
太平洋牡蛎杂交三倍体与诱导三倍体的生长比较*
2011-01-10 09:41王昭萍于瑞海张跃环李晓瑜李雅琳郭希明
中国海洋大学学报(自然科学版) 2011年7期
孔 静,王昭萍**,于瑞海,张跃环,刘 剑,李晓瑜,李雅琳,郭希明
(1.中国海洋大学海水养殖教育部重点实验室,山东青岛266003;2.青岛崂东海珍品良种培育有限公司,山东青岛266105;3.Haskin Shellfish Research Laboratory,Institute of Marine and Coastal Sciences,Rutgers University,Port Norris,New Jersey 08349 USA)
贝类多倍体[1]的诱导主要是研究贝类三倍体。三倍体较二倍体具有绝对的生长优势,个体大且繁殖季节死亡率低。又因其性腺高度不育,在繁殖季节,仍具有丰富的糖原,口感好,很受消费者喜欢,具很高的经济价值[2]。三倍体贝类比正常贝类个体多一套染色体,不会对自然群体造成物种污染,既提高了贝类产量,增加经济价值,又为海产贝类育种提供了新的育种概念等,均是三倍体贝类的研究的重要原因。
以往研究从理化诱导和生物方法均获得过贝类三倍体。就牡蛎多倍体的诱导而言,最先采用的就是化学诱导方法,Stanley等[3]最早采用不同浓度的细胞松弛素B处理美洲牡蛎(Crassostrea virginica)受精卵,得到8个月三倍体子代。之后长牡蛎(C.gigas)[4-7]、食用牡蛎(Ostrea edulis)[8]、悉尼岩牡蛎(Saccostrea commercialis)[9]和僧帽牡蛎(O.cucullata)[10]等牡蛎也成功诱导出三倍体。因理化诱导极易受外界因子如温度、密度等的影响,三倍体诱导率无法达到100%,但生物杂交方法通过四倍体牡蛎和二倍体牡蛎杂交,可产生全三倍体牡蛎[11],前景较好[12]。
以往研究证明了杂交获得三倍体的诱导率高,方法简单,避免理化诱导对受精卵造成的影响,提高了孵化率和变态率[13],也初步证实了三倍体牡蛎的生长优势[11,24]。但如何利用不同地域的二倍体群体进行大规模生产,进一步证实和提高杂交三倍体性状,研究不多。本实验利用中国太平洋牡蛎自交、诱导及中国二倍体太平洋牡蛎与美国四倍体太平洋牡蛎杂交,对诱导获得的三倍体群体及杂交获得的三倍体,进行生长发育比较来评估2种获得三倍体的方法,为三倍体诱导方法的选择及其今后的大规模生产奠定了基础。
1 材料与方法
1.1 亲贝的来源与促熟
中国太平洋牡蛎取自威海文登海区。美国四倍体太平洋牡蛎(雄)来自美国华盛顿州。亲贝在崂东海珍品培育有限公司育苗池网箱中暂养促熟。
1.2 实验设计
1.2.1 实验方案 本试验采用3个试验组,分别为2NC、3NC、3N H;其中,2NC二倍体对照组:中国二倍体太平洋牡蛎自交组;3NC诱导三倍体群体组:中国二倍体太平洋牡蛎(2NC)雌♀×中国二倍体太平洋牡蛎(2NC)雄♂,再经150μmol/L的6-二甲氨基嘌呤(6-DMAP)抑制受精卵第二极体的排放;3NH杂交三倍体组:中国二倍体太平洋牡蛎(2NC)雌♀×美国四倍体太平洋牡蛎(用4NA表示)雄♂。
1.2.2 受精 取性腺少量细胞,显微镜下鉴别雌雄,并检查精卵的成熟状况。雌雄牡蛎严格分开,以免精子污染。用解剖刀逐个刮取成熟性腺部,将卵用250目筛绢过滤后至标记好的小桶中熟化1 h,舍弃上层卵液。受精前检查卵子是否已经受精,将获得的精、卵按照实验设计组合受精,迅速混合,搅拌均匀,于孵化桶中孵化。整个操作过程中,各实验组严格隔离,使其不受外来精、卵的影响。
1.3 幼虫及稚贝的培育
受精卵经24 h发育为D形幼虫。选优后,幼虫培育密度设为8~10个/mL,培育水温21℃,饵料每天投喂3次,前期为金藻,后期为扁藻与小球藻(1∶1)混合投喂。每天全量换水,为防止不同实验组幼虫之间混杂,换水网箱单独使用。定期调整密度,消除养殖密度对幼虫生长的影响。
1.4 数据测量
(1)分别测量各组卵径、卵裂率、D幼率和D形幼虫大小。
(2)生长用壳高、壳长衡量。幼虫阶段每3 d测量1次;稚贝阶段在40、70、110和140日龄测量。每次测量设3个重复,每个重复随机测量30个个体。(3)测定浮游期各试验组幼虫的存活率。
(4)测定各试验组倍性。幼虫期取D形幼虫以及在稚贝期取个体鳃组织,收集到1.5 m L离心管中,加入DAPI荧光染液1 m L,振荡器震荡0.5 min,用Partec PAⅡ流式细胞仪(FCM)进行倍性检测。
1.5 数据处理
采用Excel作图,SPSS 13.0统计软件进行分析处理,不同试验组间数据的比较采用单因素方差分析方法,差异显著性设置为P<0.05。
2 结果
2.1 卵径、卵裂率、D幼率、D形幼虫大小及倍性
试验各组的卵径、卵裂率、D幼率、D形幼虫大小及三倍体诱导率见表1。3个试验组在卵径和卵裂率方面均无显著性差异(P>0.05),杂交三倍体的D形幼虫规格显著大于诱导群体组和二倍体自交组(P<0.05)。但在D幼率上看,3N H组个体显著高于3NC群体组(P<0.05),与对照二倍体组无显著差异(P>0.05)。在三倍体诱导率方面,杂交获得的三倍体率为100%,明显高于诱导群体组获得的三倍体倍化率。
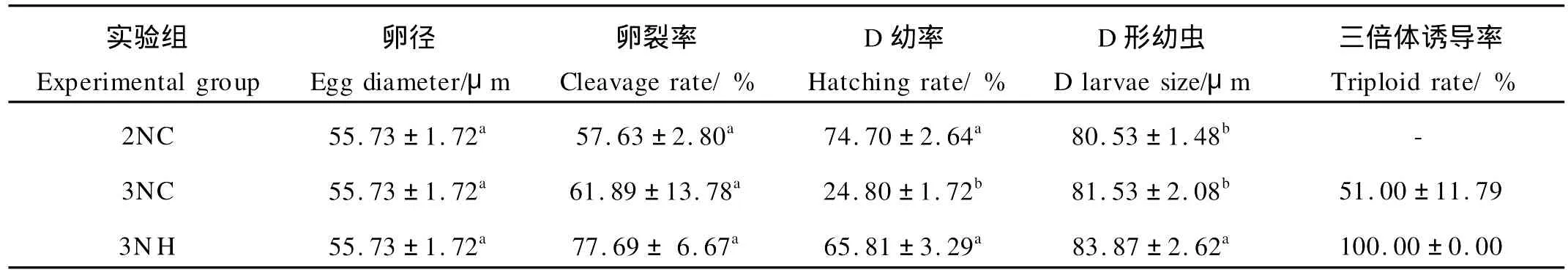
表1 诱导三倍体群体与杂交三倍体卵径、卵裂率、D幼率、D形幼虫大小及倍性测定的比较Table 1 Comparison of egg-diameter,cleavage rate,hatching rate,D larval size and ploidy for trip loid Pacific oyster by hybridized and induced
2.2 浮游期幼虫存活率
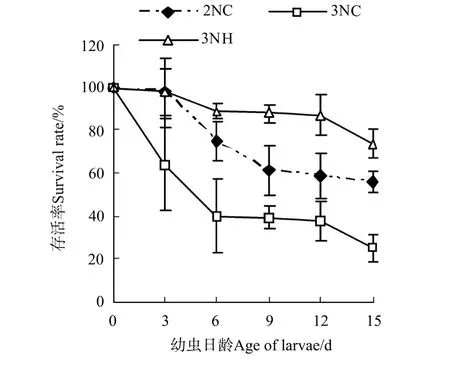
图1 诱导三倍体群体与杂交三倍体幼虫存活比较Fig.1 Survival rates of trip loid Pacific oysters by hybridized and induced
将D形幼虫的存活率定义为100%,由各试验组存活曲线(见图1)可以看出,随着幼虫日龄的增长,各组幼虫的存活率下降,且各试验组的存活率变化不同。3日龄时,2NC和3N H组存活率彼此间无显著差异(P>0.05),但显著高于3NC群体(P<0.05);6~15日龄期间,各试验组的幼虫存活率顺次为3NH>2NC>3NC群体,且各组间差异显著(P<0.05)。
2.3 杂交三倍体与诱导三倍体群体生长比较
2.3.1 幼虫的生长 各试验组幼虫的生长曲线如图2。3组的壳高生长速度分别为2NC组(13.73±0.98)μm/d,3NC群体组(15.80±0.83)μm/d和3NH组(16.41±0.61)μm/d。9日龄以前,各试验组幼虫的大小彼此间无显著差异;9~18日龄期间,3NH组壳高生长速度最快,表现出明显的生长优势。整个幼虫浮游期间,从幼虫的生长速度上看:3NH组>3NC群体组>2NC组,且3组间差异均显著(P<0.05)。
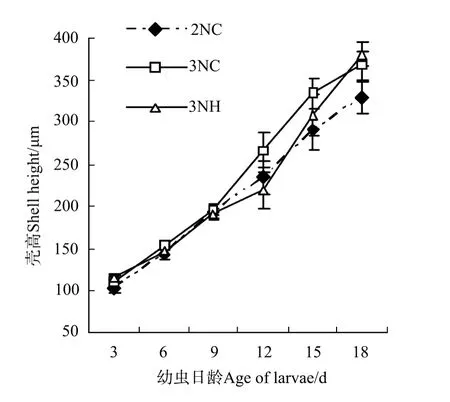
图2 诱导三倍体群体与杂交三倍体幼虫生长比较Fig.2 Shell height of trip loid Pacific oysters by hybridized and induced
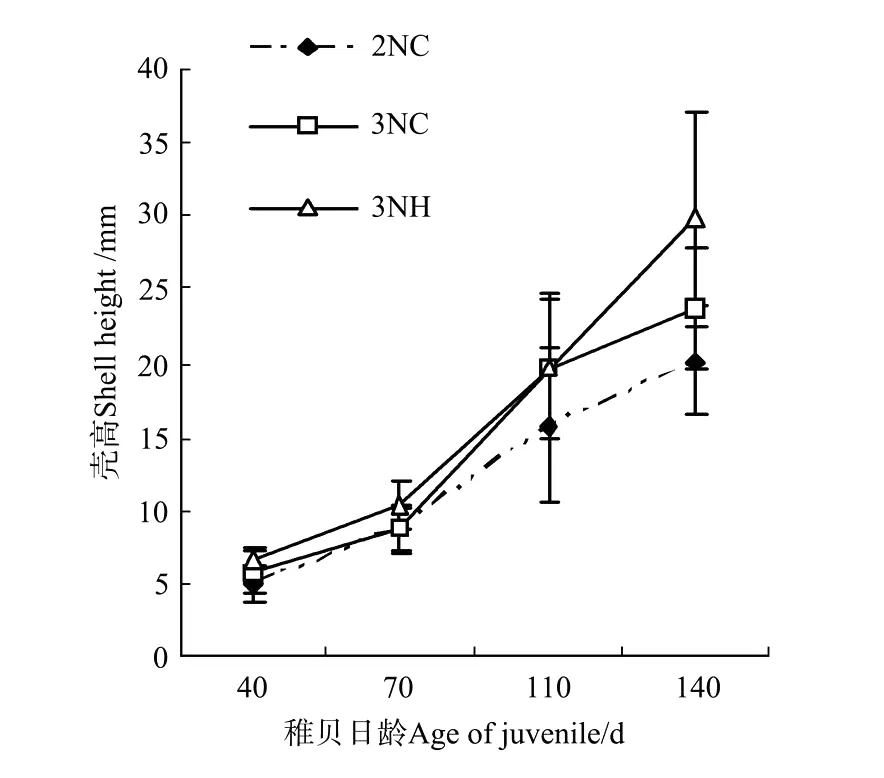
图3 诱导三倍体群体与杂交三倍体稚贝壳高的生长比较Fig.3 Shell height of juvenile of triploid Pacific oysters by hybridized and induced

图4 诱导三倍体群体与杂交三倍体稚贝壳长的生长比较Fig.4 Shell length of juvenile of triploid Pacific oysters by hybridized and induced
2.3.2 稚贝的生长 各试验组稚贝期壳高和壳长的生长曲线如图3、4。随着培养时间的延长,3组壳长和壳高均保持相同的增长趋势。稚贝期3组的壳高生长速度分别是2NC组(0.151±0.025)mm/d,3NC群体组(0.179±0.028)mm/d和3NH组(0.230±0.065)mm/d;壳长的生长速度分别是2NC组(0.099±0.010)mm/d,3NC群体组(0.110±0.016)mm/d和3NH组(0.132±0.031)mm/d。稚贝期各试验组壳高、壳长的生长速度:3NH>3NC群体>2NC,且彼此间的生长速度差异显著(P<0.05)。
2.4 稚贝期三倍体群体中二倍体和三倍体与杂交三倍体生长比较
在稚贝期(110日龄)对诱导三倍体群进行了倍性检测,并对分离出的三倍体和二倍体以及杂交三倍体进行了生长测量,壳高和壳长的生长情况如表2。结果表明,到110日龄的稚贝期,诱导群体中的三倍体在壳高和壳长2方面稍小于杂交三倍体,但无明显差异(P>0.05),而诱导群体中的二倍体则比二倍体对照组生长快,尤其壳高明显大于对照组。
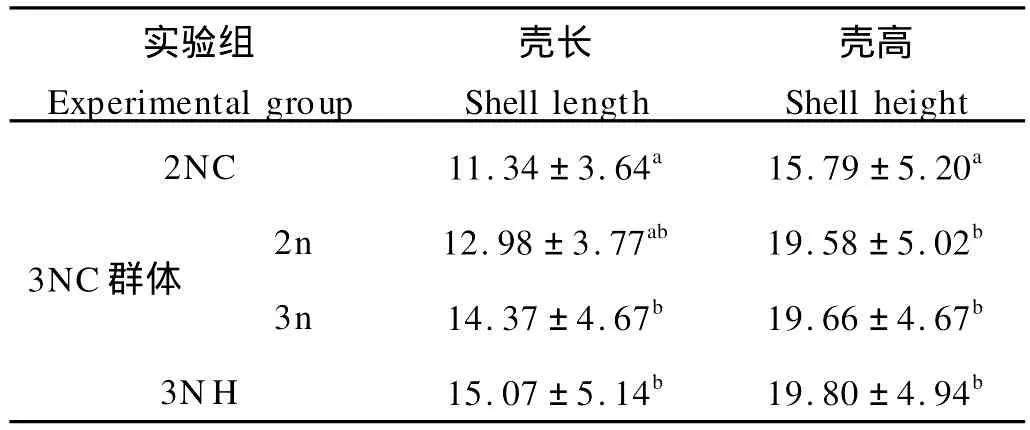
表2 稚贝期(110日龄)三倍体群体中二倍体和三倍体与杂交三倍体生长性状比较Table 2 Comparison of shell length and shell height of juvenile of dip loid and trip loid Pacific oyster by hybridized and induced(Day 110)/mm
3 讨论
本实验所选6-二甲氨基嘌呤诱导三倍体的方法是因为6-二甲氨基嘌呤作为诱导剂,诱导率高,毒性小,操作方便,价格便宜[14],现已被广泛采用。
杂交、诱导2种方法获得的三倍体卵裂率高,生长速度快,D形幼虫大小均大于二倍体对照组,但D形幼虫率很低。尤其是诱导获得的三倍体群体组D形幼虫率低,由于6-DMAP的影响,使得牡蛎幼虫出现畸形,该组死亡率在前期迅速下降。但杂交三倍体组存活率很稳定。应与化学诱导剂对受精卵产生的影响有关,这与田传远等[15]结果一致。12日龄以后,各组均出现幼虫大量沉底现象,阙华勇等[13]杂交多倍体实验中也有类似现象出现。与对照组相比,杂交三倍体和诱导三倍体群体从浮游幼虫阶段就表现出生长优势,此结果与王昭萍等[16]结论一致。从三倍体率来看,杂交获得100%的三倍体,较诱导群体组诱导率高且稳定。
通过对诱导三倍体群体和杂交三倍体的对比,从卵裂率、D幼率、存活率、生长发育和倍性等方面均体现了杂交三倍体具有优势。在稚贝期将诱导三倍体群体中的三倍体与杂交三倍体进行生长比较,杂交三倍体稍大于前者,但是差异不显著,这与王昭萍等[27]结果相吻合。分析本实验诱导三倍体群体与杂交三倍体之间的生长差异原因可能有2个:一是在诱导三倍体群体中二倍体含量过高,降低了三倍体的实际生长水平;二是杂交三倍体是中美牡蛎杂交,即为两种不同地理群体杂交,表现出了杂种优势。而诱导群体中的二倍体则比二倍体对照组生长快,可能是诱导群体中的二倍体经过了药物处理所致。
杂交获得三倍体即为诱导多倍体方法中的生物方法,该方法简单易操作,三倍体率高达100%,避免了理化处理对胚胎发育和环境的影响。这同陶抵辉、于瑞海等[17]观点一致。此结果同时也验证了杂交三倍体在生产应用上的可行性,同先前的于瑞海等[18]研究结果一致。
杂交获得三倍体的方法具有很多优点,但杂交方法中的四倍体亲本的获得是该方法的关键,也是瓶颈。虽有很多种养殖牡蛎[8,19-21]进行四倍体诱导,但目前只有太平洋牡蛎、美洲牡蛎和近江牡蛎建立了稳定的四倍体群系[22-24]。故四倍体的研究仍是多倍体方面需要继续探索的一个方向。
阙华勇等进行了四倍体太平洋牡蛎(G)和二倍体近江牡蛎(A)的杂交试验[25]。种间杂交子代GGA组幼虫个体较GA组大,眼点幼虫形成也在GA组之前。同时证实了杂交三倍体兼具了两亲本的特性,如适盐范围变宽了。而本实验两亲本太平洋牡蛎分别来自于中国威海和美国华盛顿,为种内杂交三倍体子代。种内杂交,是目前杂种优势利用的主要途径[26]。本实验并未对子代的生理等进行试验,不能确定是否兼具不同地理群体两亲本的优势,是本实验需要继续研究的一个方向。
牡蛎作为水产经济贝类之一,本实验通过对杂交三倍体和诱导三倍体群体的生长状况具体比较,验证杂交的方法获得的全三倍体,提高了牡蛎生产的产量,存活率也很高,适合在生产上广泛应用,是加速贝类养殖业的发展的一个可行的重要方向。
致谢:感谢青岛亚特兰生物科技有限公司提供四倍体牡蛎亲贝;青岛崂东海珍品良种培育有限公司郭希瑞、马强、孙军委等对本试验的帮助。
[1] 王昭萍,王如才,张筱兰.贝类多倍体育种研究及应用现状[J].齐鲁渔业,1999,16(5):32-33.
[2] 王昭萍,王如才,于瑞海.多倍体贝类的生物学特征[J].青岛海洋大学学报,1998,28(3):399-404.
[3] Stanley J G,Allen S K,Hidu H.Ployploidy induced in the American oyster Crassostrea virginica withcytochalasin B[J].Aquaculture,1981,23:1-10.
[4] A llen S K,Dow ning SL,Chaiton JC,et al.Chemically and p ressure induced triploidy in the Pacific oyster,Crassostrea gigas[J].Aquaculture,1986,57:359-360.
[5] Yamamoto S,Sugawara Y,Nomura T,et al.Induced triploidy in Pacific oyster,Crassosyrea gigas,and performance of trip loid larvae[J].Tohoku Journal of Agricultural Research,1988,39(1):47-59.
[6] Yamamoto S,Sugawara Y,Nomura T.Chemical and thermal control of triploid production in Pacific oysters and mussels,with regard to controlling meiotic maturation[M].∥Hoshim,Yamashita O.Advances in invertebrate rep roduction.Amsterdam:Elsevier,1990:455-460.
[7] Desrosiers R R,Gerard A,Peignon JM,et al.A novelmethod to produce triploids in bivalvemolluscs by the use of 6-dimethylaminopurine[J].Journal of Experimental Marine Biology and Ecology,1993,170(1):29-43.
[8] Gendreau S,Grizel H.Induced trip loidy and tetrap loidy in the European flat oyster,Ostrea edulis L[J].Aquaculture,1990,90:229-238.
[9] Nell J A,Cox E,Smith IR,et al.Studies on triploid oysters in Australia.Ⅰ.The farming potential of triploid Sydney rock oysters Saccostrea commercialis(Iredale and Roughley)[J].Aquaculture,1995,137:355-358.
[10] 曾志南,陈木,林琪,等.僧帽牡蛎三倍体的研究[J].海洋通报,1994,13(6):34-40.
[11] Guo X,DeBrosse G A,A llen S K.All-triploid Pacific oysters(Crassostrea gigas Thunberg)produced by mating tetraploids and diploids[J].Aquaculture,1996,142:149-161.
[12] 王昭萍,李慷均,于瑞海.贝类四倍体育种研究进展[J].中国海洋大学学报:自然科学版,2004,34(2):195-202.
[13] 阙华勇,张国范,刘晓,等.雄性四倍体与雌性二倍体杂交培育全三倍体长牡蛎(Crassostrea gigas)的研究[J].海洋与湖沼,2003,34(6):656-662.
[14] 王冬群,李太武.6-DMAP在贝类多倍体诱导中的应用[J].河北渔业,2003(1):3-4.
[15] 田传远,梁英,王如才.6-DMAP诱导太平洋牡蛎三倍体-诱导因素对孵化率和D幼畸形率的影响[J].中国水产科学,2000,7(1):110-112.
[16] 王昭萍,姜波,孔令峰,等.利用四倍体与二倍体杂交规模化培育全三倍体太平洋牡蛎苗种[J].中国海洋大学学报:自然科学版,2004,34(5):742-746.
[17] 陶抵辉,刘明月,肖君泽.生物多倍体诱导方法研究进展[J].生命科学研究,2007,11(4):6-13.
[18] 于瑞海,王昭萍,施坤涛.单体全三倍体太平洋牡蛎育苗技术的研究[J].海洋湖沼通报,2008(3):115-120.
[19] Stephens L B,Downing SL.Inhibition of first polar body formation in Crassostrea gigas produces tetraploids,notmeioticI triploids[J].J Shellfish Res,1988,7(3):550-551.
[20] Supan J E,Allen S K,Wilson C A.Tetraploid eastern oysters:An arduous effort[J].J Shellfish Res,2000,19(1):655.
[21] 容寿柏,李一民,刘绍琼,等.用冷热休克诱导四倍体近江牡蛎[J].湛江水产学院学报,1992,12(2):18-21.
[22] Eudeline B,Allen S K,Guo X.Optimization of tetraploid induction in the Pacific oysters,Crassostrea gigas,using the first polar body as a natural indicator[J].Aquaculture,2000,187:73-84.
[23] Guo X,Allen S K.Viable tetraploid Pacific oyster(Crassostrea gigas Thunberg)produced by inhibiting polar body Iin eggs fromtriploids[J].Mol Mar Biol Biotech,1994,3(1):42-50.
[24] Guo X,Wang Y,Xu Z,et al.Chromosome set manipulation in shellfish[M].∥Burnell G,Allam G.New technologies in aquaculture improving production efficiency,guality and environmental management.Cambridge:Waodhead Publishing Ltd,2009,165-194.
[25] Que Huayong,Allen S K.Hybridization of tetraploid and diploid Crassostrea gigas(Thunberg)with diploid C.ariakensis(Fujita)[J].Journal of Shellfish Research,2002,21(1):137-143.
[26] 李云玲,孙振兴,张明青.贝类杂交育种技术及其应用[J].齐鲁渔业,2005,22(12):22-24.
[27] Wang Zhaoping,Guo Ximing,Allen S K,et al.Heterozygosity and body size in triploid Pacific oysters,Crassostrea gigas Thunberg,produced from meiosis II inhibition and tetraploids[J].Aquaculture,2002,204:337-348.
猜你喜欢
科学养鱼(2021年12期)2022-01-14
科学养鱼(2021年11期)2022-01-11
热带农业科技(2020年3期)2020-12-15
食品与生活(2019年12期)2019-05-15
现代园艺(2017年13期)2018-01-19
江苏农业科学(2017年16期)2017-10-27
现代农业科技(2017年8期)2017-06-10
中成药(2016年4期)2016-05-17
中国果业信息(2015年6期)2015-01-23
河北渔业(2014年4期)2014-07-08
