
高精料饲粮中添加烟酸对绵羊瘤胃内乳酸代谢及其相关酶活性的影响
2023-11-10 06:09成志强管勤超臧长江陈开旭刘建成焦毅灵李凤鸣李晓斌杨开伦雒秋江
动物营养学报 2023年10期
成志强 管勤超 臧长江 陈开旭 刘建成 焦毅灵 李凤鸣 李晓斌 杨开伦 雒秋江
(新疆农业大学动物科学学院,新疆肉乳用草食动物营养实验室,乌鲁木齐 830052)
烟酸属于B族维生素,主要有烟酰胺腺嘌呤二核苷酸(NAD)和烟酰胺腺嘌呤二核苷酸磷酸(NADP)2种形式[1-2]。这2种辅酶在机体中参与脂肪酸、碳水化合物和蛋白质的代谢。反刍动物饲粮中的碳水化合物经过瘤胃微生物发酵降解为葡萄糖,葡萄糖进一步生成丙酮酸。一方面,丙酮酸在乳酸脱氢酶的作用下通过“丙酮酸+还原态烟酰胺腺嘌呤二核苷酸(NADH)+氢离子(H+)↔乳酸+氧化态烟酰胺腺嘌呤二核苷酸(NAD+)”反应生成乳酸[3-4];另一方面,丙酮酸通过丙酮酸羧化酶进一步生成草酰乙酸,再经苹果酸脱氢酶和琥珀酸脱氢酶作用下生成相对应的苹果酸和琥珀酸,最终生成丙酸,为机体提供能量[5-6]。在正常生理条件下,反刍动物机体合成的烟酸可以满足自身的需求,然而,在高精料饲粮条件下,反刍动物体内持续产生乳酸,由于NAD+的缺乏,使反刍动物对烟酸的需要量增加,动物机体合成的烟酸不能满足自身的需要量,烟酸的缺乏会导致反刍动物瘤胃乳酸蓄积,造成瘤胃酸中毒[7-9]。
饲粮中精料比例过高,奶牛瘤胃内D-乳酸浓度显著增加[10]。烟酸作为NAD+的前体物质,参与丙酮酸与乳酸的相互转化,在高精料饲粮条件下,外源性添加的烟酸提供的NAD+可以促使反应向生成丙酮酸的方向进行,减少乳酸的蓄积,缓解瘤胃酸中毒。已有研究表明,在高精料饲粮中添加800 mg/kg的烟酸可以显著降低锦江牛瘤胃内乳酸浓度,并且抑制乳酸脱氢酶(LDH)的活性[11];在高精料饲粮中添加烟酸能显著降低黄牛瘤胃内乳酸浓度[12]。目前,大多数研究主要集中在高精料饲粮中添加烟酸对反刍动物瘤胃内乳酸浓度的影响,而对乳酸代谢相关酶及碳水化合物代谢途径中间代谢产物的研究鲜有报道。本试验拟从NAD+的前体物质烟酸的角度出发,探究高精料饲粮条件下添加烟酸对绵羊瘤胃内乳酸代谢相关酶和能量代谢途径中间代谢产物的影响,为烟酸在反刍动物实际生产中的应用提供参考。
1 材料与方法
1.1 试验设计
选取体重[(36.04±1.81) kg]、体况相近且健康的8月龄哈萨克公羊24只,试验前安装好永久性瘤胃瘘管,随机分为4组,每组6只。对照组饲喂精粗比为70∶30的基础饲粮,试验Ⅰ、Ⅱ、Ⅲ组分别在对照组饲粮的基础上添加100、130和160 mg/d烟酸(烟酸购自上海源叶生物科技有限公司,分析纯,纯度≥99%)。试验期共18 d,其中预试期14 d,正试期4 d。
1.2 基础饲粮
试验用基础饲粮参照《肉羊饲养标准》(NY/T 816—2004),以日增重200 g营养需求的1.2倍水平进行配制,其组成及营养水平见表1。
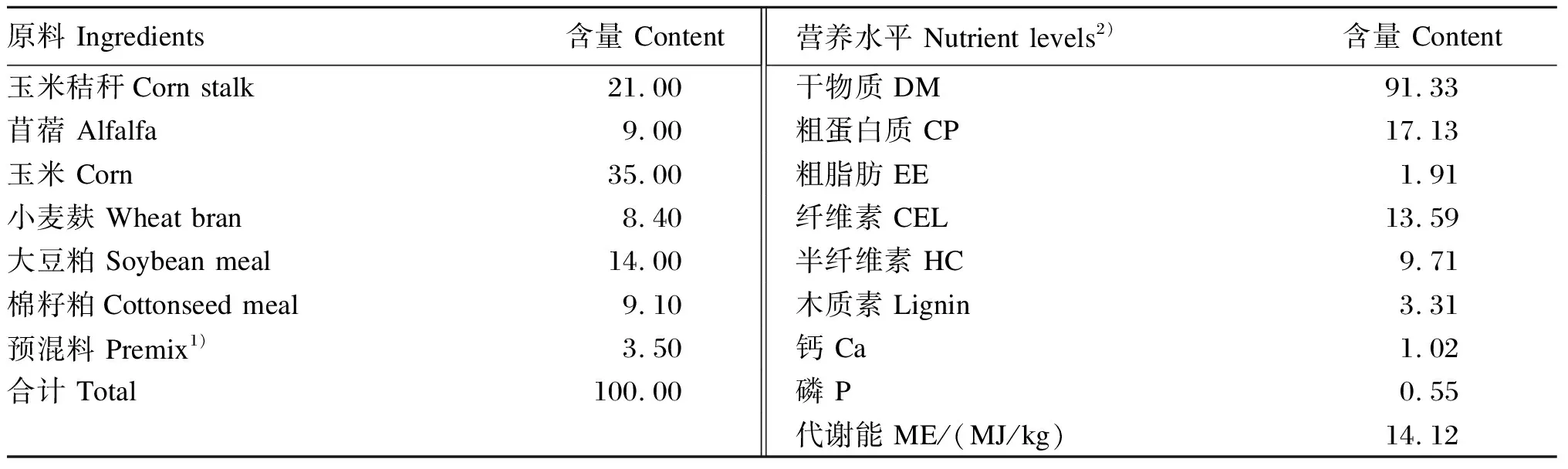
表1 基础饲粮组成及营养水平(干物质基础)
1.3 饲养管理
试验绵羊采用单栏饲养,试验前进行统一驱虫,对圈舍进行消毒处理。每日分别于07:00和19:00进行饲喂,每次饲喂量为当日饲喂总量的1/2,自由采食,自由饮水,保持圈舍光照、干燥和通风。
1.4 样品采集
于正试期每天07:00饲喂前0 h以及饲喂后1、3、5 h采集血液样品。使用肝素钠抗凝管采集颈静脉血样9 mL,在4 ℃条件下3 000 r/min离心15 min分离血清,分装于1.5 mL的Eppendorf管中,-20 ℃冷冻保存。
于正试期每天07:00饲喂前0 h以及饲喂后1、3、5 h采集瘤胃液样品。为了避免瘤胃液污染,丢弃前50 mL瘤胃液样品,将采集的80 mL瘤胃液通过4层尼龙布过滤,分装在冻存管中于-20 ℃冷冻保存。
1.5 指标测定
1.5.1 丙酮酸、苹果酸、琥珀酸浓度的测定
丙酮酸(PA)、苹果酸(MA)和琥珀酸(SA)浓度的测定参考张艳艳等[13]的方法,使用高效液相色谱仪(岛津,日本)测定。取瘤胃液1 mL至离心管,静置10 min后,12 000 r/min离心20 min;取500 μL上清至另一离心管,再加入500 μL10%三氯乙酸(TCA),静置5 min后12 000 r/min离心10 min,取20 μL上机测定。测定条件:使用Shim-pack GISTC18-AQ色谱柱(5 μm,4.6 mm×250 mm);流速1.0 mL/min;柱温30 ℃;检测波长210 nm;一次线性洗脱;进样量20 μL。
1.5.2L-乳酸、D-乳酸浓度及乳酸代谢相关酶活性的测定
瘤胃液及血清中L-乳酸、D-乳酸浓度分别采用分光光度法和酶联免疫吸附测定法进行测定,瘤胃液中丙酮酸激酶(PK)、乳酸脱氢酶(LDH)、苹果酸脱氢酶(MDH)、琥珀酸脱氢酶(SDH)活性与NADH、NAD+浓度采用分光光度法测定。以上指标测定所用试剂盒购自南京建成生物工程研究所。
1.6 数据处理与分析
用Excel 2013对试验数据进行初步整理,统计分析采用SPSS 19.0统计软件中的单因素方差分析程序进行,多重比较采用Duncan氏法进行,试验结果均以平均值表示,各组的变异程度用均值标准误(SEM)表示,差异显著性判定标准为P<0.05表示差异显著,P<0.01为差异极显著。
2 结果与分析
2.1 高精料饲粮中添加烟酸对绵羊瘤胃液中L-乳酸、D-乳酸浓度的影响
由表2可知,对于瘤胃液中L-乳酸浓度,与对照组相比,试验Ⅱ组在饲喂后1 h时显著降低(P<0.05);试验Ⅱ、Ⅲ组在饲喂后3 h时极显著降低(P<0.01),试验Ⅰ组在饲喂后3 h时显著降低(P<0.05),同时试验Ⅱ组显著低于试验Ⅰ组(P<0.05)。对于瘤胃液中D-乳酸浓度,与对照组和试验Ⅰ组相比,试验Ⅱ组在饲喂后1 h时极显著降低(P<0.01),试验Ⅱ组在饲喂后5 h时显著降低(P<0.05)。其余时间点各组间瘤胃液中L-乳酸、D-乳酸浓度差异均不显著(P>0.05)。
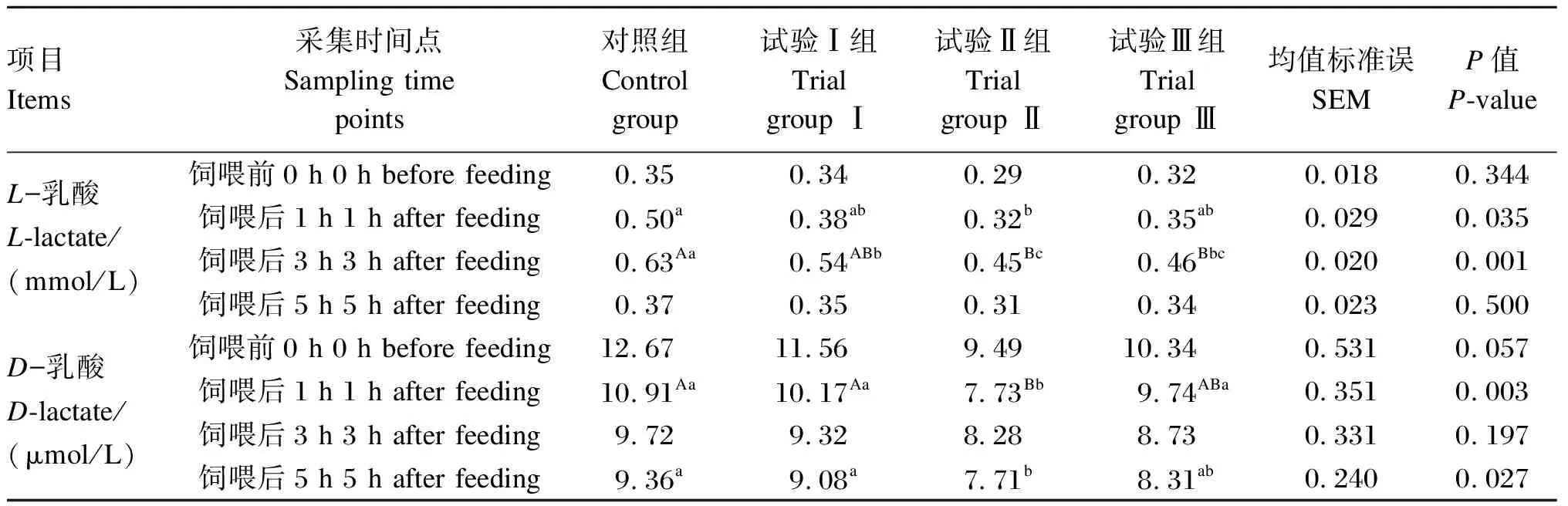
表2 高精料饲粮中添加烟酸对绵羊瘤胃液中L-乳酸、D-乳酸浓度的影响
2.2 高精料饲粮中添加烟酸对绵羊血清中L-乳酸、D-乳酸浓度的影响
由表3可知,与对照组相比,在饲喂后3 h时,试验Ⅱ、Ⅲ组血清中L-乳酸浓度极显著降低(P<0.01),血清中D-乳酸浓度显著降低(P<0.05)。其余时间点各组间血清中L-乳酸、D-乳酸浓度差异均不显著(P>0.05)。

表3 高精料饲粮中添加烟酸对绵羊血清中L-乳酸、D-乳酸浓度的影响
2.3 高精料饲粮中添加烟酸对绵羊瘤胃液中NAD+、NADH浓度的影响
由表4可知,在饲喂后3 h时,试验Ⅱ组瘤胃液中NAD+浓度极显著高于对照组(P<0.01),试验Ⅲ组显著高于对照组(P<0.05);在饲喂后3h时,对照组瘤胃液中NADH浓度极显著高于试验Ⅱ、Ⅲ组(P<0.01),显著高于试验Ⅰ组(P<0.05)。其余时间点瘤胃液中NAD+、NADH浓度各组间差异均不显著(P>0.05)。

表4 高精料饲粮中添加烟酸对绵羊瘤胃液中NAD+、NADH浓度的影响
2.4 高精料饲粮中添加烟酸对绵羊瘤胃液中乳酸脱氢酶、丙酮酸激酶、苹果酸脱氢酶和琥珀酸脱氢酶活性的影响
由表5可知,瘤胃液中乳酸脱氢酶活性,在饲喂后1 h时对照组显著高于试验Ⅱ组(P<0.05),在饲喂后3 h时对照组极显著高于试验Ⅱ、Ⅲ组(P<0.01),试验Ⅰ组显著高于试验Ⅱ、Ⅲ组(P<0.05),在饲喂后5 h时对照组极显著高于试验Ⅱ、Ⅲ组(P<0.01),试验Ⅰ组显著高于试验Ⅱ组(P<0.05);瘤胃液中丙酮酸激酶活性,在饲喂前0 h和饲喂后1 h时试验Ⅱ组极显著高于对照组(P<0.01),显著高于试验Ⅰ组(P<0.05);瘤胃液中苹果酸脱氢酶活性在饲喂后1 h时试验Ⅱ组极显著高于对照组和试验Ⅰ组(P<0.01),显著高于试验Ⅲ组(P<0.05);瘤胃液中琥珀酸脱氢酶活性,在饲喂后1 h时试验Ⅱ组极显著高于对照组(P<0.01),显著高于试验Ⅰ组(P<0.05),在饲喂后3 h时试验Ⅱ组显著高于对照组(P<0.05)。其余时间点瘤胃液中乳酸脱氢酶、丙酮酸激酶、苹果酸脱氢酶和琥珀酸脱氢酶活性各组间差异均不显著(P>0.05)。
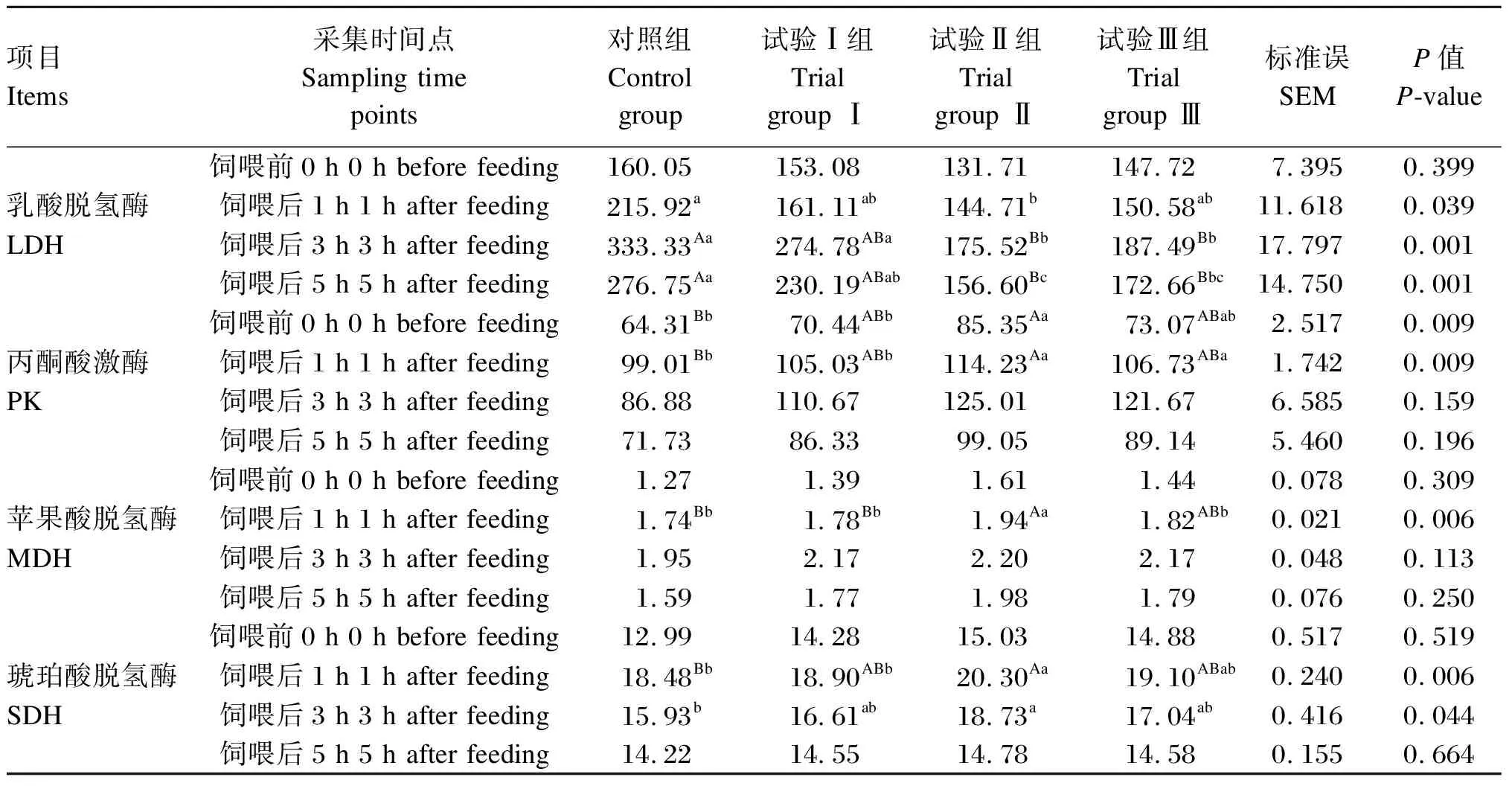
表5 高精料饲粮中添加烟酸对绵羊瘤胃液中乳酸脱氢酶、丙酮酸激酶、苹果酸脱氢酶和琥珀酸脱氢酶活性的影响
2.5 高精料饲粮中添加烟酸对绵羊瘤胃液中丙酮酸、苹果酸和琥珀酸浓度的影响
由表6可知,瘤胃液中丙酮酸浓度,在饲喂前0 h时试验Ⅱ、Ⅲ组显著高于对照组(P<0.05),在饲喂后1 h时试验Ⅲ组显著高于对照组(P<0.05),试验Ⅱ组极显著高于对照组(P<0.01);瘤胃液中苹果酸浓度,在饲喂后1 h时试验Ⅱ、Ⅲ组显著高于对照组(P<0.05);瘤胃液中琥珀酸浓度,在饲喂后1 h时试验Ⅱ、Ⅲ组极显著高于对照组(P<0.01),试验Ⅰ组显著高于对照组(P<0.05),在饲喂后3 h时试验Ⅱ组极显著高于对照组(P<0.01),试验Ⅲ组显著高于对照组(P<0.05)。其余时间点瘤胃液中丙酮酸、苹果酸、琥珀酸浓度各组间差异均不显著(P>0.05)。
3 讨 论
3.1 高精料饲粮中添加烟酸对绵羊瘤胃液和血清中L-乳酸、D-乳酸浓度的影响
乳酸是反刍动物瘤胃内碳水化合物代谢过程的中间代谢产物[14],瘤胃液中乳酸浓度的高低是衡量瘤胃功能是否正常的重要指标。在高精料饲粮条件下发生瘤胃酸中毒时,随着乳酸的积累,瘤胃液中L-乳酸浓度较低,D-乳酸浓度逐渐升高[15],这是因为瘤胃内广泛存在L-乳酸脱氢酶,L-乳酸脱氢酶催化L-乳酸通过丙烯酸途径转化为丙酸;而机体中缺乏D-乳酸脱氢酶,大量D-乳酸蓄积在瘤胃内,只有少量D-乳酸通过瘤胃上皮细胞经过D-乳酸脱氢酶进一步代谢出瘤胃,最终进入血液[16]。本试验结果表明,在饲喂后1、3 h时,各试验组瘤胃内L-乳酸、D-乳酸浓度均低于对照组,其中试验Ⅱ组最低。这可能是因为试验组饲粮添加烟酸后,烟酸作为NAD+的前体物质,提供大量的NAD+,使乳酸与丙酮酸的可逆反应向生成丙酮酸方向进行,减少乳酸的产生;另外,烟酸还可能促进了瘤胃内乳酸利用菌(埃氏巨型球菌和反刍兽新月单胞菌)的生长,维持瘤胃内环境的平衡,避免乳酸的蓄积。本试验结果与杨艳[11]的研究结果一致。血清中乳酸浓度的变化与瘤胃中乳酸浓度变化呈线性相关,随着瘤胃液中乳酸浓度的升高而升高;此外,血清中L-乳酸、D-乳酸浓度随烟酸添加量的增加呈现降低的趋势。
3.2 高精料饲粮中添加烟酸对绵羊瘤胃液中NAD+、NADH浓度以及乳酸脱氢酶活性的影响
糖酵解的最终产物是丙酮酸。一般情况下,丙酮酸的去路主要有2条:一是丙烯酸途径,另一个是琥珀酸途径[17-19]。丙酮酸在乳酸脱氢酶的作用下还原为乳酸,乳酸在乳酰辅酶A、丙烯酰辅酶A以及丙酰辅酶A的作用下进一步生成丙酸。乳酸脱氢酶活性是评价瘤胃产乳酸能力的重要指标。在丙酮酸转化为乳酸的代谢途径中,NAD+能够抑制乳酸脱氢酶的活性,从而抑制乳酸的生成。在本试验条件下,添加烟酸增加了瘤胃液中NAD+的浓度,降低了NADH的浓度,尤其是试验Ⅱ组,NAD+浓度上升最明显,达到1.38 μmol/L,NADH浓度下降最明显,降至0.90 μmol/L,而试验Ⅱ组瘤胃液中乳酸脱氢酶活性也是最低的,其活性为144.71 U/L,乳酸脱氢酶活性与NAD+浓度密切相关,可能是对照组高精料饲粮消耗了过多的NAD+,乳酸脱氢酶活性急剧上升,乳酸浓度也骤然增加;而在高精料饲粮中添加烟酸后,瘤胃液中NAD+浓度迅速增加,抑制了乳酸脱氢酶的活性,乳酸浓度减少。本试验结果与杨艳[11]的研究结果一致。这一结果刚好验证了本试验上述研究结果,即添加烟酸后瘤胃液和血清中乳酸浓度降低。
3.3 高精料饲粮中添加烟酸对绵羊瘤胃丙酮酸、苹果酸、琥珀酸浓度及其相关酶活性的影响
琥珀酸途径是反刍动物碳水化合物代谢过程中的主要途径之一,丙酮酸在丙酮酸羧化酶、苹果酸脱氢酶、琥珀酸脱氢酶等酶的催化下生成对应的草酰乙酸、苹果酸和琥珀酸,最终生成丙酸[6]。苹果酸是机体代谢过程中产生的重要有机酸,也是三羧酸循环代谢过程中重要的中间产物,促进乳酸利用菌的生长。有研究表明,在生成丙酸的代谢途径中,苹果酸可以弥补草酰乙酸的缺乏,刺激反刍兽新月形单胞菌对乳酸的利用[20]。饲粮中添加苹果酸可显著提高瘤胃液挥发性脂肪酸中丙酸比例,降低乳酸浓度[21-24]。琥珀酸作为琥珀酸途径中的另一中间代谢产物,对改善反刍动物瘤胃发酵有重要作用。有研究表明,在奶牛饲粮中添加琥珀酸,瘤胃液中乙酸和丙酸浓度显著增加,丙酸浓度随琥珀酸添加量的增加呈线性增加[25]。琥珀酸脱氢酶和苹果酸脱氢酶是三羧酸循环的关键酶,其活性反映了有氧代谢能力。在本试验条件下,在高精料饲粮中添加烟酸增加了瘤胃液中丙酮酸、苹果酸和琥珀酸的浓度,提高了瘤胃液中丙酮酸激酶、苹果酸脱氢酶和琥珀酸脱氢酶活性,尤其是试验Ⅱ组。这表明烟酸对绵羊能量代谢有显著影响,能增强有氧代谢能力,降低糖酵解代谢能力;另外,烟酸提供的NAD+会将乳酸持续转化为丙酮酸,进一步增加琥珀酸途径中间代谢产物,继续产生丙酸[26],为机体提供能量,这与Yu等[27]的研究结果相似。
4 结 论
① 在本试验条件下,高精料饲粮中添加烟酸提高了绵羊瘤胃内乳酸代谢途径中丙酮酸、苹果酸以及琥珀酸的浓度,并且降低了瘤胃液和血清中L-乳酸、D-乳酸浓度。
② 高精料饲粮中添加烟酸提高了绵羊瘤胃液中NAD+浓度以及丙酮酸激酶、苹果酸脱氢酶以及琥珀酸脱氢酶活性,降低了NADH浓度以及乳酸脱氢酶活性。
③ 绵羊高精料饲粮中烟酸的推荐添加量为130 mg/d。
猜你喜欢
牡丹江医学院学报(2021年5期)2021-12-05
食品安全导刊(2018年30期)2019-01-28
心肺血管病杂志(2018年11期)2018-12-18
中国医药生物技术(2015年4期)2015-12-26
精细石油化工(2015年3期)2015-12-14
四川师范大学学报(自然科学版)(2015年1期)2015-02-28
现代检验医学杂志(2015年5期)2015-02-06
河南医学研究(2014年3期)2014-02-27
中国饲料(2012年21期)2012-06-29
天津科技(2011年1期)2011-10-13
