
自然干旱胁迫及复水处理对红秆寒竹生理特性的影响
2010-12-05 02:49应叶青魏建芬晏金凤
浙江农林大学学报 2010年4期
应叶青,郭 璟,魏建芬,晏金凤,路 英,方 伟
(浙江农林大学 浙江省现代森林培育技术重点实验室,浙江 临安 311300)
红秆寒竹 Chimonobambusa marmorea f.variegata为寒竹 Chimonobambusa marmorea的栽培变型[1],在浙江和福建等省有分布,世界各地多已有引种栽培[1-2],其株高为 80~120 cm,秆红色至粉红色,枝叶繁茂,叶片偶尔具不规则白色条纹,耐修剪,适于盆栽,观赏价值极高,为庭院及室内绿化的优良观赏竹种[2]。通过红秆寒竹的抗性研究,充分发挥其观赏特性,加快其开发利用,可丰富园林绿化竹种种类,为中国城乡绿化、室内装饰等提供高品质的观赏竹绿化苗木。然而到目前为此,国内外在观赏竹领域的研究相对较少,且主要集中在资源调查分类、选育栽培和园林应用上,而对观赏竹的环境适应性的研究甚少[3]。本研究通过自然持续干旱胁迫下红秆寒竹形态变化、生理响应的研究,初步了解其抗旱能力及抗旱机制,为该竹种的引种栽培及园林应用提供参考。
1 材料与方法
1.1 试验材料及设计
实验材料红秆寒竹于2001年从日本引种,在浙江林学院林木良种基地栽培,为防止冻害发生,冬天搭暖棚保温越冬。2004年6月将部分材料移栽上盆于温室栽培,部分继续野外自然栽培。2008年6月中旬于温室内进行分株换盆,盆栽土壤体积比为珍珠岩 ∶泥炭 ∶蛭石 =1∶2∶1的混合土,并浇透水。2008年9月底对它们进行干旱处理,实验前灌透水1次(连续3 d,每次浇至盆底托盘中有水渗出),以后不再浇水,自然持续干旱。实验从9月29日开始,分别在干旱胁迫0,10,20,30,40 d后采集红秆寒竹中部枝条生长充实的功能叶,测定叶绿素(Chl)质量分数、超氧化物歧化酶(SOD)活性、过氧化氢酶(CAT)活性、丙二醛(MDA)质量摩尔浓度等生理指标,采样重复3次·次-1。另外,在干旱处理的第40天傍晚对红秆寒竹复水,之后的1,3,5 d分别取样测定上述生理指标。
1.2 指标测定
土壤含水量采用烘干称量法测定:称取容器中部土样10 g,装入铝盒,在105~110℃下烘干6~8 h,计算土壤含水量。叶绿素质量分数测定采用浸提法;SOD活性测定采用氮蓝四唑法;用硫代巴比妥酸(TBA)反应法测定丙二醛质量分数[4]。CAT活性测定采用郝再彬等[5]方法。
1.3 数据统计分析
数据统计分析采用统计分析软件SAS进行。
2 结果与分析
2.1 干旱胁迫下土壤含水量及植株外形上的变化
干旱处理后,土壤含水量随胁迫持续发生了显著的变化(图1),土壤含水量由正常浇水时的80.6%下降至干旱第40天的9.2%,各观察时间点上土壤含水量差异显著。胁迫结束时,土壤含水量已经很低,但观察到红秆寒竹植株没有发生明显萎焉现象,叶色从翠绿色转变为黄绿色。

图1 断水后土壤含水量随时间的变化Figure 1 Change of soil water content with time after not supplying water
2.2 干旱胁迫对红秆寒竹叶绿素质量分数的影响
2.2.1 干旱胁迫对叶绿素总量的影响 叶绿素是绿色植物进行光合作用的主要色素,叶绿素质量分数与组成同植物光合作用关系密切。干旱胁迫对红秆寒竹的叶绿素总量的影响如表1所示:叶绿素总量在干旱胁迫 0~30 d间,由开始4.470 mg·g-1降到第30天的1.676 mg·g-1,在处理 0~30 d中每个观察点上,前后2次测定的叶绿素总量都有极显著的差异,这说明在胁迫初中期,干旱对红秆寒竹叶绿素总量影响很大;在干旱处理30 d以后及复水后5 d内,植株保持稳定的叶绿素总量,说明植株通过一定的自我调节基本适应了干旱胁迫状态。复水后(1~5 d),叶绿素总量基本保持不变,可能是叶绿素合成的器官受到一定程度的损害或者合成途径受到阻碍,以致在短期内难以恢复。
2.2.2 干旱胁迫对叶绿素a和叶绿素b质量分数的影响 由表1可以看出,叶绿素a和叶绿素b质量分数的变化趋势与叶绿素总量变化一致。在0~30 d内的每个观察点上,前后2次测定值均有极显著的差异,当叶绿素 a质量分数降到 1.219 mg·g-1,叶绿素 b质量分数降到 0.458 mg·g-1时,即处理 30 d,土壤含水量降到13.5%后,叶绿素a和叶绿素b质量分数基本稳定,复水后也没有明显的变化。
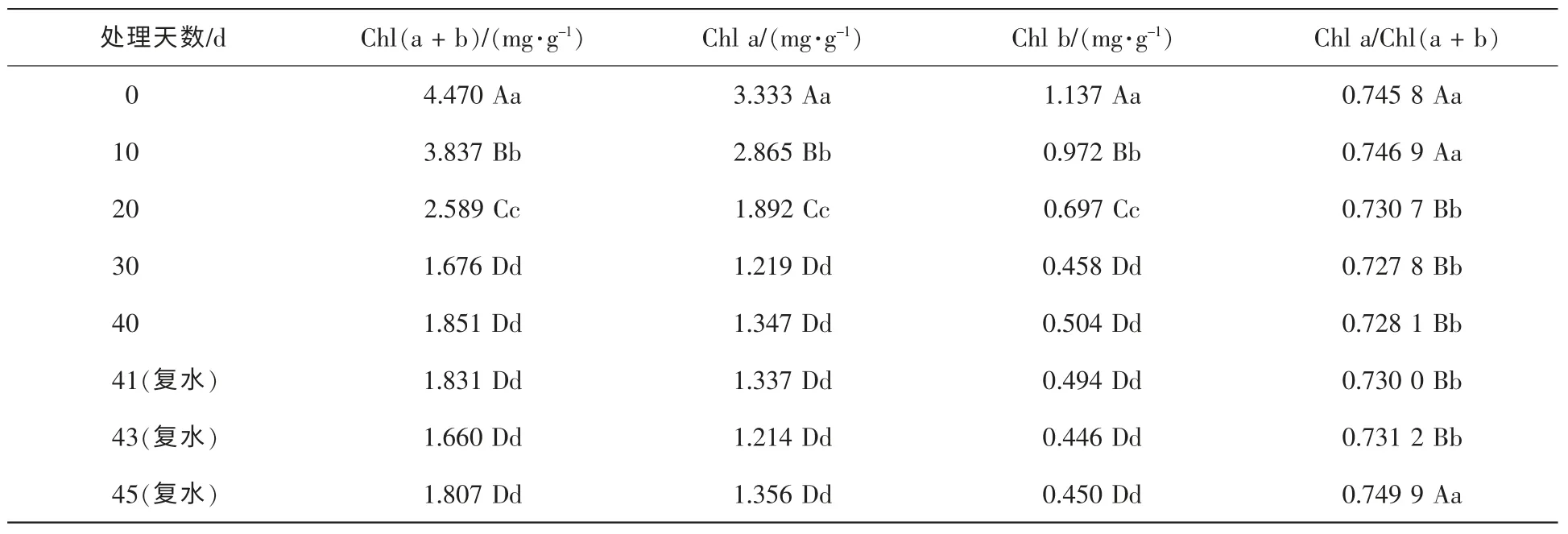
表1 红秆寒竹叶绿素质量分数随干旱时间的变化Table 1 Changes of chlorophyll content over time under drought stress
2.2.3 干旱胁迫对叶绿素a与叶绿素总量比值的影响 叶绿素a是植物叶绿体中的重要光能吸收色素,占叶绿素总量的74.58%(表1)。在自然干旱10 d,基质含水量为41.6%时,虽叶绿素a和叶绿素b及叶绿素(a+b)都极显著的下降,叶绿素a/叶绿素(a+b)仍处于稳定的水平,没有显著差异,表明在干旱胁迫初期两者下降幅度一致;在10~20 d阶段,叶绿素a降幅超过叶绿素b的降幅,叶绿素a/叶绿素(a+b)发生极显著变化;20~30 d阶段,叶绿素a与叶绿素(a+b)降幅一致;此后干旱至复水3 d时,叶绿素a/叶绿素(a+b)保持稳定水平,各色素质量分数没有显著变化。复水5 d后,叶绿素a质量分数增加虽没有达到显著水平,但叶绿素a/叶绿素(a+b)有了极显著变化,回复到正常水平。
2.3 干旱胁迫对红秆寒竹丙二醛质量摩尔浓度、SOD活性和CAT活性的影响
2.3.1 干旱胁迫对红秆寒竹丙二醛质量摩尔浓度的影响 由表2可以看出,随着干旱胁迫增强,红秆寒竹丙二醛质量摩尔浓度不断增加,由 10.23 μmol·g-1升至 19.96 μmol·g-1,变化达极显著水平。其中,有2个阶段丙二醛质量摩尔浓度保持在稳定水平,在胁迫10~20 d内丙二醛保持在一个稳定水平,在30 d时有显著下降,可能是在适度的干旱胁迫下,激活了植物体自身的抗旱调节系统,植物抗旱能力提高;在胁迫40 d时,丙二醛质量摩尔浓度达到19.96 μmol·g-1,增幅极显著,达到较高的水平,以后波动不大,一直处于较高的水平。在复水5 d后,丙二醛质量摩尔浓度降至14.48 μmol·g-1,回复到较低的水平,丙二醛的质量摩尔浓度在短时间内快速降到较低的水平,说明尽管土壤含水量降到9.2%,但持续时间不长时,植株没有受到很严重的破坏,能够在较短时间内恢复正常生长。
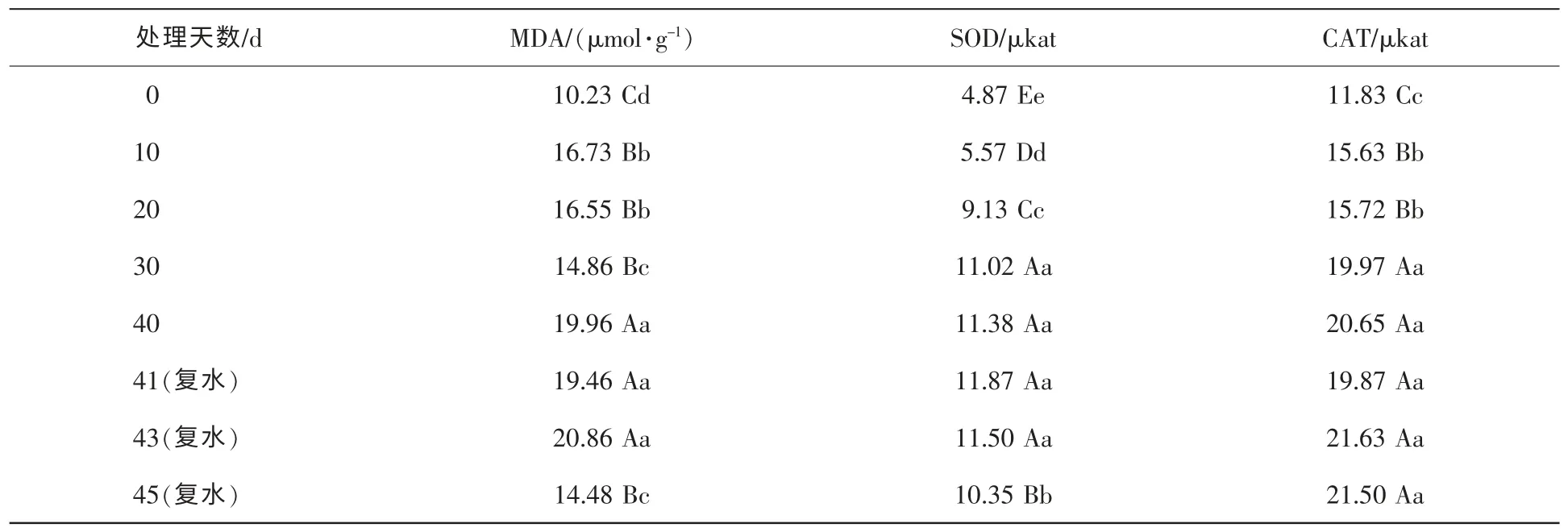
表2 红秆寒竹MDA质量摩尔浓度及SOD和CAT活性随干旱时间的变化Table 2 Changes of MDA content and SOD and CAT activity over time under drought stress
2.3.2 干旱胁迫对红秆寒竹CAT活性的影响 由表2可以看出:在干旱胁迫开始时,红秆寒竹CAT活性为 11.83 μkat。干旱处理 10 d和 20 d后,CAT活性分别达到 15.63 μkat和 15.72 μkat,也就是在干旱处理0~20 d内,CAT活性随着时间的增长而增加,并且变化达到了显著性水平;在干旱处理30 d时,CAT活性升至19.97 μkat,有极显著的变化,说明当土壤含水量降至23.3%时,植株体内过氧化氢积累达到了较高水平,自身防御机制发生作用以降低本身受伤害程度。在干旱处理30 d后及复水之后,则少有变化,维持在一个较高且稳定的状态。
2.3.3 干旱胁迫对红秆寒竹SOD活性的影响 干旱胁迫对红秆寒竹SOD活性影响如表2所示。在干旱胁迫下,红秆寒竹的SOD活性呈升高的趋势,干旱胁迫40 d时红秆寒竹的SOD活性是胁迫前的2.4倍,达到极显著水平。在干旱胁迫0~30 d内,植株SOD活性由水分正常状态下时的4.87 μkat升至干旱处理30 d后的11.02 μkat,在每个时间点上测定的SOD活性都有极显著的增幅。表明随着干旱胁迫的加强,红秆寒竹体内活性氧增加,SOD活性迅速提高,清除活性氧。干旱胁迫30 d后至复水3 d,SOD活性一直维持较为稳定的水平。但在复水5 d后,SOD活性降到10.35 μkat,降幅显著。
3 讨论
在干旱处理过程中,红秆寒竹叶绿素总量及叶绿素a、叶绿素b在处理的初中期持续下降,降幅达到极显著水平,这与林树燕等[3]在平安竹 Pseudosasa japonica‘Tsutsumiana’,铺地竹Pleioblastus argenteastriatus,小佛肚竹Bambusa ventricosa中研究发现叶绿素呈先上升后下降的结论不尽一致。这可能是与取样时间有关,3种观赏竹叶绿素高峰出现较早的为铺地竹和小佛肚竹(4 d),最迟为平安竹(8 d),而红秆寒竹第1次测定为第10天,以后应加大测定密度,以更为准确地分析土壤水分与红秆寒竹叶绿素质量分数变化的关系。叶绿素不仅是植物光合作用的主要色素,也对红秆寒竹的叶色造成影响,从而影响了植株的观赏性。干旱胁迫致使红秆寒竹叶绿素质量分数下降,并在复水后短期难以修复,因此,在红秆寒竹的栽培管理中要注意加强水分的管理,以保持其叶片色泽,保证红秆寒竹的观赏性。
CAT和SOD是植物体内重要的保护酶类。CAT可促使过氧化氢分解为分子氧和水,清除体内的过氧化氢。水分胁迫下植物的CAT和SOD活性与抗旱性呈正相关,抗旱性强的品种其活性先升高后下降,而抗旱性弱的品种一直降低[6-9]。抗旱强的品种较抗旱弱的品种能维持较高的SOD和CAT活性。在上升阶段,前者比后者SOD和CAT活性上升幅度要大;在下降区,前者比后者下降要慢一些[10]。在干旱胁迫下,红秆寒竹CAT和SOD活性均随着胁迫的加深呈增加的趋势,在处理30 d后维持在较高且稳定的水平,CAT和SOD协同作用,很好地避免了红秆寒竹受到活性氧的伤害。
丙二醛是膜脂过氧化的主要产物之一,具有很强的细胞毒性。丙二醛的积累与生长量呈负相关[11]。在胁迫过程中保持相对较低的丙二醛质量摩尔浓度和相对较高的酶活性这与其抗旱性强弱相关[12]。抗氧化酶活性和丙二醛质量摩尔浓度可作为植物抗旱资源筛选和利用的依据[13]。丙二醛质量摩尔浓度增幅小的品种对干旱忍耐较强,增幅大的品种耐旱力弱[14]。红秆寒竹在胁迫前30 d,土壤含水量已降到13.48%时,CAT和SOD活性维持在较高且稳定的水平,而丙二醛质量摩尔浓度处于一个较低的水平,表明较高的保护酶活性良好地保护了其叶片细胞膜完整性,红秆寒竹具有较强的耐旱能力。
[1]丁雨龙.观赏竹新品种引进与快速繁育[J].林业科技开发,2002,16(3):13-14.DING Yulong.The introduction and rapid multiplication of new varieties of ornamental bamboos[J].China For Sci Technol,2002,16(3):13-14.
[2]耿伯介,王正平.中国植物志:第9卷第1分册[M].北京:科学出版社,1996:328-329.
[3]林树燕,丁雨龙.3种观赏竹抗旱生理指标的研究及其综合评价[J].竹子研究汇刊,2006,25(2):7-9.LIN Shuyan,DING Yulong.Researches on indexes of drought resistance of three ornamental bamboo species and corresponding comprehensive evaluation.[J].J Bamboo Res,2006,25(2):7-9.
[4]邹琦.植物生理学实验指导[M].北京:中国农业出版社,2000.
[5]郝再彬,苍晶,徐仲.植物生理实验[M].哈尔滨:哈尔滨工业大学出版社,2004:113-114.
[6]王茅雁,邵世勤,张建华,等.水分胁迫对玉米保护酶系活力及膜系统结构的影响[J].华北农学报,1995,10(2):43-49.WANG Maoyan,SHAO Shiqin,ZHANG Jianhua,et al. Effect of water stress upon the activities of protective enzyme system and the structures of membrane system in maize[J].Acta Agric Boreali-Sin,1995,10(2):43-49.
[7]SCANDALIOS J G.Oxygen stress and superoxide dismutases[J].Plant Physiol,1993,101:7-12.
[8]JIMENEZ C,PICK U.Differential reactivity of b-carotene isomers from Dunaliella bardawil toward oxygen radicals[J].Plant Physiol,1993,101:385-390.
[9]蒋明义,杨文英,徐江,等.渗透胁迫诱导水稻幼苗的氧化伤害[J].作物学报,1994,20(4):733-738.JIANG Mingyi,YANG Wenying,XU Jiang,et al. Osmotic stress-induced oxidative injury of rice seedlings[J].Acta A-gron Sin,1994,20(4):733-738.
[10]周瑞莲,王刚.水分胁迫下豌豆保护酶活力变化及脯氨酸积累在其抗旱中的作用[J].草业学报,1997,6(4):39-43.ZHOU Ruilian,WANG Gang. Water stressinduced changes in protective enzyme activities and effects of proline enhancement on drought resistance in pea[J].Acta Pratacult Sin,1997,6(4):39-43.
[11]汪耀富,阎栓年,于建军,等.土壤干旱对烤烟生长的影响及机理研究[J].河南农业大学学报,1994,28(3):250-259.WANG Yaofu,YAN Shuannian,YU Jianjun,et al. Study on the effects of soil drought stress on the growth of fluecured tobacco and its mechanism[J].J Henan Agric Univ,1994,28(3):250-259.
[12]张怡,罗晓芳,沈应柏.土壤逐渐干旱过程中刺槐新品种苗木抗氧化系统的动态变化[J].浙江林学院学报,2005,22(2):166-169.ZHANG Yi,LUO Xiaofang,SHEN Yingbai.Dynamic change s of anti-oxidation system in new cultvars of Robinia pseudoacacia under gradual drought stress of soil[J].J Zhejiang For Coll,2005,22(2):166-169.
[13]李迎春,樊卫国,陈双林.干旱胁迫对梨属4个重要种幼苗膜脂过氧化和抗氧化酶活性的影响[J].浙江林学院学报,2008,25(4):437-441.LI Yingchun,FAN Weiguo,CHEN Shuanglin.Soil drought stress on embrane-lipid peroxidation and antioxidant enzymes in pear rootstock[J].J Zhejiang For Coll,2008,25(4):437-441.
[14]李雪莲,张国芳,谷艳蓉,等.4种多年生禾草苗期抗旱性比较研究[J].四川草原,2005(1):13-15.LI Xuelian,ZHANG Guofang,GU Yanrong,et al.Comparison about drought tolerance of four perennial forages during seedling[J].J Sichuan Grassl,2005(1):13-15.
猜你喜欢
今日农业(2021年14期)2021-11-25
今日农业(2020年13期)2020-12-15
河北画报(2020年10期)2020-11-26
河南水利年鉴(2020年0期)2020-06-09
建材发展导向(2019年11期)2019-08-24
英美文学研究论丛(2018年2期)2018-08-27
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
食品与生活(2014年2期)2014-03-04
雕塑(2000年4期)2000-06-24
