
不同树龄咖啡叶片与土壤的C、N、P生态化学计量研究
2023-11-11 15:55陈肖崔现亮罗娅婷赵利坤李学玲
热带作物学报 2023年9期
陈肖 崔现亮 罗娅婷 赵利坤 李学玲
关键词:生态化学计量学;咖啡;叶片;土壤;树龄
中图分类号:S571.2 文献标识码:A
咖啡(Coffea arabica L.)为茜草科咖啡属的多年生经济作物,生长速度快,产量高,价值高,销售渠道广,与茶叶、可可并称为世界三大饮料作物[1]。云南省普洱市气候温和,昼夜温差大,热量条件好,全年无霜期长,雨量适中,海拔在317~3370 m 之间,冬无严寒,夏无酷暑,非常适合咖啡生长[2]。
生态化学计量学特征的研究一般从分子水平、细胞个体水平、群落与生态系统、全球区域尺度几个层次进行。从全球区域尺度来看,土壤的化学计量比能反映内部重要元素的循环,一方面有利于确定生态过程对全球变化的响应,另一方面可以用来判断土壤矿化作用和生物固持作用。植物N、P 作为敏感限制性元素,能反映土壤中N、P 的有效性,对此研究更深入了解全球碳模拟和宏观生态学的变化[3-6]。土壤为植物提供了营养物质,植物叶片的元素大部分来源于土壤,土壤养分的利用决定了植物元素的浓度,叶片中养分含量状况能够较好地反映土壤养分供给的能力[7]。植物和土壤是一体的,植物中的N∶P 可以预测土壤N、P 的养分供应情况[8-9]。AERTS 等[10]和CHAPIN 等[11]的研究也表明N∶P 可作为当前限制性养分判断的指标之一。阈值试验[11]研究表明,当叶片N∶P<14 时,植物生长主要受N 限制;叶片N∶P>16 时,植物生长主要受P 限制;当N∶P 比值为14~16 时,植物生长受N 和P 共同限制[12]。此外,植物的碳同化速率可以通过叶片的C∶N 和C∶P 来反映[13]。
在国内,ZHANG 等[14]最早对生态化学计量学进行了综述,目前热点研究对象主要是生态系统、植物器官、生长阶段和种群。植物在不同生长阶段时,会根据植物对环境的适应将有限的养分资源按照一定比例分配给不同器官,因此植物形成了一定的生长特征和元素的分配规律。在不同生育阶段,C∶N 随着生长呈增加的趋势,一般来说,幼龄的植物叶片含氮量最高,由于植物幼株要生长发育,需要大量的N、P,因此幼龄植株的N∶P 相对较低[15],到成熟期达到最高,造成这种趋势的原因是由于随着植物的衰老,氮代谢从同化向再分配转化,其体内N 的分解速度和释放量相对大于C,所以使得比值下降。而C∶N随着生长呈降低的趋势[16-17]。对植物器官的研究中主要集中在叶片生态化学计量特征、CNP 元素循环、生物养分限制判断、氮磷添加与叶片相关性、C、N、P 生态化学计量学特征对植物生长的指示性作用、叶片的动态特征、植物叶片与土壤的相关性等方面[18-25],其中把气温、水分、海拔、演替阶段作为变量条件来进行研究较多,而对不同树龄乔木之间研究较少,有较少学者如叶柳欣等[26]在研究中表明:杨梅叶片C、N 含量与树龄无关。土壤SOC、TN、TK 含量则随着树龄增长而先降低而后升高,杨梅生长过程中对P 较为敏感。邓成华等[27]和吴家森等[5]研究中表明:油茶叶片C 含量在不同树龄间差异并不显著,油茶生长的限制元素是N。马任甜等[28]在对刺槐的研究中表明:P 相对缺乏,成为刺槐人工林生长的主要限制因子。
综上所述,生态化学计量的研究层次范围较广,在咖啡土壤方面从生态化学计量学方面来研究的还未涉及,鉴于此,本研究通过对不同树龄咖啡叶片和土壤生态化学计量特性的研究,了解咖啡土壤和叶片元素间的相关性、养分循环过程和养分限制关系,判断限制元素,有助于系统揭示咖啡在不同生长阶段植物-土壤养分的分布特征,为咖啡的土壤养分管理提供理论依据,同时为生态化学计量学理论研究提供新思路。
1 材料与方法
1.1 材料
研究地点位于云南省普洱市思茅区南屏镇某咖啡庄园(100°57′28″E,22°39′40″N),面积约20 hm。海拔1070~1200 m,年均气温17.8 ℃,年均降雨量1524.4 mm,无霜期318 d,气候类型多样,降雨充沛、纬度较低、温度较高、湿润。土壤类型主要为砖红壤,受生物积累作用呈灰棕色,土体深厚,质地偏沙,土壤偏酸性,分散性大,絮固作用小,耕作容易,宜种性广。
1.2 方法
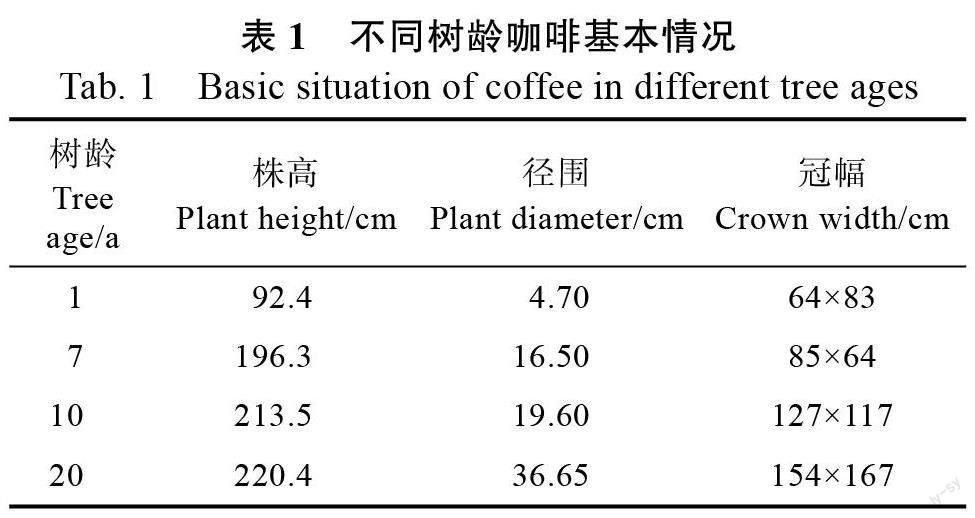
1.2.1 实验设计与采样 研究的咖啡树龄有1、7、10、20 a 四个不同树龄,设4 个标准地,测量标准地内咖啡树的株高、径围和冠幅并计算其平均值(表1)。每个标准地内设10 m×10 m 的样方4 个,共计16 个,在每个样方内根据地形用对角线取样法,进行5 个点的取样,用土钻在每个样点取咖啡树根际集中区域0~10、10~30 cm 两个土层的土样,把相同土层深度同一树龄的土样进行混合,用四分法分取约1 kg,共计32 个土样,密封带回实验室;在土样采集的同一咖啡树上,沿咖啡树东南西北4 个方位,按照树冠外部和内部剪取无病害成熟叶片30 片,共计16 个叶片样品,将叶片放入自封袋中带回实验室。
1.2.2 样品处理 (1)土样处理。将带回实验室的土样除去肉眼可见的砂石、根系、腐烂叶片等雜质,将大块土样碾碎后,于通风的地方平铺风干1 周,过80 目筛后备用。
(2)叶片样品处理。将带回的新鲜叶片先用蒸馏水清洗,然后在105 ℃杀青30 min,最后烘箱80 ℃烘干至恒重,粉碎机粉碎过100 目筛备用。
1.2.3 样品测定[29] 采用重铬酸钾容量法-外加热法测定植物碳(C)和土壤的有机碳(SOC)含量;用k9840 自动凯氏定氮仪进行土壤全氮(TN)的测定;HSO-HO 消煮,奈氏比色法测植物氮(N);HClO-HSO 法测土壤全磷(TP); 用HSO-HO 消煮,钼锑抗比色法测植物中的磷(P)。
1.3 数据处理
所有数据采用SPSS 26 软件进行分析。对咖啡叶片的C、N、P和土壤的SOC、TN、TP和C∶N、C∶P、N∶P 间的差异显著性采用单因素方差分析。咖啡叶片与土壤C、N、P 含量之间的关系采用Pearson 相关性分析判断。数据表示均为平均值±标准差。
2 结果与分析
2.1 不同树龄咖啡叶片C、N、P 含量及化学计量比
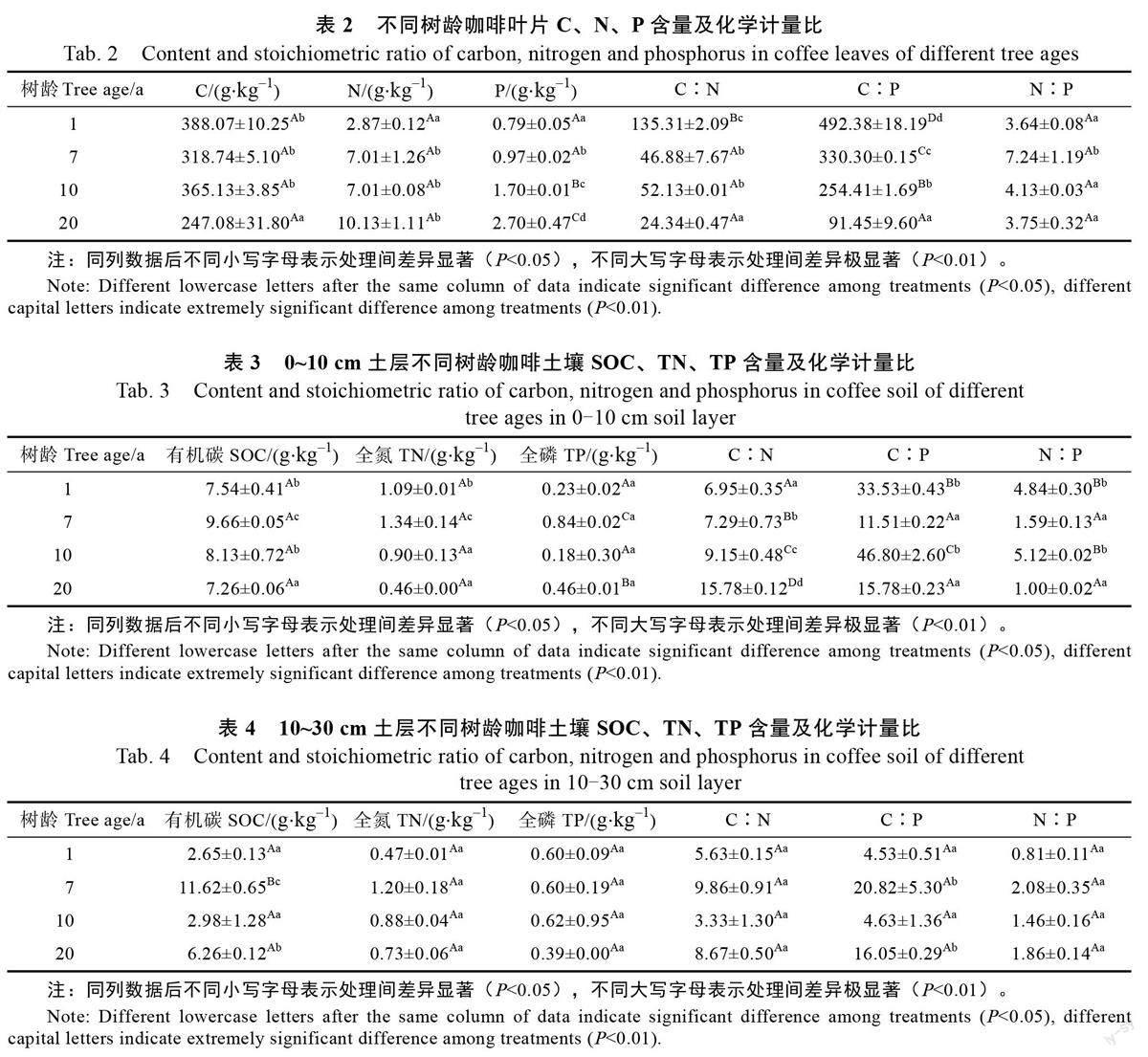
由表2 可知, 叶片C 含量介于247.08~388.07 g/kg 之间,C 含量均值为329.76 g/kg,其中1 a 的咖啡叶片C 含量最高,随树龄段的增加C 含量逐渐减少,20 a 的咖啡叶片C 含量和其余3 个树龄段差异显著(P<0.05);N 含量为2.87~10.13,均值为2.76 g/kg,随树龄的增加,N 含量增加,1 a和其他3 个树龄段达到显著水平(P<0.05),7、10、20 a 的差异不明显;P 含量为0.79~2.70,均值为1.54 g/kg,随着树龄的增加,P 含量显著增加,差异达到显著水平(P<0.05),其中1、7、10、20 a四个树龄差异达到显著水平(P<0.05);C∶N 均值为64.66,在不同树龄段差异较大,7 a 和10 a无显著差异,和1 a、20 a 差异极显著水平(P<0.01),C∶P 均值为282.39,和树龄呈反比,随树龄的增加,比值逐渐减小,不同树龄段差异达到极显著水平(P<0.01);N∶P 均值为4.69,7 a 和其余年龄段差异达到显著水平(P<0.05)。
2.2 不同树龄咖啡土壤SOC、TN、TP 含量及化学计量比
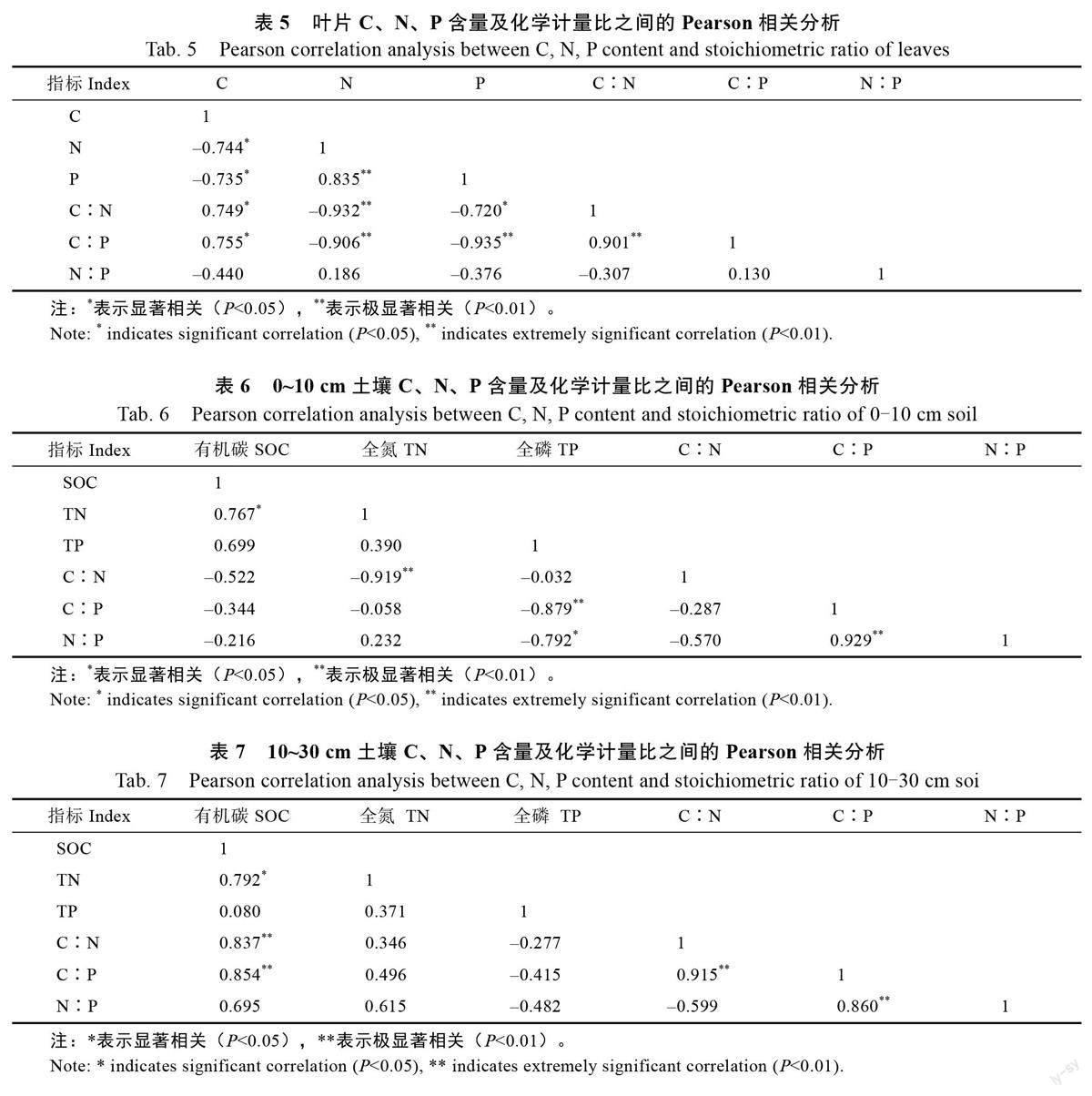
由表3、表4 可知,土层深度0~10 cm 的土壤SOC 含量均值为8.15 g/kg,10~30 cm 土层的均值为5.88 g/kg,由于土壤中的SOC 主要来源于凋落物的归还,首先富集在土表随时间推移下渗,所以0~10 cm 土层的SOC 含量略高于10~30 cm土层,除1 a 和10 a 外,其余差异显著(P<0.05),1 a 的土壤SOC 含量较低;0~10 cm 土层TN 含量均值为0.95 g/kg,10~30 cm 土层为0.82 g/kg,不同土层间无明显差异,1、7、10 a 间差异显著(P<0.05);不同土层间的TP 含量分别为0.43、0.56 g/kg,不同树龄段的0~10 cm 土层TP 含量差异显著(P<0.05),10~30 cm 土层无显著差异;0~10 cm 土层的C∶N 均值为9.79,略高于10~30 cm,其中0~10 cm 土层的不同树龄段存在极显著差异(P<0.01),10~30 cm 土层无显著差异;C∶P 值0~10 cm 高于10~30 cm 土层,差异显著;0~10 cm土层的N∶P高于10~30 cm土层,其中0~10 cm 土层中1 a 和7 a,10 a 和20 a 差异达到极显著水平(P<0.01),10~30 cm 土层无显著差异。
2.3 咖啡葉片C、N、P 含量和土壤SOC、TN、TP 含量和化学计量比之间的关系
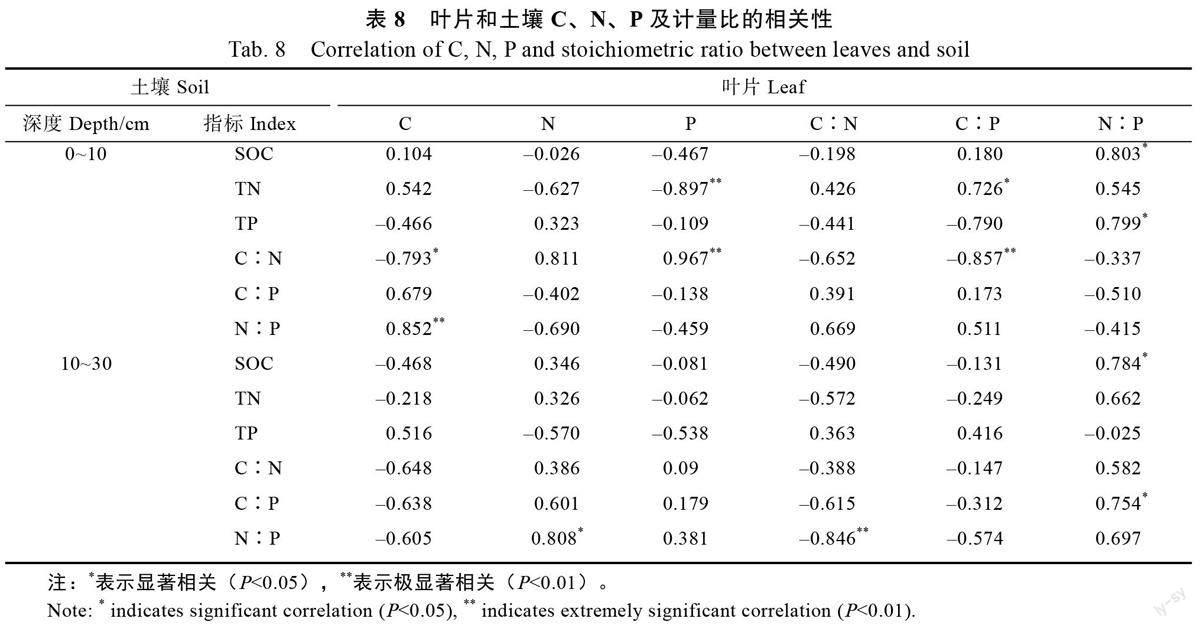
由表5 可知,叶片C 和N、P 呈显著负相关关系(P<0.05),与C∶N、C∶P 呈显著正相关关系(P<0.05);N与P呈极显著正相关关系(P<0.01),与C∶N、C∶P 呈极显著负相关关系(P<0.01);P 和C∶N,C∶P 呈显著负相关关系(P<0.05),与C∶P 呈极显著负相关关系(P<0.01),与C∶N、C∶P 呈正相关关系(P<0.01)。
由表6 可知,0~10 cm 土层的C∶P 与N∶P呈极显著正相关关系(P<0.01)。SOC 与TN 呈显著正相关关系(P<0.05),TN 与C∶N 呈极显著负相关关系(P<0.01),TP 与N∶P 呈显著负相关关系(P<0.05),与C∶P 呈极显著负相关关系(P<0.01);C∶P 与N∶P 呈极显著正相关关系(P<0.01)。
由表7 可知,10~30 cm 土层土壤SOC 与TN呈显著正相关关系(P<0.05),与C∶N、C∶P 呈极显著正相关关系(P<0.01);C∶N 与C∶P,C∶P与N∶P 呈极显著正相关关系(P<0.01)。
由表8 可知,咖啡叶片的C 含量与0~10 cm土层的N∶P 呈极显著正相关关系(r=0.852,P<0.01),与C∶N 呈显著负相关关系(r=‒0.793,P<0.05),与10~30 cm 土层土壤的各项指标无相关性:叶片N 含量与10~30 cm 土层土壤的N∶P呈显著正相关关系(r=0.808,P<0.05);叶片P含量与0~10 cm 土层土壤的TN 呈极显著负相关关系(r=‒0.897,P<0.01),与C∶N 呈极显著正相关关系(r=0.967,P<0.01);叶片C∶N 与10~30cm 土层的N∶P 呈极显著负相关关系(r=‒0.846,P<0.01);叶片C∶P 与0~10 cm 土层土壤的TN呈显著正相关关系(r=0.726,P<0.05),与0~10 cm土层的C∶N 呈极显著负相关关系(r=‒0.857,P<0.01);叶片N∶P与0~10 cm土层土壤的SOC、TP 呈显著正相关关系(r=0.803、r=0.799,P<0.05),与10~30 cm 土层土壤的SOC、C∶P呈显著正相关关系(r=0.784、r=0.754,P<0.05)。
3 讨论
3.1 不同树龄咖啡叶片C、N、P 含量及化学计量比
咖啡叶片C 在247.08~388.07 g/kg 之间,均值329.755 g/kg,低于全球植物叶片C 元素含量462 g/kg,N 含量介于2.87~10.13 g/kg 之间,均值为6.76 g/kg,明显低于全球尺度12.20 mg/g,叶片P 含量在0.79~2.70 g/kg 之间,均值1.54 g/kg,接近于全球尺度(1.8 g/kg)和略低于中国东部南北森林生态样带优势树种叶片平均P 含量(2.0 g/kg)[30],叶片中3 种元素均低的原因与咖啡土壤养分水平偏低有关。
随树龄的增长,咖啡叶片C 含量逐渐减少,因为咖啡成熟期生长基本停滞,光合作用速率降低,吸收的营养元素主要转移到生殖器官,用于种子的发育,故此时C 含量最低。随树龄的增加含N 量增加,叶片N 的含量能反映植物生长速率,叶片含N 量高,光合速率快,生长快,本研究说明咖啡生长速率较低,但随树龄的增加,生长速率加快。咖啡P 含量随树龄的增加,含量显著增加,20 a 的含P 量最高,这是由于生长旺盛期植株生长速率较快,细胞代谢增强,需要合成大量的蛋白质和核酸[11],加上植株生物量增加,富含C 的结构性物质积累,造成N、P 元素含量大幅度上升。
咖啡叶片C∶N 均值为64.66 g,高于全球水平25 g[31],C∶P 均值为282.39,高于全球水平232,与树龄呈负相关,生长速率与体内C∶P 呈负相关,N∶P 为4.13~7.24,均值为4.69,低于全球水平,生长速率理论(growth rate hypothesis,GRH)认为,生长速率的改变引起生物体C、N、P 化学计量改变,高生长速率的植物通常具有较低C∶N、C∶P 和N∶P,从而将P 更多分配到核糖体RNA,满足核糖体迅速合成蛋白质以支持快速生长[6],咖啡叶片C∶N、C∶P 和N∶P 随树龄增大有所降低,表明在低幼龄林阶段,咖啡树生长快,对C、N、P 养分需求高,而在成熟林阶段,咖啡维持相对恒定的慢速生长。大量研究表明,N∶P 明显小于14 时,主要受N 元素影响,其中7 a 的N∶P 最高,说明受N 元素限制最明显。
3.2 不同树龄咖啡土壤SOC、TN、TP 含量及化学计量比
不同树龄咖啡0~10、10~30 cm 土层SOC 含量均值分别为8.15、5.88 g/kg,随土层的增加,SOC 含量减少,是由于SOC 主要来源于凋落物的归还,凋落物首先富集在土层表面,随时间的推移再逐渐淋溶下渗到深层土壤;不同土层的咖啡TN 含量均值分别为0.95、0.82 g/kg,在土层上无明显差异;4 个树龄0~10、10~30 cm 土层TP 含量在0.18~0.84、0.47~1.20 g/kg 之間,不同土层差异不明显,均低于全球水平3.90 g/kg[32],接近我国土壤P 含量平均值0.56 g/kg,这是因为亚热带区红壤风化淋溶作用较强,富含的铝、铁、锰氧化物等矿物对P 素专性吸附和固定能力强,导致研究区P 元素含量少。4 个树龄段中,7 a 的C、N、P 含量均为最高,土壤的C、N、P 含量主要来源于枯枝落叶的归还和根系的周转,7 a 的咖啡树正值生长发育的旺盛期,树木冠幅大,叶片多,代谢快,凋落物多。
C∶N 是土壤质量的敏感指标,土壤中有机C和N 的循环受C∶N 影响,从相关性上看,C、N均和C∶N 具有极显著相关性,土壤C∶N 可以衡量土壤N 素矿化能力,与土壤SOC 分解速率呈负相关。0~10、10~30 cm 的咖啡C∶N 的变化范围分别为6.95~25.78、3.32~9.85,低于中国和世界土壤C∶N 的平均值(11.90 和13.33)[33],不同树龄咖啡的土壤C∶N 差异显著。土层0~10、10~30 cm 的咖啡的C∶P 变化范围分别为11.51~46.80、4.53~20.82,土层间差异较大,但均低于全国平均水平136,也低于全世界的平均值186。0~10 cm 土层咖啡土壤N∶P 变化范围为1.00~4.84,10~30 cm 土层为0.81~2.08,均低于全国土壤N∶P 平均值5.20,土层和树龄对N∶P 均有影响,这可能与咖啡经营过程中,低C 投入,高N、高P 投入,土壤P 素固定有关。咖啡土壤C、N、P 含量及相关性研究表明N∶P 和P 元素有显著相关性,说明P 元素是造成C∶N 变化的主要原因。
3.3 咖啡叶片C、N、P 含量和土壤SOC、TN、TP 含量和化学计量比之间的关系
研究发现叶片C、N、P 及化学计量显著大于土壤SOC、TN、TP 及化学计量,这与邓成华等[27]研究的不同树龄的油茶一致,叶片的C、N、P 与土壤的SOC、TN、TP 无相关性,叶片C∶N 与0~10 cm 土层的TN 呈显著正相关关系,说明土壤TN对植物的C∶N有影响,会直接影响光合效率。相比之下叶片的C、N、P 及计量比更受浅土层0~10 cm 土壤的影响。
猜你喜欢
天天爱科学(2022年12期)2022-11-10
好日子(2022年3期)2022-06-01
装备制造技术(2020年1期)2020-12-25
果树资源学报(2020年3期)2020-05-13
新疆农业科学(2019年11期)2019-12-31
天然产物研究与开发(2016年11期)2016-06-15
Coco薇(2015年12期)2015-12-10
小天使·四年级语数英综合(2015年6期)2015-07-17
航空学报(2015年4期)2015-05-07
食品科学(2013年10期)2013-03-11
