
β-半乳糖苷酶的来源、改造、异源表达及其在食品中的应用
2023-12-03 12:37李云亮谢鹏飞刘晓霜晁嘉品赵晓雪王晓静马海乐
食品工业科技 2023年23期
李云亮,谢鹏飞,刘晓霜,晁嘉品,赵晓雪,王晓静,马海乐,
(1.江苏大学食品与生物工程学院,江苏镇江 212000;2.卫仕宠物营养科学研究院(江苏)有限公司,江苏镇江 212000)
β-半乳糖苷酶(β-galactosidase EC.3.2.1.23),全称为β-D-半乳糖苷半乳糖水解酶,可通过切割乳糖中由β-1-4-糖苷键连接的D-半乳糖残基,使乳糖降解成为可被机体吸收的β-半乳糖和葡萄糖[1]。β-半乳糖苷酶来源丰富[2],广泛存在于哺乳动物、植物和微生物中。在幼年哺乳动物的皮肤、特别是小肠等器官组织中存在丰富的β-半乳糖苷酶[3-4],而成年个体的消化道内β-半乳糖苷酶含量较低,无法很好地分解和利用乳糖;虽然很多食物、植物中均发现了β-半乳糖苷酶,如番茄[5]、水稻[6]、拟南芥[7]、苹果[8]和咖啡豆[9]等,然而植物来源的β-半乳糖苷酶在组织中含量并不高,因此提取得率较低[10];微生物中β-半乳糖苷酶资源很丰富,大肠杆菌、酵母菌、霉菌等细菌、真菌均能产β-半乳糖苷酶。
目前商业化使用的β-半乳糖苷酶主要采用大肠杆菌、黑曲霉等微生物发酵法制备,但存在酶活不高、稳定性较差等问题,且很多微生物发酵产物多以胞内酶的形式存在,导致酶纯化工艺复杂、成本增加。为解决此类问题,研究学者对β-半乳糖苷酶基因进行异源表达,或者将其改造后再进行异源表达,即将β-半乳糖苷酶基因转入至外源宿主菌[11]中,利用宿主细胞自身代谢资源(能量分子、氨基酸等)来实现表达[12],展现了良好的应用前景。基于此,本文对β-半乳糖苷酶的应用、β-半乳糖苷酶基因的来源及其异源表达、表达宿主和表达方式展开综合论述,为β-半乳糖苷酶的开发与改良提供科学依据与理论参考。
1 β-半乳糖苷酶的来源及改造
1.1 β-半乳糖苷酶来源
目前NCBI 中登记的β-半乳糖苷酶基因已有1.7 万余个,图1 中显示真核生物共12188 个、细菌4393 个、古生菌415 个以及病毒19 个(截止2023.04.06)。同一家族来源的β-半乳糖苷酶具有相似的催化机制,不同家族来源的β-半乳糖苷酶则在酶学性质上有着较大差异,表1 列出了较为常见的不同来源的β-半乳糖苷酶特性。贺璐等[13]发现细菌来源的β-半乳糖苷酶多为常温酶,如大肠杆菌和保加利亚乳杆菌来源的β-半乳糖苷酶最适反应温度一般在40 ℃左右,最适作用pH 在6.5~7.5 之间。酵母菌来源的β-半乳糖苷酶具有较强的水解活性[14],常用于牛乳和乳清中乳糖的水解。霉菌来源的β-半乳糖苷酶最适反应温度通常在50 ℃以上,最适pH 偏酸性,其中黑曲酶[15]来源的β-半乳糖苷酶耐热耐酸性较强,可用于干酪、牛乳和酸性乳清的水解处理,而米曲霉[16]来源的β-半乳糖苷酶则具有较强的转糖基活性,可用于生产低聚半乳糖。
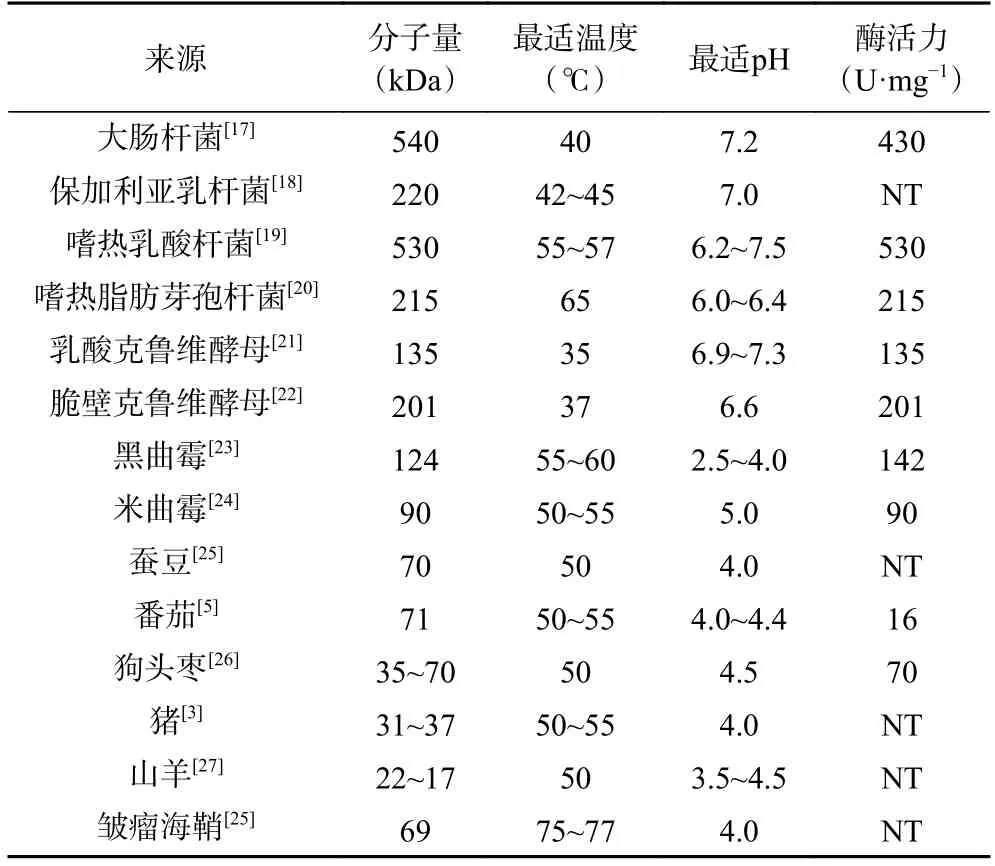
表1 不同来源β-半乳糖苷酶特性Table 1 Characteristics of β-galactosidase from different sources
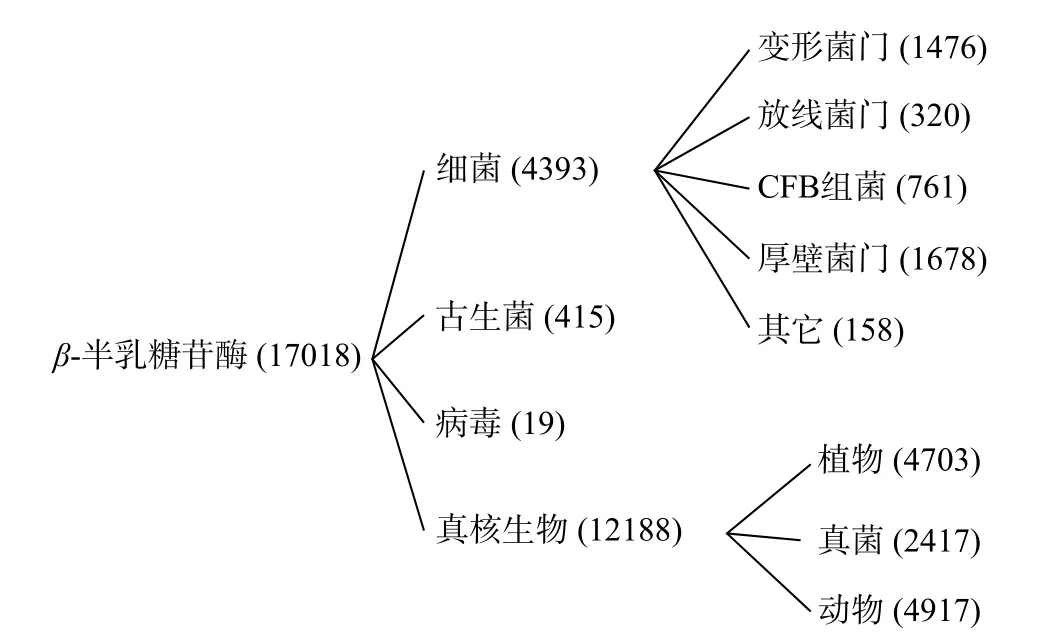
图1 已登记β-半乳糖苷酶基因来源(截止2023.04.06)Fig.1 Registered gene sources of β-galactosidase(until 2023.04.06)
1.2 β-半乳糖苷酶改造
天然微生物以及动植物中的β-半乳糖苷酶存在热稳定性差、酶活低等问题,往往不能很好地满足目前工业发展需求,可通过分子进化手段对β-半乳糖苷酶基因进行定向改造,将其在体外模拟随机突变[28]、重组,使得目的基因发生大量变异,筛选获得具有特定性质的目的基因。彭慧等[29]将来源于T.scotoductus的野生型β-半乳糖苷酶GaLT0 的N 端氨基酸序列插入一个经过人工改造的双亲短肽序列(AEAEAKAKAEAEAKAKAEAEAKAK),经改造后的β-半乳糖苷酶,稳定性更高,55 ℃环境下半衰期提高了10 倍,2 h 内可水解牛奶中95%的乳糖。杨萍[30]通过将来源于米曲酶的乳糖酶基因作为出发点,通过PCR 引物设计,对基因序列中第211 位氨基酸进行改造,将Thr 突变为Asn、Asp、Ser、Pro 和Gly,发现当第211 位氨基酸突变为Pro 后,在60 ℃环境下的半衰期相对于野生型的2.06 min 提高到177.73 min,极大地改良了乳糖酶的热稳定性。对β-半乳糖苷酶基因的改造不仅局限于使其具有更广泛的环境适应范围,还可以增加自身表达水平、提高酶活力。李晨霞等[31]通过易错PCR 技术构建了米曲霉β-半乳糖苷酶(AoBgal35A)的突变体文库[32]并对其进行分析,发现AoBgal35A 中第955 位的苏氨酸(Thr)突变为丙氨酸(Ala),使得极性残基突变为非极性残基,最终获得的β-半乳糖苷酶的最适pH 提高了0.5,酶活达4760 U/mL,是目前米曲酶来源的β-半乳糖苷酶基因在毕赤酵母中获得的最高表达水平。目前随着研究的不断深入,日趋成熟和完善的多种新型分子进化技术如DNA 改组[33]、易错PCR[34]、交错延伸等方法将有望成为β-半乳糖苷酶基因改造的有效工具。
2 β-半乳糖苷酶的异源表达系统
选择合适的表达系统是使β-半乳糖苷酶基因得以高效表达的关键因素,目前β-半乳糖苷酶异源表达的系统主要分为原核生物和真核生物表达系统,表2 列出了目前常用的表达系统及其特点。

表2 常见表达系统特点Table 2 Characteristics of common expression systems
2.1 原核表达系统
原核表达系统中最常见的是大肠杆菌表达系统[40],大肠杆菌生长速度极快,具有连续发酵的能力,以及表达量高、生产成本低[41]、遗传背景清晰[42]等特点,利用其作为表达宿主表达β-半乳糖苷酶基因,技术路线成熟、可行性高[43]。陈卫等[44]将来源于嗜热脂肪芽孢杆菌的β-半乳糖苷酶基因连接至大肠杆菌表达载体pET-20b 中,转化至大肠杆菌BL21 中诱导表达的重组蛋白比酶活为6.66 U/mg 蛋白,相比于出发菌株提高了50 倍。徐顺清等[45]将来源于乳酸克鲁维酵母的乳糖酶基因克隆至大肠杆菌表达载体pET-28a(+)上,转化至大肠杆菌BL21 后28 ℃诱导4.5 h,测得乳糖酶酶活为44.78 U/mL 发酵液,但在添加Ca2+、Zn2+、Fe2、CO2+、Mg2+和Mn2+金属离子后,酶活分别提高了18.53%、54.49%、293.65%、347.56%、422.95%和549.81%,而Cu2+的添加使得酶活完全被抑制,这为之后完善β-半乳糖苷酶发酵体系提供了理论依据。利用大肠杆菌表达系统生产β-半乳糖苷酶时,若稀有密码子连续出现,则会一直蛋白质的合成,发生密码子错配等现象,所以可在基因的设计前进行密码子的优化,如非连续性多核苷酸定点突变法对cDNA 中稀有密码子进行定点突变,或提高某种氨基酸的tRNA 浓度等。此外,蛋白在表达过程中会存在内毒素等有害物质残留风险,限制了其在食品领域的应用,所以食品级表达系统的建立对β-半乳糖苷酶的生产也是十分重要的。
乳酸菌,如乳酸乳球菌、乳杆菌和嗜热链球菌等属于安全的益生菌[46],利用其作为表达的宿主菌,具有较高的安全性,近些年来该表达系统正成为研究的热点。孙芝兰等[47]将Paenibacillussp.K1 乳糖酶基因构建至载体pSEC 和Pmg36e 上,电击转化至乳酸乳球菌(Lactococcus lactisMG1614)中,其表达后获得的β-半乳糖苷酶对1%的乳糖进行发酵,乳糖残余量为0.56%,水解率增加一倍。由于乳酸乳球菌具有较高的食品安全特性,可使用其系统进行食品工业中相关蛋白酶制剂的开发。乳酸菌表达系统大多都是胞内表达,自身也存在分泌困难[48]等问题,而芽孢杆菌表达系统与乳酸菌表达系统相比,具备较强的分泌高活性蛋白的能力[49]。许俊勇等[50]首次将来源于环状芽孢杆菌(Bacillus circulans)的β-半乳糖苷酶基因在枯草芽孢中进行异源表达,经过3 L 罐的高密度培养,最终获得的酶活为138.29 U/mL,是普通摇瓶发酵的20.3 倍。
利用上述原核表达系统表达的β-半乳糖苷酶以游离酶的形式表达时,可采用酶固定化的方法增强其稳定性和重复使用性,如包埋法[51]、吸附法[52]等,但这些方法存在操作过程复杂、成本较高的缺点。为了简化工艺,酶自固定化法应运而生,自固定化指的是酶(或融合酶)在特定条件下,自发形成不溶状态的过程,可采取将含有水不溶性的融合标签基因和可溶性酶基因组合在一起,使酶以不溶性包涵体的形式表达。王晓静[53]以李云亮[54]构建的金枪鱼多肽TunaAI 32 聚体基因为基础设计出了两种水不溶性的融合标签基因,并与乳糖酶基因进行融合表达,该自固定化乳糖酶的热稳定性比市售乳糖酶更强,在60 ℃保温1 h 后相对酶活仍达到57.35%,其在低温环境中储藏10 d,仍可保持90%以上的酶活,展示出了良好的应用前景。因此自固定化融合标签开发和设计可成为今后科研人员的研究重点之一。
2.2 真核表达系统
真核表达系统相对于原核表达系统具有翻译后的加工修饰功能[49],内部具有严格调控基因[55]的能力,表达的外源蛋白更接近于天然蛋白质,利用这一特点,真核表达系统常被用于具有复杂结构蛋白质的表达,其中以酵母表达系统最为常见[56]。王敏等[57]将来源于米曲酶β-半乳糖苷酶的基因片段转化至乳酸克鲁维酵母GG799 中,在连续发酵120 h 后,获得的蛋白酶活为120.65 U/mL,由于Pklac1 载体含有真菌乙酰胺酶基因,发酵过程中不用添加任何抗生素,获得的β-半乳糖苷酶经过超滤即可获得较纯酶液,简化了后期处理过程。而酿酒酵母作为表达宿主也适合用于酿酒工业和面包等食品的生产加工过程中。酵母表达系统虽增加了蛋白胞外分泌的可能性,使得蛋白纯化过程相对于胞内表达系统更加容易,但酵母表达系统也存在着分泌蛋白量少,发酵密度低的问题。
而在哺乳动物表达系统中,可采用哺乳动物细胞作为表达宿主,通过其表达的外源蛋白接近于天然蛋白质,可用于重组药物的生产中,但在表达过程中,外源蛋白的生物学活性与宿主免疫原性有时也会产生差别,导致蛋白不能持久稳定表达。此外,此系统所用的表达宿主价格较为昂贵,不太符合大规模对于β-半乳糖苷酶的工业化生产中。对于真菌表达系统,由于表达自身半乳糖苷酶系(α-型和β-型)的能力比较强,利用该类型宿主表达经改造后β-半乳糖苷酶具有很大的价值,但目前相关研究较少,未来有必要对真菌表达系统开展深入研究,为其工业化应用打下基础。
3 β-半乳糖苷酶在食品中的应用
根据我国食品添加剂使用卫生标准GB 1886.174-2016[58],黑曲酶来源的β-半乳糖苷酶可以作为食品添加剂进行使用。β-半乳糖苷酶作为安全性较高的生物酶制剂,其水解乳糖活性及转糖基活性使其在食品工业等领域大有作为。
3.1 缓解乳糖不耐症问题
乳糖不耐症是由负责消化乳糖的肠道β-半乳糖苷酶的活性减少或丧失引起的一种疾病,其以乳糖吸收不良为特征,可能导致腹鸣、腹痛、腹泻和肠胃胀气等临床症状,在哺乳动物中普遍发生于羔羊、仔猫和仔犬等各种幼仔中。据统计在人体内β-半乳糖苷酶的活性随年龄的增长而下降,少数人完全乳糖不耐受,成年人体内β-半乳糖苷酶的活性仅为正常婴幼儿的5%~10%[59]。目前最有效的方法是在乳制品生产过程中添加β-半乳糖苷酶以分解乳糖,制备低乳糖或者无乳糖的牛奶产品[60]。刘杰等[61]在牛乳中添加0.65%的β-半乳糖苷酶反应2 h 可水解72.48%的乳糖,并与益生菌微胶囊混合制备了新型低乳糖益生菌乳粉。我国目前市面上的低乳糖奶制品[62]有零乳糖牛奶、无乳糖舒化奶等,都是通过在生牛乳中添加β-半乳糖苷酶的方式降低乳糖含量。
3.2 生产低聚半乳糖
低聚半乳糖(galactooligosaccharides,GOS)作为一种益生因子调理肠胃功能、可被肠道内青春双歧杆菌[63]、两岐双歧杆菌[64]、婴儿双歧杆菌[65]、长双歧杆菌[66]、短双歧杆菌[67]、干酪乳杆菌[68]、嗜酸乳杆菌[69]、唾液乳杆菌[70]等八种益生菌所利用,抑制有害菌的生长、维持肠道菌群生态平衡[71]、减少肠易激综合症的发生[72],起到提高人体免疫力的作用[73],极具市场开发价值。GOS 主流制备方法是酶法,即利用乳糖为原料,经β-半乳糖苷酶催化水解及转半乳糖苷的作用,从而形成了由葡萄糖与乳糖组成的混合杂质低聚糖[74]。吴昊[75]研究了β-半乳糖苷酶在Mg2+作用下,GOS 的生产率为31.37%远高于对照组,同时将GOS 与抹茶粉进行复合,也解决了抹茶饮料沉淀的问题。据研究表明GOS 不仅作为肠道菌群的益生元,也可作为食品添加剂使用,改善食品风味,提高产品质地,具有广阔的市场应用前景[76]。
3.3 改良乳制品
在乳制品的加工过程中,由于乳糖溶解度较低,在浓缩乳制品或者冷冻乳制品时,一部分钙盐和乳糖结合导致蛋白质沉淀、乳糖结晶,使得产品口感下降[77],故而在生产加工过程中添加β-半乳糖苷酶水解乳糖,可以避免由乳糖引起的蛋白质聚集现象。另外,在酸乳生产加工中,因发酵周期长,李兴[78]发现添加β-半乳糖苷酶可使酸乳凝固时间减少15%~20%,有效缩短发酵周期,且蔗糖用量和产品口感都有所改善[79];在乳清的加工中,因乳清含有约4%左右乳糖,添加β-半乳糖苷酶后可产生乳清糖浆[2],乳清糖浆甜度类似于蔗糖,但溶解度较乳糖提升了数倍,可作为蔗糖的替代品被用于乳清酒、乳清饮料的生产中[80],极大地改善了产品的风味品质,同时拓宽乳清的使用范围。
4 结语与展望
β-半乳糖苷酶在缓解乳糖不耐受症、生产GOS和改良乳制品等领域具有重要的应用价值。可将动植物,尤其是微生物来源的β-半乳糖苷酶基因进行修饰后转入至大肠杆菌BL21、酿酒酵母等表达宿主菌中,通过原核或真核表达系统进行异源表达,相对于传统发酵法,此方式可有效提高其产量与酶活,降低生产成本。但是目前主流的真核或原核表达系统依然存在纯化成本高、安全性偏低等问题,今后除了在现有的异源表达系统基础上开展大量、深度的研究外,还需要对兼具安全性和高分泌表达能力的表达系统进行探索;此外,如何开发新型载体和自固定化融合标签基因,以简化β-半乳糖苷酶固定化的生产工艺并提高产品质量,也应是相关科研人员努力探索的方向之一。加大对β-半乳糖苷酶的异源表达系统的研究力度,不仅对传统食品科学工程领域的发展具有重要意义,更有助于拓展β-半乳糖苷酶在医疗、基因诊断治疗和环境监测等多个领域的应用范畴。
猜你喜欢
海洋通报(2022年4期)2022-10-10
农业工程学报(2022年5期)2022-06-22
中华戏曲(2020年2期)2020-02-12
食品与机械(2017年4期)2017-07-05
食品与生物技术学报(2017年5期)2017-04-08
华东理工大学学报(自然科学版)(2015年3期)2015-11-07
饲料博览(2014年7期)2014-04-05
食品工业科技(2014年15期)2014-03-11
食品工业科技(2014年9期)2014-03-11
中国质量与标准导报(2013年8期)2013-03-11
