
未净化Z:ZCLA长爪沙鼠封闭群繁育及遗传稳定性分析
2011-02-01 09:28李长龙石巧娟卢领群柯贤福戴方伟萨晓婴
中国比较医学杂志 2011年6期
李长龙,石巧娟,卢领群,柯贤福,戴方伟,萨晓婴
(浙江省医学科学院 浙江省实验动物中心,杭州 310013)
长爪沙鼠(Mongolian Gerbil,Meriones unguiculatus)俗称蒙古沙鼠,属于啮齿目,仓鼠科,沙鼠亚科,沙鼠属,又称长爪沙土鼠、蒙古沙鼠和黄耗子等,野生长爪沙鼠主要分布于我国内蒙古及其毗邻的干旱和半干旱地区[1],是一种正在开发的“多功能实验动物”,现已广泛的应用于神经学、寄生虫学、病毒学、细菌学、内分泌学、遗传学、血液学、脂类和糖代谢、肿瘤学等研究中具有极为重要的价值[2-4]。1935年日本学者从我国东北和蒙古东部捕捉野生长爪沙鼠并在大连卫生所驯养,后经日本北里研究所的长野博士和日本实验动物中心研究所野村博士进一步驯化和实验动物化,后引种到美、英、法等国。20世纪60年代开始作为实验动物应用于医学研究。日本国家实验动物中心(NIBIO)现已培育成3个长爪沙鼠近交系,主要由无毛、白化和癫痫敏感等品系[5]。我国有两个主要的长爪沙鼠群体,分别保存于浙江省实验动物中心和首都医科大学[1]。浙江省实验动物中心从1978年从内蒙古捕捉野生长爪沙鼠并开始人工繁殖,经过30多年的实验动物化研究,培育出具有自主知识产权的未净化长爪沙鼠封闭群-Z:ZCLA长爪沙鼠[6]。该群体目前是国内种群维持时间最长、应用单位最多、使用量最大、基本生物学特性研究较为完整的群体[1]。
线粒体DNA(mitochondrial DNA,m tDNA)是高等动物唯一的核外遗传物质。与核基因组相比,线粒体基因组结构简单、进化速度快(核基因的5~10倍),呈严格的母性遗传,遗传行为相对独立,变异发生的几率相对稳定、无组织特异性、提取方便[7]。由于mtDNA的这些独特遗传特性,已被广泛地用于物种起源与进化、生物分类、及群体遗传结构等方面的研究[8-10]。在进化分析时,研究者往往选择不同的区域进行不同时间尺度的进化分析。线粒体控制区(D-Loop region)是m tDNA中进化最快、变化最复杂的区域,其高突变率以及母系遗传方式使之成分析具有血统关系的群体内遗传稳定性有效的遗传标记[11]。管敏强等[12]利用线粒体控制区来分析封闭群小鼠的遗传稳定性。目前国家标准对于封闭群实验动物的遗传检测主要是以生化标记和基因多态性为主。线粒体控制区高度的种内群体遗传多态性特点,使之成为遗传检测的有效方法[13]。本文利用线粒体 D-Loop区高突变率的特点,分析本中心未净化Z:ZCLA长爪沙鼠的遗传稳定性。
1 材料和方法
1.1 动物来源及繁育方案
浙江省实验动物中心1978年从内蒙古捕捉野生长爪沙鼠,采用国际上培育远交封闭群品系的方法-组间循环交配法,进行封闭驯养、繁殖、选育,维持种群,经过将近30多年,46代封闭选育,已育成一个特性稳定,繁殖性能高的未净化长爪沙鼠新品系,定名为Z:ZCLA长爪沙鼠,并通过浙江省创新科技成果鉴定。
1.2 饲养条件和生长指标测定
繁殖群动物与生产群动物均饲养在未净化实验动物设施中。采用排气扇自然通风方式,氨气浓度控制在 20 mg/m3以下;自然采光;工作照度为150~300 lux;噪声控制在69 dB以下;对温湿度进行相对控制,温度18~30℃,日温差小于3.5℃;相对湿度60% ~90%;门窗设计防野鼠和昆虫的铁网;整个环境以及垫料、笼具、水瓶定期用新洁尔灭、百毒杀、过氧乙酸等消毒,以控制动物特有传染病和人畜共患病,落下菌数控制在少于40个/皿h。我国目前没有长爪沙鼠饲料标准,我中心在长期实验动物化进程中探索一套出行之有效的饲料配方。在上述营养条件下,随机选择40对长爪沙鼠,记录其繁殖胎次、产子数等数据。记录其仔鼠的体重和体长等变化情况。
1.3 遗传稳定性分析
1.3.1 动物来源和基因组 DNA的抽提:Z:ZCLA长爪沙鼠来自浙江省实验动物中心生产群。选择遗传距离较远的个体的非亲代、非同胞的雌鼠33只进行试验。所有个体采集肝脏组织样低温保存,采用常规苯酚、氯仿提取基因组DNA,DNA样品稀释50 ng/μL备用。

表1 Z:ZCLA长爪沙鼠饲料营养成分Tab.1 The food composition for Mongolian Gerbil
1.3.2 引物合成:按照引物设计原则,在长爪沙鼠线粒体控制区内设计 1对引物。上游 5'-TTCC CAGGACATCAAGAAGG-3',下游 5'-GGGTGTGGC TAGGCTAAGTG-3'。引物均由上海美吉生物技术有限公司合成。
1.3.3 PCR扩增:PCR扩增反应体系为25μL,其中含有基因组 DNA约 50 ng,0.20 mM的引物,200 mM的 dNTPs,2.0 mM,1.0 UTaq DNA聚合酶(TaKaRa Biotech,China)。扩增条件为:预变性94℃ 4 min;变性94℃ 45 s,退火60℃ 1 min,延伸72℃1 m in,共35个循环;最后72℃ 延伸8 m in。
1.3.4 测序和比对:扩增产物按照DNA片段纯化试剂盒(Tiangen Biotech,China)说明书方法纯化,纯化产物送上海美吉生物技术有限公司进行测序。测序结果用DNAMAN软件进行拼接并与已发表的长爪沙鼠线粒体控制区序列进行比对[14],发现可疑的变异位点后用Chormas软件读取测序的峰图记进一步分析确认。
1.4 统计学方法
2 结果与分析
2.1 饲养繁殖情况
Z:ZCLA长爪沙鼠属于杂食性动物,浙江省实验动物中心(浙江省实验动物饲料生产基地)生产的颗粒饲料(包括:繁育饲料和育成饲料)能够够满足长爪沙鼠的营养需求,饲喂效果较好,经过近30年的饲喂,长爪沙鼠仔鼠生长发育良好,成年长爪沙鼠毛色光亮,运动敏捷,未发现营养不良情况。
据实验观察,长爪沙鼠一夫一妻交配行为较明显,离乳后应选择生长发育良好的仔鼠进行配对,以保证后代的质量。长爪沙鼠是多次发情动物,在人工饲养条件下可全年交配繁殖。统计40对长爪沙鼠,共计455胎产子情况(见图1),其平均每胎产仔7.01只,且每胎产仔数大多在5~9只,占总胎数的79.34%。从胎间隔上看:观察100对母鼠不同生产胎次的间隔,胎间隔最短20 d,最长127 d,胎间隔大多在20~60 d之间,占总数的72.16%。在繁殖期内,有时发生配对雌(或雄)鼠死亡,补充配对时,往往发生咬斗现象,甚至咬伤致死,随着培养代数的增加,这种咬斗现象减少,持续时间亦缩短。长爪沙鼠和大小鼠一样,有产后即时交配受孕的特性。

图1 长爪沙鼠每胎产子数及其比例Fig.1 Number of birth and the ratio in Mongolian Gerbi
长爪沙鼠仔鼠出生后测量体重、体长、尾长,观测到116 d。检测结果经统计学分析发现:长爪沙鼠从出生到26 d体重、体长、尾长的增长比较平稳,在26 d~95 d的时间里,沙鼠各项生长发育指标的增长较快,95 d后增长又趋于平稳,甚至停止。这与沙鼠的生长发育规律相符。从体重和体长指标上比较发现,在离乳前雌、雄差异不明显,离乳后的整个发育期,雄鼠的体重和体长均大于雌鼠(详见图2~3)。

图2 长爪沙鼠日龄和体重Fig.2 Age and weight body in Mongolian Gerbil

图3 长爪沙鼠日龄和体长Fig.3 Age and stem length in Mongolian Gerbil
2.2 遗传学稳定性检测结果
2.2.1 PCR扩增结果:33只长爪沙鼠DNA样品全部扩增出特异性条带,扩增产物经过琼脂糖凝胶电泳显示与预期产物片段大小基本一致(图4)。
2.2.1 多态性分析:测序结果显示,33只Z:ZCLA
长爪沙鼠线粒体D-Loop序列完全一致,与之前报道的长爪沙鼠无任何差别。未见序列多态性和序列不稳定性(图5,图6)

图4 D-Loop区域扩增产物电泳图(部分) M:DNA DL5000 MarkerFig.4 The product of PCR in agarose gel electrophoresis
3 讨论
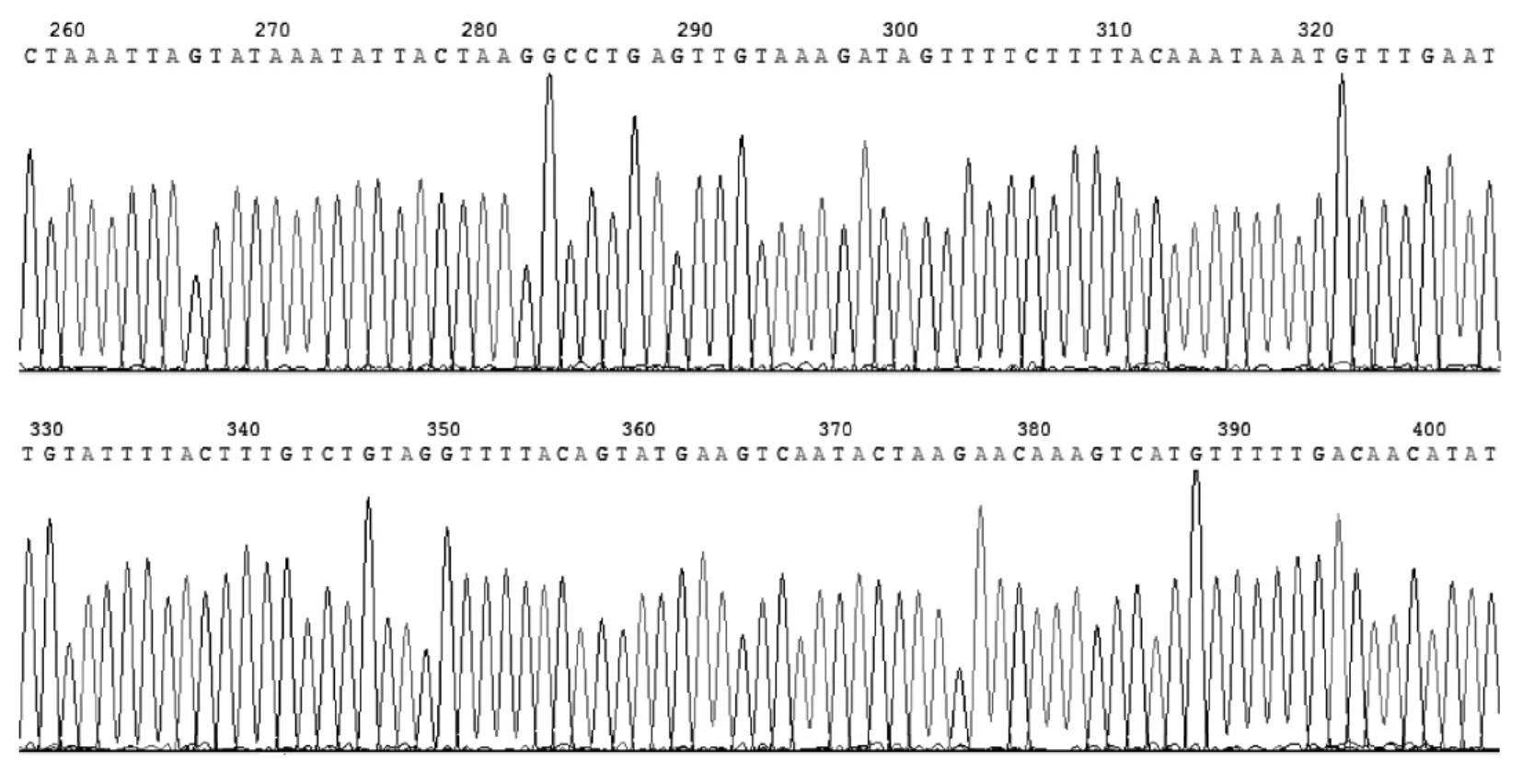
图5 D-Loop区域测序峰图(部分)Fig.5 The result of campare in sequence from gerbil(partical)

图6 D-Loop区域扩测序结果比对图(部分)DNAMAN 5.5 Fig.6 The result of campare in sequence from gerbil(partical)
Z:ZCLA长爪沙鼠是国内第一个从野生长爪沙鼠培育成功的、维持时间最长、使用最广、生物学特性研究较系统的特性稳定、繁殖率高的长爪沙鼠封闭群新品系,属国内首创。目前Z:ZCLA长爪沙鼠已被浙江省医学科学院、上海第二军医大学、重庆第三军医大学、徐州医学院、浙江大学医学院等十几个单位用于寄生虫、脑出血、微生物学、脂类和糖类代谢方面的研究,取得了重大科技成果。特别是浙江省疾病预防控制中心朱智勇研究员在国内外首次发现Z:ZCLA长爪沙鼠是流行性出血热病毒(EHFV)敏感的实验动物,是研究EHFV特性和疫苗研制的首选实验动物。利用Z:ZCLA长爪沙鼠肾细胞作培养基质,在世界上首创流行性出血热疫苗(I类),获国家科技进步一等奖。浙江天元生物药业股份有限公司已使用60万多只Z:ZCLA长爪沙鼠生产流行性出血热疫苗用于防病,取得十分显著的社会效益和经济效益。
遗传质量控制是实验动物质量控制的重要组成部分,对于实验结果的准确性、科学性和可重复性提供可靠的保证。利用m tDNA分析小鼠的遗传稳定性已有报道,如用PCR-RFLP和PCR-SSCP技术对近交系小鼠、封闭群小鼠 mtDNA的 D-Loop、tRNA(Met+Glu+Ile)和ND3基因部分序列进行分析,结果发现品系内、品系间均表现出相同的酶图谱[15-16]。但是PCR-RFLP技术检测DNA多态性具有一定的局限性,所包含的遗传信息量较少,因此对于实验动物而言,PCR-RFLP技术不是一种理想的遗传检测手段。直接测序法能够直接分析DNA的变异,包含完整的遗传信息。管敏强(2009)[12]利用直接测序法测定封闭群ICR小鼠的m tDNA序列并分析其遗传稳定性。
线粒体DNA没有组蛋白保护和DNA修复系统,是所有的突变均能够保存下来,因此其突变频率比核DNA高的多,多态信息含量丰富。D-Loop区域是m tDNA中突变频率最高的区域,在人群和动物种群内均具有丰富的多态性[17-18]。作者曾首先对长爪沙鼠线粒体DNA控制区的DNA序列进行克隆、测序和生物信息学分析,发现长爪沙鼠D-Loop共包括997 bp,在啮齿类实验动物中与大鼠和小鼠具有较近的遗传关系[14]。为了分析我单位Z:ZCLA长爪沙鼠遗传稳定性,随机选择非同胞、非亲代的33只个体Z:ZCLA长爪沙鼠的m tDNA D-Loop区的序列多态性,没有发现序列突变,与前期报道的序列完全一致,推测Z:ZCLA长爪沙鼠群体内遗传变异频率较低,稳定性较高。
[1]方喜业,邢瑞昌,贺争鸣.实验动物质量控制[M].北京:中国标准出版社,2008:913-916.
[2]Hegsted DM,Gallagher A.Dietary fat and cholesterol and serum cholesterol in the gerbil[J].J Lipid Res,1967,8(3):210 -214.
[3]Tovar-palacio C,Potter SM,Hafermann JC,et al.Intake of soy protein and soy protein extracts influences lipid metabolism and hepatic gene expression in gerbils[J].JNutr,1998,128(5): 839-842.
[4]Forsythe WA 3 rd.Comparison of dietary casein or soy protein effects on plasma lipids and hormone concentrations in the gerbil (Meriones unguiculatus)[J].J Nutr,1986,116(7):1165 -1171.
[5]丁贤明,钱宝珍,Junichiro M,等.长爪沙鼠的遗传多样性分析[J].遗传,2008,30(7):877-884.
[6]聂金荣,聂朝晖.Z:ZCLA长爪沙鼠培育及其生物学特性研究[J].医学研究通讯,2004,33(6):27-28.
[7]徐其放,陈嘉昌,朱世杰,等.动物线粒体DNA的特异结构及应用分子系统学分析的方法[J].中国比较医学杂志,2005,15(5):315-319.
[8]廖顺尧,鲁成.动物线粒体基因组研究进展[J].生物化学与生物物理进展,2000,27(5):508-512.
[9]赵兴波,李宁,吴常信.动物线粒体核质基因互作的研究进展[J].遗传,2001,23(1):81-85.
[10]Nishibori M,Shimogiri T,Hayashi T,et a1.Molecular evidence for hybridization of species in the genus Gallus except for Gallus varius[J].Anim Genet,2005,36(5):367-375.
[11]Donne-GousséC,Laudet V,Hanni C.A molecular phylogeny of anseriformes based on mitochondrial DNA analysis[J].Mol Phylogenet Evol,2002,23(3):339-356.
[12]管敏强,曹琼洁,陈忠义,等.线粒体DNA序列分析封闭群小鼠遗传稳定性[J].实验动物与比较医学,2009,29(2): 113-116.
[13]张亚平,施立明.动物线粒体DNA多态性的研究概况[J].动物学研究,1992,13(3):189-295.
[14]李长龙,卢领群,郭红刚,等.长爪沙鼠线粒体DNA控制区全序列测定及分析[J].中国比较医学杂志,2009,20(4): 40-45.
[15]Dai JG,M in JX,Xiao YB,et a1.The absence of mitochondrial DNA diversity among common laboratory inbred mouse strains[J].JExp Biol,2005,208(23):4445-4450.
[16]戴纪刚,魏泓,肖颖彬.三个封闭群小鼠线粒体DNA PCRRFLP分析[J].第三军医大学学报,2000,22(1):29-32.
[17]冯强,席焕久,温有锋,等.那曲地区藏族m tDNA D环区高变区I序列多态性[J].解剖学杂志,2006,29(2):150-152.
[18]Liu RY,Yang GS,Lei CZ.The Genetic diversity ofm tDNA DLoop and the origin of Chinese goats[J].Acta Genetica Sinica,2006,33(5):420-428.
猜你喜欢
中华实用诊断与治疗杂志(2022年1期)2022-08-31
世界科学技术-中医药现代化(2022年3期)2022-08-22
意林·少年版(2021年13期)2021-08-30
海洋通报(2021年1期)2021-07-23
昆明医科大学学报(2021年3期)2021-07-22
生物学通报(2021年4期)2021-03-16
浙江医学(2018年16期)2018-09-08
第二课堂(小学版)(2017年11期)2017-11-27
中央民族大学学报(自然科学版)(2015年1期)2015-06-11
中国药业(2014年21期)2014-05-26
