
高原鼠兔时空动态的元胞自动机模拟
2010-06-12 01:35刘汉武周华坤
植物保护 2010年6期
刘汉武, 周 立, 刘 伟, 周华坤
(1.运城学院应用数学系,运城 044000; 2.中国科学院西北高原生物研究所,西宁 810001)
高原鼠兔(Ochotona curzoniae)(以下简称鼠兔)是青藏高原的生态关键种。它是小型食肉动物的主要食物,它构建的洞穴为小型鸟类和蜥蜴提供巢穴,它的挖掘活动有助于营养物质的循环并促进生态系统的演变,它还是一种新开发的实验动物[1]。鼠兔的存在是高寒草甸(以下简称草甸)生态系统健康发展和保护生物多样性的重要条件。近年来过度放牧和全球气候变化导致草甸严重退化,这为鼠兔提供了良好的栖息环境,导致其数量激增[2]。鼠兔和放牧动物争夺食物;它的挖掘活动破坏土壤结构,造成水土流失,导致草场进一步退化;作为鼠疫的疫源动物和包囊虫的中间宿主,也频繁地给人类带来麻烦。正由于鼠兔的双重作用,应该加强对其种群动态的监控和管理,使其保持适当的种群规模和空间分布,既发挥其关键种的作用,同时又不破坏草甸。这要求研究鼠兔种群的空间动态规律。
元胞自动机方法常被用来模拟种群的空间动态。元胞自动机是一个时间、空间和状态都离散的动力系统,散布在规则格网中的每一元胞依确定的局部规则作同步更新,大量元胞通过简单的相互作用而构成动态系统的演化。关于元胞自动机的详细论述可参见文献[3-4]。Karafyllidis[3]利用元胞自动机研究了温室效应对昆虫和微生物地理分布的影响,Wang等[4]利用元胞自动机研究了在可控系统中一年生杂草的扩散机理,Matsinos等[5]利用元胞自动机研究了竞争、扩散、干扰对草地植物群落的影响。
本文基于鼠兔和草甸的生态学特征建立元胞自动机模型,来探究鼠兔的扩散机理,更有力地指导鼠兔数量的监测与控制。
1 材料与方法
1.1 元胞自动机模型的建立
影响鼠兔分布的因素很多,如生境位置、土壤质地、离水源的距离、灌丛植物盖度、阔叶植物高度[6]、天敌分布[7]、人类活动[8]等。造成鼠兔扩散的因素有环境变化、捕食压力[7]、分窝等。影响出生死亡的因素有种群密度与环境容纳量的比值、食物条件等。其中有些因素并不是独立的,为简单起见,这里只考虑植被高度、鼠兔密度与环境容纳量的比值。由于特殊的地理和气象条件,草甸有生长季节(4-9月)和非生长季节(10月-次年3月),鼠兔有繁殖季节(4-8月)和非繁殖季节(9月-次年 3月)。将870 m×870 m均质草甸分成17.4 m×17.4 m[9]的小方格,每一个方格是一个元胞,共有50×50个元胞。采用8邻居法,时间步长为1个月,使用周期边界。
0.25 m×0.25 m面积上植被生物量干重x(g)在生长季节(4-9月)满足 Logistic方程[9]x=;在非生长季节(10月-次年3月)满足 Malthus方程[9]x=93.16exp(-0.21t)。植被高度h(m)正比于植被生物量 x,为 h=0.042x[9]。
过低的植被不能为较多的鼠兔提供食物,但随着植被高度的增加,由于视觉受限,鼠兔不能有效发现捕食者,因此鼠兔不选择植被过高的生境生活,所以元胞(i,j)内在时刻t的环境容纳量Kti,j(有效洞口数)先随植被增高而增大,然后减小,直至为零。这里j为[9]
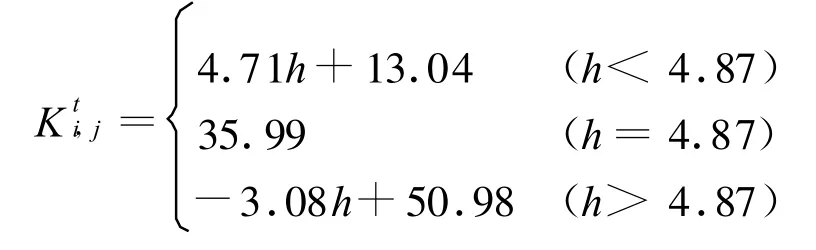
有效洞口的动态表现为在元胞内的生死和元胞间的扩散。在时刻t元胞(i,j)内的有效洞口数在繁殖季节(4-8月)满足 Logistic方程,在非繁殖季节(9月-次年3月)满足Malthus方程
每只鼠兔在植被生长季节日消耗干物质2.4 g,在植被的非生长季节,日消耗23.52 g[10]。鼠兔造成危害的临界阈值为9.05只/hm2[11]。根据刘季科等[11]的数据可折算出鼠兔的洞口系数为0.4,所以在每个元胞内造成危害的经济阈值为有效洞口0.69个,并且,每个有效洞口在植被生长季节日消耗干物质0.96 g,在非生长季节日消耗9.41 g。
1.2 元胞自动机模型的检验
2006年,在青海省果洛藏族自治州玛沁县进行了鼠兔的扩散试验。选择180 m×180 m样地,分成81个20 m×20 m的样方(图1),灰色区域为源区,共4×9个样方,此区域有广阔的背后区域,且源区及背后区域有大量鼠兔,白色区域为扩散区,共5×9个样方。试验开始时统计源区各样方中有效洞口数,灭杀扩散区的全部鼠兔,随后每月记录各样方内有效洞口数。扩散区外是高的人工垂穗披碱草草地,那里没有鼠兔,这样可以保证扩散区内的鼠兔都是从源区迁入或由迁入的鼠兔生产的。
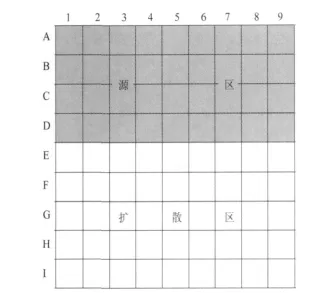
图1 扩散试验样地
由于在元胞自动机模型中元胞边长为17.4 m,而扩散试验样方的边长为20 m,所以在检验模型时将模型中元胞边长调整为20 m,其他参数成比例调整,且将模型中元胞数调整为5×5个。取样方(3~7)×(C~G)中4月份有效洞口数作为初始值代入模型进行模拟,将模拟值和实测值绘于图2,从图中可以看出实测值和模拟值很接近,所以可以认为模型是合理的。
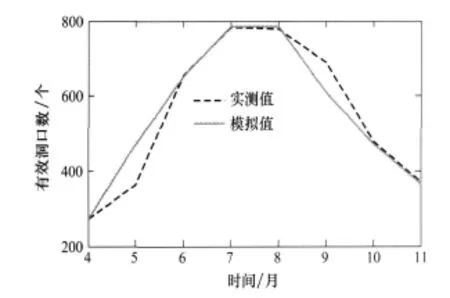
图2 鼠兔种群规模的模拟值和观察值
1.3 模拟
模拟时取r4=-0.26,这样当4月份每个元胞中有5个有效洞口时有效洞口数年复一年地接近周期变化。模拟时取α为0.1。
1.3.1 对扩散的模拟
定义扩散力S为所有元胞与所有邻居间大于α的C值差的和,即元胞(i,j)与它的8个邻居都有一个C的差值,所有大于α的差值的和即为si,j。考虑图3所示的两种初始配置,其中每个黑色元胞内有5个有效洞口,其余的元胞内没有有效洞口,两种配置中有效洞口数相同,都是180个,但集中的程度不同。
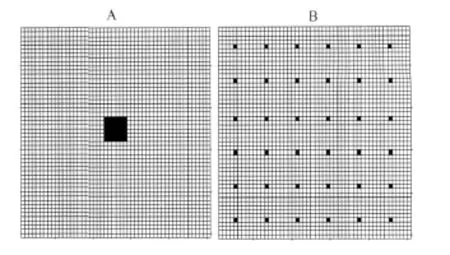
图3 有效洞口的初始配置
1.3.2 模拟鼠兔对恢复草甸的入侵
退化草甸的治理和恢复往往是局部的,恢复的草甸和退化草甸之间有漫长的边界线。在治理刚刚结束时,恢复的草甸内没有鼠兔,而其相邻的退化草甸内有鼠兔,这样退化草甸内的鼠兔就会向恢复的草甸入侵。
模拟时不区分具体的治理措施,只看治理后植被的高度。考虑A 、B、C、D、E、F共 6种情况,即模拟开始时植被高度及其环境容纳量都分别增加0、0.05、0.1、0.15、0.2、0.25 m,其中 A 表示退化的草甸,其他情况下植被逐渐增高。在退化草甸上,鼠兔的数量分7个梯度,即在每个元胞内,分别有0.4、1 、2、3 、4、5 、6 个有效洞口 。
1.3.3 模拟全球变暖对鼠兔的影响
草甸生态系统极其脆弱,对全球气候变化极其敏感,对全球变暖的响应具有超前性。有研究表明全球变暖使草甸生长期延长,生产力下降[12],同时,也有研究表明模拟增温可使植被生长期缩短、生物量减少[13]。在模拟时,植被情况考虑两个方面,共9种情况,即植被生长期延长、不变、缩短;植被生物量增加、不变、减少。
鼠兔对气温升高的反应还未见报道,这里就表1中6种假设分别模拟,当出生率、死亡率改变时,先将出生率调整为原来的95%或105%,然后适当调整死亡率,使得当初始的4月每个元胞内有5个有效洞口时鼠兔种群规模年复一年的近似周期变化,括号内为模拟时使用的出生率和死亡率。
有效洞口的初始配置采用图3B,有鼠兔栖息的元胞内分别有0.4个和5个有效洞口。植被考虑两种情况:退化草甸和非退化草甸。
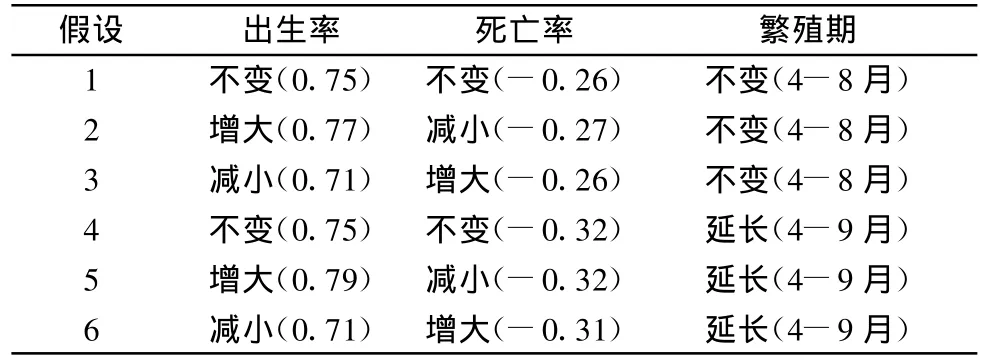
表1 鼠兔种群对气温升高所做反应的6种假设
2 结果与分析
2.1 鼠兔扩散的规律
有效洞口扩散的数量与扩散力的变化趋势基本一致,图4是配置B的情形。当两个元胞间C值的差大于α时,这个差就是扩散力的一部分;同时这个差也导致这两个元胞间有有效洞口的扩散,扩散的数量与这个差有关系,也与当时相邻元胞内有效洞口的数量、环境容纳量及α有关系,所以有效洞口的扩散数量与扩散力之间有类似的变化趋势,但不完全同步。
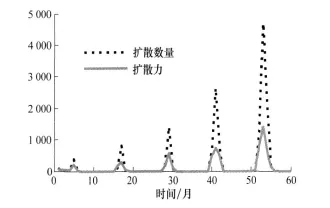
图4 配置B下有效洞口扩散数量与扩散力
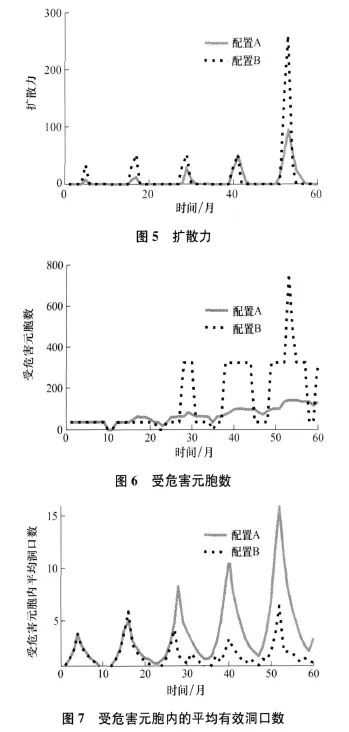
扩散力为所有元胞与所有邻居间大于α的C值差的和,所以扩散力的大小决定于元胞间C值的差和这些差当中有多少个比α大。形象地说,扩散力的大小取决于在多少元胞中要求迁出、有多少元胞可以迁入和迁移的动力有多大。这样与配置A相比,配置B的扩散力要大,图5为含有鼠兔的元胞内有0.4个有效洞口的情形。
两种配置下,鼠兔对草甸的危害如图6所示,鼠兔对草甸总的危害为曲线与横轴之间所夹的面积。图中A、B两个配置下有有效洞口的元胞都含有0.4个有效洞口。从第15到第27个月,配置B的情况危害比较小,这是由于鼠兔通过迅速扩散,占有大量元胞,每个元胞内的有效洞口数都比较少,从而受危害的元胞数也比较少,这可以称为扩散的稀释作用,在配置B的情况下稀释作用较大。从第28个月开始大体上配置B的情况危害比较大,这是由于鼠兔扩散后,虽然每个元胞内的有效洞口数量相对减少,但仍达到危害水平,从而受危害的元胞数比较多,这种情况下稀释作用表现在受危害的程度小。图7为在图6的情况下受危害元胞内有效洞口数的平均值,可以看出自第28个月以后配置B的被危害元胞内的有效洞口要少。
量α对鼠兔的扩散有着重要影响,图8说明了在配置B情况下α对鼠兔扩散的影响。图中曲线分别表示α的值从0逐渐增大时,30个月内被栖息元胞数、扩散力、受危害元胞数的变化过程。图a和b是配置B中每一被栖息元胞内分别有0.4个和5个有效洞口的情形。随α从0逐渐增大,被栖息元胞数从很大迅速减小并逐渐趋于平稳,这是由于α较小时鼠兔可以迅速扩散占领大量元胞。当α变化时扩散力表现出没有规律的震荡,这是因为扩散力不只决定于α,还和各元胞内的C值有关。α变化时受危害元胞数表现出两种不太一样的情形如果开始时被栖息的元胞内有较少的有效洞口,α较小时鼠兔可以通过扩散占有大量元胞,使得每个元胞内的鼠兔都很少,受危害的元胞也就较少;反之,如果开始时被栖息的元胞内有较多的有效洞口,α较小时鼠兔可以通过扩散占有大量元胞,使得每个元胞内的鼠兔相对较少但仍能达到危害水平,这样受危害的元胞就比较多了。
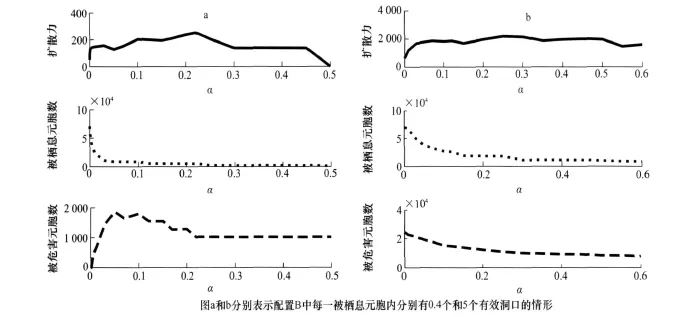
图8 α对扩散的影响
2.2 鼠兔对恢复草甸的入侵
当退化草甸上鼠兔密度比较大时,在相同的时间内入侵恢复草地的距离就比较远。表2为情况C时,鼠兔入侵距离与退化草甸上每个元胞内有效洞口数的关系。
植被比较低时(情况 A、B、C),植被越高,在相同的时间内入侵距离越大。图9为情况A、B、C时4年内的入侵距离。当植被比较高时,治理后的草地上C值与α的关系受多种因素的影响,入侵距离没有明显的规律。
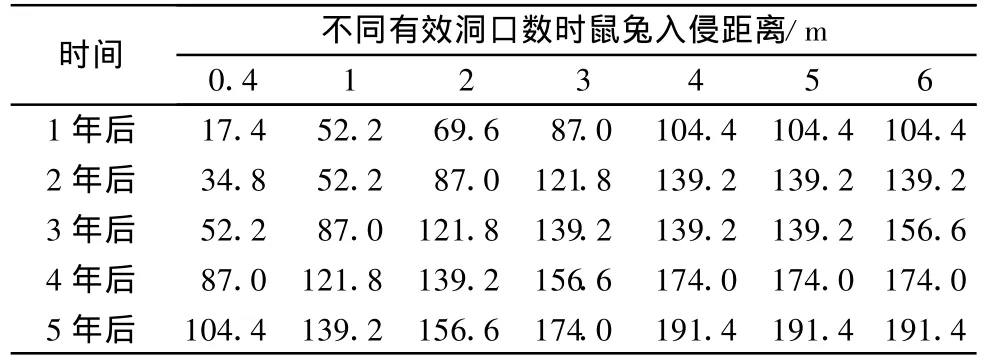
表2 情况C时元胞内不同有效洞口数条件下鼠兔的入侵距离
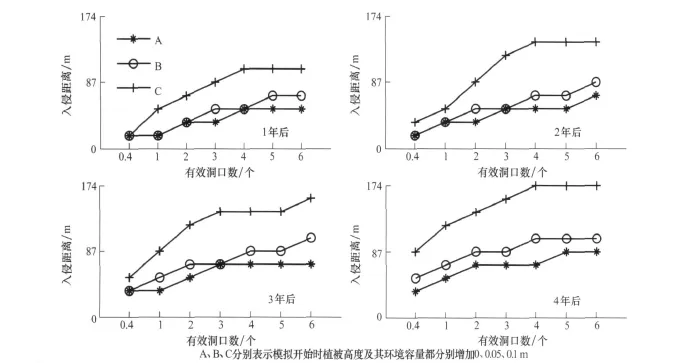
图9 鼠兔的入侵距离
治理后植被低矮时,环境容纳量较大,导致扩散量较大,所以往往鼠兔入侵到的地方危害也随之发生。扩散之后,鼠兔新占据的地方与无鼠兔的地方的C值差较小,在短时间内不再扩散,要等到新占据的元胞内种群规模积累到一定程度才继续扩散,所以入侵的距离一般较短。与之相反,治理后植被较高时,往往鼠兔入侵到的地方危害并不发生,入侵距离一般较长。图10为退化草甸上每个元胞内有3个有效洞口时的入侵过程。
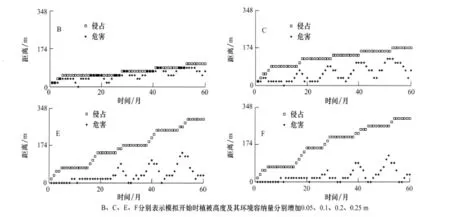
图10 鼠兔的入侵过程
2.3 全球变暖对鼠兔的可能影响
无论植被是9种情况中的哪一种,鼠兔种群都表现出相似的变化方式,所以下面只就植被的生长期延长、生物量下降情况给出模拟结果。在模拟时,生长季节由4-9月调整为4-10月,环境容纳量由0.152 m调整为0.144 m,生长率由0.93调整为0.9,枯死率由-0.21调整为-0.26。此时,在0.25 m×0.25 m面积内,增温后植被的最大干重14.06 g是增温前最大干重14.51 g的97%。
如果鼠兔的繁殖期不延长(假设1、2、3),增温前后,在种群规模、生死数量、扩散数量及危害程度上几乎没有区别。
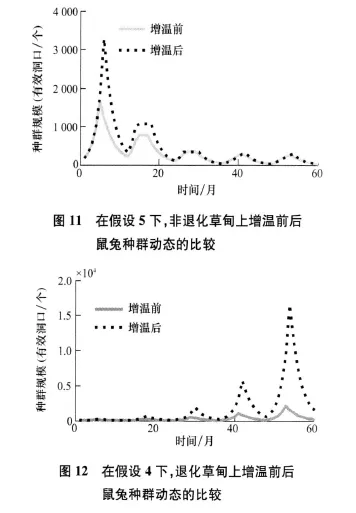
如果增温后鼠兔的生长期延长(假设4、5、6),在退化草甸和非退化草甸上表现出两种不同的情形。在非退化草甸上,尽管增温后鼠兔的生长期延长了,增温前后在鼠兔种群规模、生死数量、扩散数量及危害程度上还是几乎没有区别。图11为在假设5的情况下,非退化草甸上初始时刻含鼠兔的元胞内有5个有效洞口时增温前后的比较。在退化草甸上,鼠兔的环境容纳量较大,加上增温后生长期的延长,使得鼠兔种群迅速增大,同时使得生死数量、扩散数量及危害程度都变大。图12为在假设4的情况下,退化草甸上初始时刻含鼠兔的元胞内有0.4个有效洞口时增温前后的比较。
3 讨论
扩散可减轻鼠兔对草甸的危害,但应该注意危害的减轻是暂时的,鼠兔扩散的最终结果是找到更适合的栖息地,随之而来的是种群以更快的速度增长,并形成更大的种群,产生更大的危害。如果恢复后的植被较高,邻近的鼠兔对其入侵距离远、危害轻,如果恢复后的植被较低,则相反。其原因在于较高的植被不适合鼠兔生存,所以恢复植被,造成不适合鼠兔生存的环境是根治鼠兔危害的最终办法。全球气候变暖已是一个不争的事实,鼠兔,这种接近r对策的动物,没有理由不延长其繁殖期。这样,在退化草甸上,鼠兔种群将迅速增大,使草甸进一步退化,产生更适合鼠兔生存的环境,这种恶性循环会周而复始永无止境地进行下去。因此退化草甸的恢复必须借助于外力,要借助于人的干预。
在模拟时,模型中鼠兔的冬季死亡率取较为特殊的-0.26,这样当4月份每个元胞中有5个有效洞口时有效洞口数年复一年地接近周期变化。这样处理其实就是固定鼠兔种群的动态,使得模拟中出现的鼠兔种群变化都是由于其他因素造成的,而不是来自鼠兔种群本身。
在很大程度上,建立的模型是理论上的,通过模拟可以得到有关草甸及鼠兔的一些一般性规律,但要将模型应用到实际还有很多因素要考虑。如放牧通过直接改变植被高度而改变鼠兔的环境容纳量;实际的草甸并非完全均质,存在植被上的差异,有的地方还有河流、道路等;还要考虑鼠兔的捕食者,且捕食者的分布也不是均匀的;年与年之间的出生率和死亡率也并非常数,受当年的植被状况、气象因素等影响。
[1]梁俊勋.介绍一种新型的实验动物——高原鼠兔[J].动物学杂志,1990,25(4):46-49.
[2]刘伟,王启基,王溪,等.高寒草甸“黑土滩”退化草地的形成原因及生态过程[J].草地学报,1999,7(4):300-307.
[3]Ioannis K.A model for influence of the greenhouse effect on insect and microorganism geographical distribution and population dynamics[J].BioSystems,1998,45(1):1-10.
[4]Wang J H,K ropff M J,Lammert B,et al.U sing CA model to obtain insight into mechanism of plant population spread in a controllable system:annual weeds as an ex ample[J].Ecological Modelling,2003,166(3):277-286.
[5]M atsinos Y G,T roumbis A Y.Modeling competition,dispersal and effects of disturbance in the dynamics of a grassland community using a cellular automaton model[J].Ecological Modelling,2002,149(1-2):71-83.
[6]王育,王小明,王正寰,等.高原鼠兔生境选择的初步研究[J].四川大学学报(自然科学版),2004,41(4):1041-1045.
[7]魏万红,曹伊凡,张堰铭,等.捕食风险对高原鼠兔行为的影响[J].动物学报,2004,50(3):319-325.
[8]江小蕾.植被均匀度与高原鼠兔种群数量相关性研究[J].草业学报,1998,7(1):60-64.
[9]刘汉武.高原鼠兔种群时空动态的元胞自动机模拟[D].西宁:中国科学院西北高原生物研究所,2008.
[10]杨振宇,江小蕾.高原鼠兔对草地植被的危害及防治阈值的研究[J].草业科学,2002,19(4):63-65.
[11]刘季科,张云占,辛光武.高原鼠兔数量与危害程度的关系[J].动物学报,1980,26(4):378-385.
[12]王谋,李勇,黄润秋,等.气候变暖对青藏高原腹地高寒植被的影响[J].生态学报,2005,25(6):1275-1281.
[13]李英年,赵亮,赵新全,等.5年模拟增温后矮嵩草草甸群落结构及生产量的变化[J].草地学报,2004,12(3):236-239.
猜你喜欢
舰船科学技术(2022年20期)2022-11-28
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
计算机工程与科学(2022年2期)2022-03-22
数学物理学报(2021年3期)2021-07-19
海外文摘·文学版(2021年3期)2021-05-19
智富时代(2019年4期)2019-06-01
智富时代(2019年4期)2019-06-01
活力(2019年21期)2019-04-01
智富时代(2018年5期)2018-07-18
