离的影响
2.3 Se-Met对青鳉性腺组织发育的影响
正常青鳉卵巢组织由不同发育时期的卵细胞组成,根据张静等[19]的研究将卵巢分为5个发育时期,Ⅰ期:染色质核仁期;Ⅱ期:绒毛膜期;Ⅲ期:细胞核核仁期;Ⅳ期:卵黄形成期;Ⅴ期:成熟卵期。青鳉卵巢切片染色结果显示,Ⅰ~Ⅳ期卵母细胞呈现不同程度的堆积,相反,Ⅴ期卵母细胞数量呈现降低的趋势,并且都出现不同程度的卵泡排列以及结构凌乱等卵巢萎缩现象(图2-A~图2-D)。与对照组相比,50 μg/g Se-Met组的Ⅲ期卵细胞计数显著升高(P<0.05)(表4)。
雄性青鳉性腺组织发育良好,在精巢组织中可以看到精原细胞均分布于官腔中央及大小不一的生精小管中,随着Se-Met浓度的升高,青鳉精原细胞数呈降低趋势(图2-E~图2-H)。与对照组相比,10和50 μg/g Se-Met组的精子计数显著降低(P<0.05)(表4)。
2.4 Se-Met对青鳉大脑和卵巢中生殖相关基因表达的影响
由表5可见,与对照组相比,2 μg/g Se-Met组大脑和卵巢中各生殖相关基因的mRNA相对表达水平无显著差异(P>0.05);10和50 μg/g Se-Met组雌鱼和雄鱼大脑中CYP19b的mRNA相对表达水平显著升高(P<0.05),10和50 μg/g Se-Met组卵巢中CYP19a的mRNA相对表达水平显著升高(P<0.05),10和50 μg/g Se-Met组卵巢中FSHb的mRNA相对表达水平显著升高(P<0.05),50 μg/g Se-Met组卵巢中GnRH1b的mRNA相对表达水平显著升高(P<0.05),50 μg/g Se-Met组卵巢中LHb的mRNA相对表达水平显著升高(P<0.05),10和50 μg/g Se-Met组卵巢中SL的mRNA相对表达水平显著降低(P<0.05)。
2.5 Se-Met对F1代青鳉胚胎状态的影响
由表6可见,与对照组相比,2 μg/g Se-Met组青鳉胚胎的未受精率和畸形率没有显著差异(P>0.05),而50 μg/g Se-Met组青鳉胚胎的未受精率显著升高(P<0.05),10和50 μg/g Se-Met组青鳉胚胎的畸形率显著升高(P<0.05)。其中,畸形主要表现为头部浮肿、心囊浮肿、双头鱼等(图3-A~图3-F)。
3 讨 论
硒作为生物体内必需的微量营养素,其主要功能是维持正常的新陈代谢[20]。硒缺乏与认知和神经运动障碍有关,且可引起黄鲶幼鱼体内活性氧(ROS)的过量产生,抑制其生长并引起氧化应激[21-22]。因此,外源硒的补充被认为是对鱼类健康有益的[23]。与无机形式的硒相比,有机硒具有更好的生物利用度[24]。但过度补硒也会造成硒中毒。在内分泌组织(甲状腺、肾上腺、垂体、睾丸、卵巢)中的硒含量高于其他许多器官,研究发现,精子的发生依赖于充足的硒供应,而硒过量可能损害卵巢功能[25]。应强调的是,硒含量过高或缺乏硒对人体健康有害,而硒元素在体内正常运作所需的剂量与有害剂量之间的差异很小,因此必须谨慎地补充。本研究发现过量Se-Met对青鳉的生殖毒性,检测到Se-Met在青鳉卵巢内的残留,并首次发现Se-Met喂饲造成青鳉求偶行为的减弱,也造成成熟卵母细胞减少、未成熟卵母细胞大量堆积、卵巢及卵细胞形态紊乱。结果提示,合理控制硒的摄入是非常重要的课题。
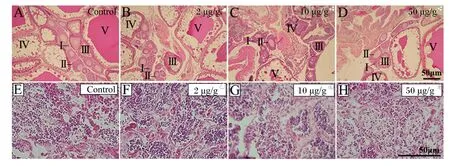
A~D分别为对照组、2 μg/g Se-Met组、10 μg/g Se-Met组、50 μg/g Se-Met组的卵巢。其中罗马数字Ⅰ~Ⅴ分别对应Ⅰ期:染色质核仁期;Ⅱ期:绒毛膜期;Ⅲ期:细胞核核仁期;Ⅳ期:卵黄形成期;Ⅴ期:成熟卵期。E~H分别为对照组、2 μg/g Se-Met组、10 μg/g Se-Met组、50 μg/g Se-Met组的精巢。比例尺为50 μm。
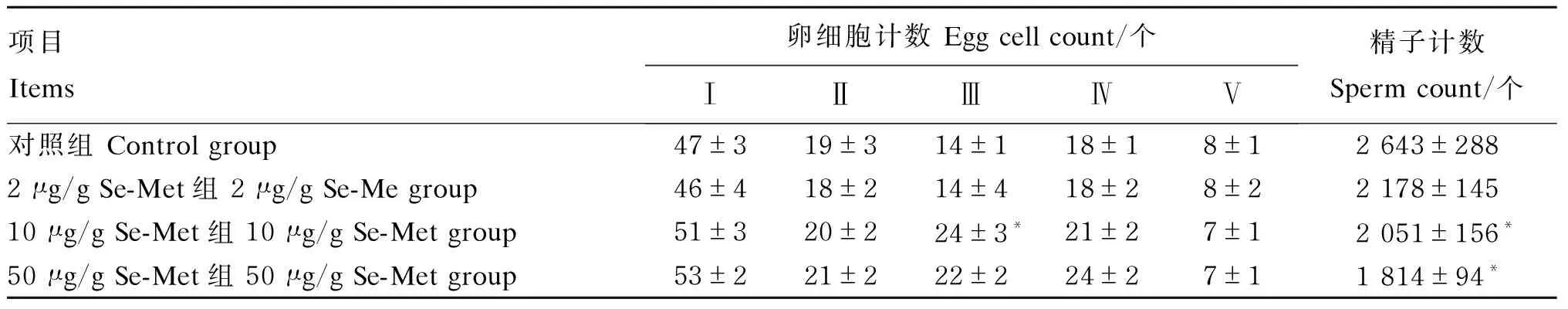
表4 Se-Met对青鳉卵细胞和精子计数的影响
Naderi等[26]给斑马鱼进行了为期60 d的Se-Met(3.5、11.1、27.4和63.4 μg/g硒)暴露,Se-Met导致斑马鱼联想学习障碍,并阐明这种障碍与巴胺能神经传递的失调有关。斑马鱼的神经系统对外源性毒物敏感,特别是在胚胎神经发生过程中,在过量硒处理的胚胎中观察到斑马鱼运动行为失调,包括运动距离缩短和触觉反应迟钝[27]。但目前尚未发现Se-Met改变求偶行为的报道,本试验首次发现Se-Met对青鳉求偶行为产生了影响,我们猜测这可能与体内激素紊乱有关。
生殖内分泌紊乱对鱼类繁殖的影响主要表现为雄性精子密度、活力下降;雌性卵母细胞生长和成熟过程停滞,从而影响配子活力和生育能力[28]。斑马鱼暴露于草甘膦类除草剂(0.065和6.500 mg/L)15 d后,出现初期卵泡数量增加、晚期卵泡数量减少等对卵巢成熟不利的现象,并且表明这些变化可能导致生殖毒性发生[29]。本研究发现,青鳉在使用10和50 μg/g Se-Met处理后,卵巢组织中成熟的Ⅳ期细胞数量减少,而相应的Ⅰ、Ⅱ、Ⅲ期的卵母细胞出现堆积,这可能是由于过量Se-Met引起激素紊乱导致卵巢功能损伤。给予0.2、0.4或0.8 mg/kg BW硒可促进大鼠精子活力,而过量的硒补充(4.0和8.0 mg/kg BW)导致大鼠睾丸重量、精子浓度和活力下降,从而引起睾丸和附睾的组织病理学损伤[30]。在本研究中也发现了相似的变化,随着Se-Met浓度的升高,青鳉精子计数呈降低趋势。
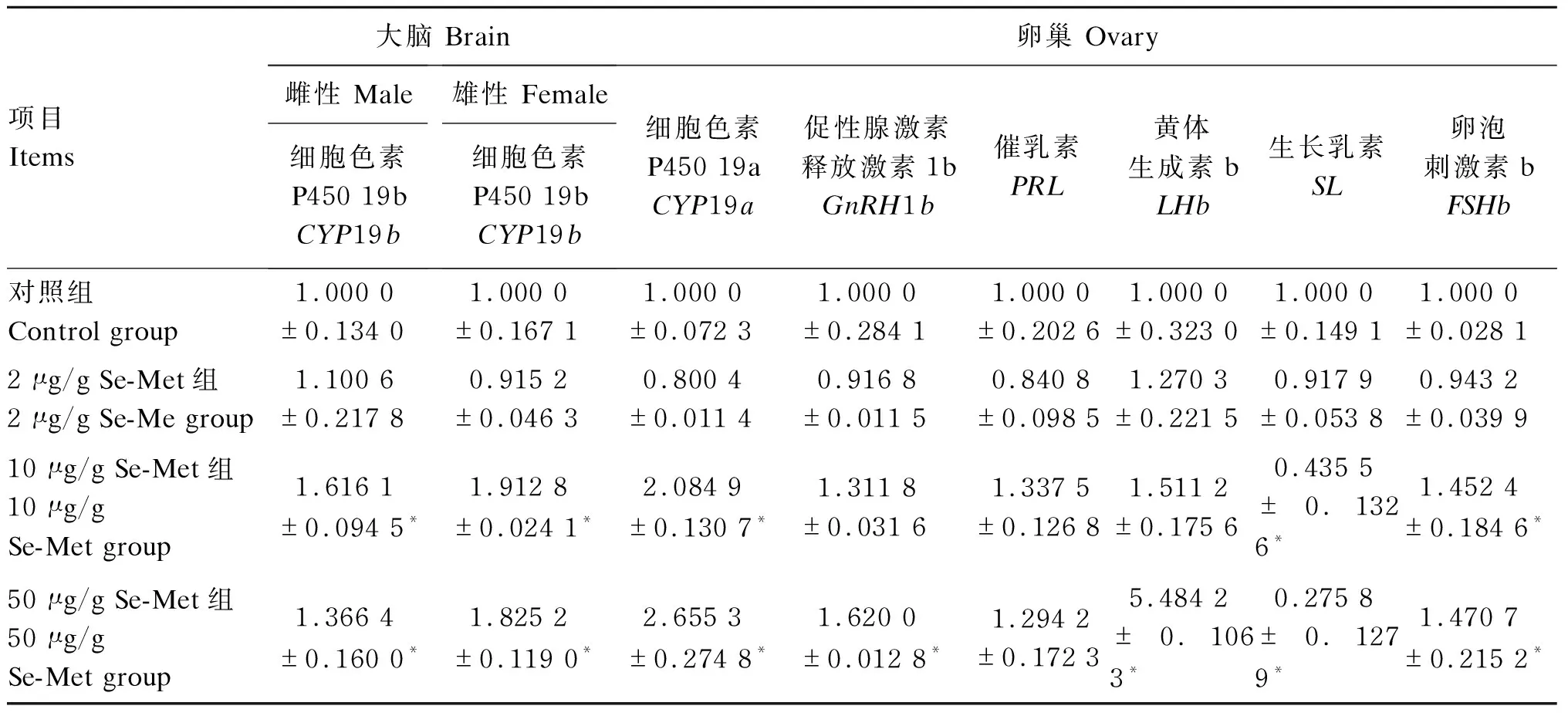
表5 Se-Met对青鳉大脑、卵巢中生殖相关基因表达的影响
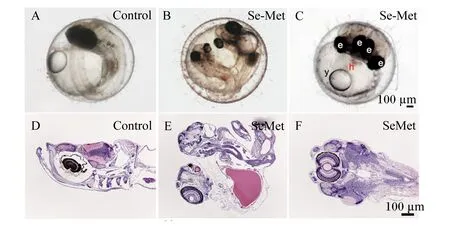
Control:对照组;Se-Met:50 μg/g Se-Met组。图A:正常受精卵;图B:心囊浮肿畸形受精卵;图C:双头畸形受精卵;图D、图E和图F分别是图A、图B和图C的组织切片HE染色图。e:眼睛;y:卵黄囊;h:心脏。
过量的硒摄入被发现具有致畸风险,由于可以通过母体的屏障渗透到胎盘和乳腺中,因此很多新生动物会出现发育异常、畸形率增高等硒中毒症状[31]。Gerhart等[32]持续28 d给黑头鲦鱼饲喂富硒饲料(1.21~26.5 μg/g硒),母体没有产生明显的生殖毒性,但却检测到胚胎硒浓度以剂量依赖性方式增加,并出现大量畸形,但不影响胚胎的孵化率或存活率。Chernick等[6]向成年青鳉饲料中添加Se-Met(12.5、25或50 μg/g),连续饲喂6 d后实施交配,所产胚胎孵化率降低,出现较多的畸形。这些研究都阐述了Se-Met饲喂造成子代的影响。这与本研究检测的过量Se-Met补充造成子代畸形率及死亡率升高的结果相似。但目前Se-Met对母体卵巢和精巢的影响并没有报道。本试验首次进行了求偶行为和性腺的组织学变化的研究。
由CYP19基因编码的芳香酶主要功能为促进雄激素向雌性激素的转化,以此来维持体内性类固醇水平的稳定,青鳉基因组存在2个芳香酶同源基因,分别为CYP19a和CYP19b,其中CYP19a在卵巢中高表达[33]。Nikhil等[34]发现硒和铜的暴露可刺激羊卵巢颗粒细胞CYP19a1的表达增加。性腺的CYP19a是类固醇生成途径中的末端酶,并催化雄激素向雌激素的转变,因此,CYP19amRNA标记表达可显著干扰性类固醇代谢,引起激素失衡[35]。与之前的报告结果相同,本试验检测到在低浓度Se-Met处理后,卵巢中CYP19a的mRNA相对表达水平有下调的趋势;而高浓度Se-Met处理后,CYP19a的mRNA相对表达水平升高,这可能暗示着卵巢诱导CYP19a表达是对芳香酶抑制的常见代偿反应。与斑马鱼相似,青鳉体内CYP19b对雌激素激活比CYP19a更敏感[36]。CYP19b通常认为与鱼类的繁育和求偶行为有密切关联。有研究显示,雌激素及类污染物造成鱼类大脑中CYP19b表达的上调[37-39]。但我们没有发现Se-Met对CYP19b表达影响的报道。在本研究中,Se-Met处理的鱼卵巢中CYP19b的mRNA相对表达水平升高,表明雌激素和雄激素水平的平衡可能受到干扰。
生殖功能是由HPG轴,也称为GnRH-促性腺激素类固醇轴调控。GnRH在HPG轴控制生殖内分泌方面也发挥重要作用[40]。GnRH可介导FSH和LH的分泌[41]。FSH和LH在卵泡发育的阶段主要依赖于细胞外和细胞内分子相互作用的网络。靶组织(性腺)对FSH和LH的反应性在很大程度上取决于其特异性受体的表达[42]。在自然排卵周期中FSH和LH水平出现周期激增,LH水平的增高可促进卵母细胞成熟并触发排卵,然而,FSH水平的过度升高与萎缩性卵巢有关[43]。暴露于有毒环境下,化学物质可以通过影响激素作用或干扰下丘脑和/或垂体影响卵巢功能,甚至通过破坏滤泡直接导致卵巢衰竭,最终破坏神经内分泌反馈,导致FSH和LH水平升高[44]。Peng等[45]研究也表明,GnRH以及FSH的表达上调可能是由毒性物质引起的GnRH负反馈调节导致。在本研究中,高浓度的Se-Met造成卵巢卵泡发育障碍,同时也检测到GnRH、FSH和LH的mRNA相对表达水平上调。
在脊椎动物中,PRL主要由垂体泌乳素合成和释放,并在生长发育、代谢、泌乳、免疫调节、行为和繁殖过程中发挥重要作用[46]。研究表明,PRL基因在卵巢疾病中起着重要的作用。接触内分泌干扰物的鲶鱼卵巢中PRL水平升高[47]。本研究中发现,Se-Met没有显著引PRL的mRNA相对表达水平升高。PRL在鱼类中的作用还有待观察,但目前的研究可能暗示这种现象的发生。SL是一种鱼类特异性腺垂体肽激素,属于生长激素(GH)/PRL家族[48]。该激素被认为在早期卵子生成、性腺成熟和性腺类固醇生物合成期间起作用。SL可通过调节脂质代谢来促进卵母细胞成熟,血浆SL水平在性成熟过程中升高[49]。借助转录组学技术发现,SL可能在卵母细胞发育的早期阶段发挥作用,因为卵巢SLR转录本随着卵母细胞成熟而下调[50]。此外,有研究表明,SL表达的降低可能会促进GnRH的表达[51]。这与本研究结果相符,本研究发现过量Se-Met造成青鳉求偶行为指标的低下,却同时造成生殖关联基因表达的增加,这可能是一种负反馈调节机制。虽然硒补充产品在水产养殖方面广泛被应用,但硒的剂量选择之间的临界值非常小,因为稍高的剂量就可能导致硒中毒。
4 结 论
Se-Met影响成年青鳉的繁育,造成其求偶行为指标降低,卵巢组织中成熟卵泡数量减少和未成熟卵母细胞堆积,精原细胞数量减少。这种变化可能是由于Se-Met影响了CYP19b、CYP19a、GnRH1b、PRL、LHb、SL及FSHb表达而造成。本研究结果提示,给鱼类添加Se-Met时要严格控制使用剂量。
猜你喜欢
东坡赤壁诗词(2022年4期)2022-10-30
文萃报·周五版(2022年24期)2022-06-21
植物保护(2019年4期)2019-09-04
安徽农业科学(2018年22期)2018-05-14
河南畜牧兽医(2017年8期)2017-11-24
上海农业学报(2017年3期)2017-04-10
华南师范大学学报(自然科学版)(2014年5期)2014-08-28
食品工业科技(2014年15期)2014-03-11
微创泌尿外科杂志(2014年4期)2014-02-28
生殖医学杂志(2013年5期)2013-03-11
