
北沟林场天然次生林主要乔木树种种间关系分析1)
2012-07-02 00:04汲文宪陈丽华卞西陈
东北林业大学学报 2012年9期
汲文宪 陈丽华 郭 峰 卞西陈 王 鹏
(北京林业大学,北京,100083)
种间关联(物种联结性)是指不同物种在空间分布上的相互关联性[1],是对一定时期内植物群落组成物种之间相互关系的静态描述,通常是由于物种所处生境的不同而引起的。植物群落中的种间关系十分复杂,有竞争和排斥,也有互利和合作,了解群落中的种间关系,对于深入认识群落的结构、功能、动态以及分类等非常必要[2]。根据种间作用方式的不同,种间联结主要分为正关联、负关联和无关联(null association)。近年来,从山区到草原再到湿地[3-5],国内外学者对种间关系的研究方法和不同植被类型的优势种种间的联结性都有所研究。
我国华北地区典型地带性植被为暖温带落叶阔叶林,兼有温带针叶林分布。但由于早期大范围的人为干扰破坏,原生天然林越来越少,逐渐演替为杨、桦、椴、栎等混生次生林。探讨不同群落种间联结性的特征,对研究群落演替、森林经营管理以及植被恢复和重建具有重要的意义[6]。本文通过标准固定样地的调查,采用χ2检验、Ochiai指数、Pearson相关分析和Spearman秩相关分析法对河北省围场县北沟林场天然次生林主要乔木树种的种间联结特征进行了分析,并据此探讨了该区域植物群落中主要乔木树种之间的相互关系,以期对该地区植物群落特征及其动态变化的理解奠定基础,为森林健康经营模式的研究提供理论基础。
1 研究区概况
研究区位于滦河上游的河北省围场满族蒙古族自治县境内(41°47'~ 42°06'N,116°51'~ 117°45'E),位于阴山山脉、大兴安岭山脉余脉向西南延伸和燕山山脉的结合部,地质发展历史和地貌发育形成比较复杂,海拔高度为750~1 829 m,自然坡度为1/150~1/350。该区域属于寒温带向中温带过渡、半干旱向半湿润过渡、大陆性季风型高原山地气候,多年平均气温为5.0℃,无霜期为67~128 d。年降水量为380~560 mm,主要集中在7—9月份,占全年降水量的69%,年蒸发量为1 462.9~1 556.8 mm,平均相对湿度63%。该地区土壤主要以棕壤、褐土和灰色森林土为主。由于围场内地貌类型多样,降水充沛,气候多变,形成了丰富多样的植被资源。森林以天然次生林和人工林为主,主要乔木树种有华北落叶松(Larix principis-rupprechtii)、山杨(Populus davidiana)、白桦(Betula platyphylla)、油松(Pinus tabulaeformis)、蒙古栎(Quercus mongolica)、五角枫(Acer truncatum)等。
2 研究方法
2.1 样方设置和数据调查方法
采用样方法进行群落调查。样地设在山地深处,地理位置较偏僻,人为干扰程度较轻。样地海拔1 294 m,阳坡,坡度 19.5°,立地条件较好,郁闭度0.8。采用相邻网格法将样地分成100个20 m×20 m的样方网格,以网格为单元,调查记录固定样地的网格名、经纬度、坡向、坡位、坡度、海拔和地形地貌等,并记录树木的名称、坐标、树高、胸径、冠幅、枝下高、优势度、损伤状况、干型质量、病虫害和起源等。100个样方中共记录乔木树种14种,其中3种频率出现较低,低频率种类的种间联结性相应也较小,因此仅将频率出现大于5%的11个乔木树种(表1)列成11×100的数据矩阵,进行种间关联性和相关性分析的研究。

表1 北沟林场天然次生林乔木序号及种名
2.2 数据处理
计算每个样方物种的重要值,采用物种重要值综合指标来反映物种的特征。
重要值=(相对频度+相对显著度+相对密度)/3。式中:相对频度=(某物种的频度/样方内所有物种的频度之和)×100;相对显著度=(某物种的胸高断面积之和/样方内全部物种的胸高断面积之和)×100;相对密度=(某物种的个体数/全部物种的个体数总和)×100。
2.2.1 成对物种种间关联性分析
按物种在各样方内存在与否将s×n(s为种数,n为样方数)的数量数据矩阵转化为0,1形式的二元数据矩阵,然后分别构建种对间的2×2列联表,分别计算χ2值。利用χ2检验分别对这些联表进行检验,测定这些种对之间是否具有联结性。χ2检验的公式[7]为:

在下列两种情况下,χ2值被认为具有偏差:2×2列联表中任一小格期望值小于1;多于2个小格的期望值小于5。这种偏差用Yates连续校正系数进行纠正[8]。Yates系数纠正公式:
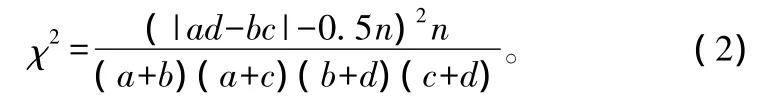
式中:n为样方数;a为两物种均出现的样方数;b、c为仅有1个物种出现的样方数;d是两物种均未出现的样方数。
当 ad-bc>0 时为正联结,当 ad-bc<0 时为负联结;当3.841≤χ2<6.635 时,表示种对间联结性显著(0.01<P<0.05),当 χ2>6.635 时,则表示种对间联结性极显著(P<0.01),否则为不显著。
2.2.2 种间关联程度的测定
选用测定两物种关联度较好的Ochiai指数计算种间联结度:

OI指数表示种对相伴随出现的机率和联结性程度,当指数等于0时,表示种间完全相异,无关联;当指数等于1时,物种间关联性最大。
2.2.3 Pearson相关分析和Spearman秩相关分析
采用11种物种的重要值作为数量指标进行Pearson相关分析和 Spearman秩相关分析[7],衡量两个物种间的关联程度。
Pearson相关系数是基于野外调查的连续性数据,要求原始数据服从正态分布,其表达式为:

Spearman秩相关系数是基于野外调查数据秩的大小,属于非参数检验,其表达式为:

式中:dk=(xik-xi)(xjk-xj);rp(i,j)和 rs(i,j)分别为物种i和j之间的Pearson相关系数和Spearman秩相关系数;xik和xjk分别是物种i和j在样方k中的重要值;¯xi和¯xj分别是物种i和j在所有样方中重要值的平均值。
3 结果与分析
3.1 χ2检验分析
通过χ2检验可知,北沟林场天然次生林群落中11种主要乔木树种组成的55组种对中,有8对物种联结性达到了显著(P<0.05)或极显著(P<0.01)水平,其中显著和极显著正关联7对,显著和极显著负关联1对,占乔木种对的14.5%,其余种对都处于不显著水平,此结果表明:种对间联结性较松散,存在相当的独立分布格局。呈正关联和负关联的种对分别为29对和26对,正负关系比为1.12,分别占乔木种对数的52.7%和47.3%。55组种对中正关联的稍多一点,说明次生林群落中乔木种群存在一定的差异,物种对资源的竞争程度或相互依赖程度不强,群落优势种处于稳定状态,对外界环境干扰有一定的抵抗力。
29组正联结的种对中,白桦与华北落叶松、华北落叶松与油松、蒙古栎与青杄、棘皮桦与油松、红桦与油松呈显著水平(P<0.05);五角枫与花楸、蒙古栎与棘皮桦呈极显著水平(P<0.01)。26组负联结的种对中仅有山杨与棘皮桦呈极显著水平(P<0.01)。刘喆和岳明[9]指出,种间正相关主要是由于它们的生态学特征和对生境的生态适应性相似及具有相互重叠的生态位所致;种间负相关,则主要是由于它们具有不同的生态学特征,对生境具有不同的生态适应性和相互分离的生态位所致。

表2 χ2检验、Pearson相关系数和Spearman秩相关系数检验结果
3.2 关联测度OI分析
χ2检验只能判断种间联结性显著与否,但这并不意味着那些检验不显著的种对之间不存在联结性,另外,χ2检验也不能区分联结强度的大小,因此有必要用OI值测定联结强度。由图2可知,关联度在0.2≤OI<0.4 的种对数最多,有24 对,占乔木种对数的43.6%。说明多数种对间的关联程度不紧密,这与χ2检验的结果相一致。关联度大于0.6的种对数较少,只有5对,占乔木种对数的9.1%,分别为:山杨与白桦、山杨与华北落叶松、山杨与五角枫、白桦与华北落叶松、白桦与五角枫。其中:山杨与白桦的OI值达到0.907,说明山杨和白桦的联结性最强,这与χ2检验结果中山杨和白桦为不显著水平不一致。说明χ2检验只能做一般的定性分析,若要精确分析研究对象,还需要与其他的分析方法结合。这与孙勃,张金屯[10]的研究结果一致。

图1 χ2检验半矩阵图

图2 Ochiai指数半矩阵图
3.3 Pearson 相关分析
由表2和图3可知,11种乔木树种组成的55组种对中表现为显著(P<0.05)和极显著(P<0.01)相关的种对数有12对,其中有8对表现为正相关,占乔木种对数的14.5%;有4对表现为负相关,占乔木种对数的7.3%。总体上仍是显著或极显著的联结性种对较少,而且呈显著和极显著正相关的种对数大于负相关种对数,这与χ2检验的结果一致。但是,χ2检验表现为显著和极显著的种对中,仅有6对与Pearson相关分析相符合,这是由于Pearson相关假定物种服从正态分布,而植被数据不能满足这一假设,所以结果会有偏差。
3.4 Spearman秩相关分析
Spearman秩相关系数属于非参数检验,它并不要求知道物种服从何种分布,因此应用起来更为灵活和方便。由图4可知,呈极显著和显著相关的种对有21对,占乔木种对数的38.2%。其中:16对呈正相关,5对呈负相关,几乎包含所有χ2检验中呈显著与极显著关联的种对;不显著相关的种对有34对,占乔木种对数的61.8%,且负相关种对数多于正相关。经Spearman秩相关分析的极显著相关种对数和显著相关种对数均要比χ2检验和Pearson相关分析的多,这是由于χ2检验只能做一般的定性分析,因此,在应用中要结合Ochiai关联度指数进行分析,得到的结果才能较为准确;Pearson相关分析要求物种服从正态分布,而自然界大多数物种服从集群分布[11],物种数据符合正态分布的要求限制了Pearson相关指数的准确性,因此Spearman秩相关指数的分析结果灵敏度最高,这与以前的研究结果[3,12]是一致的。

图3 Pearson检验半矩阵图

图4 Spearman秩相关系数半矩阵图
3.5 生态种组的划分
群落中生态习性相似的种可以划分为一个生态种组[13],而群落内的种间联结性和相关性揭示了群落中不同种类因受小生境因子影响而体现在空间分布上的相互关系[14]。生态种组的划分有助于人们正确地认识和研究群落的结构、功能以及发展和演替。依照χ2检验、Ochiai关联指数、Pearson相关分析和Spearman秩相关分析结果,结合植物种群的生物学和生态适应特征,将北沟林场天然次生林群落中11种乔木树种划分为3个生态种组。
第一生态组为:种1(山杨)、种2(白桦)、种6(花楸)、种8(红桦)和种11(青杄),这些树种为向阳性植物,不耐阴,有一定的耐旱性,多生长在干燥的阳坡;第二生态组为:种4(五角枫)、种9(油松)和种10(硕桦),这些树种不喜光,耐寒,喜温凉湿润气候,多生长在阴坡或半阴坡;第三生态组为:种3(华北落叶松)、种5(蒙古栎)和种7(棘皮桦),这些树种喜光,耐寒,喜凉爽湿润气候,多生长在向阳山坡。
4 讨论
群落在长期的演替过程中,由于种内种间的竞争,群落的组成成分逐渐趋于稳定。由于生态学特性的差异,各个物种都占据有利于自己的位置,和谐共处,相互竞争大为减弱,所以多数种对联结程度不强,关系松散,独立性较强[15]。本文运用4种联结指数对北沟林场天然次生林群落中11种乔木树种进行分析的结果,一方面表现出种对间的联结关系绝大多数未达到显著程度,种对间的联结较松散、独立性相对较强,这与北沟林场天然次生林群落目前的发展阶段及物种本身的生态学特性有关,目前大多数群落正处于较稳定的顶级阶段。另一方面,不同检验方法对种对间的检验结果表现出种对间的正负关联比略大于1,说明仍具有明显的次生性或正处于演替中期阶段,多数物种间关系不紧密,未形成明显的总体联结性,群落处于相对稳定状态。随着演替的进行,负联结比例将随演替进程下降,群落结构和种类组成将逐渐趋于完善和稳定,种间关系也将趋于正关联,以求得多物种间的稳定共存[13,16]。
种对间呈正关联,主要是由于它们具有相近的生物学特性,对生境具有相似的生态适应性和相互重叠的生态位所致;种对间的负关联,则主要是由于它们具有不同的生物学特性,对生境具有不同的生态适应性和相互分离的生态位所致[17]。在相同的生长环境和竞争条件下,对生境要求相同或相似的种对在适合其生存的生境中表现出显著的正关联,对生境的要求及资源利用有差异,必然会导致种间对资源的利用性竞争,种对往往会表现出显著的负关联,这在前人的研究中已经得到了验证[15,18-20],本研究结果进一步证实了这一规律。本研究中,耐旱不耐阴的向阳植物山杨和白桦以及花楸和青杄,耐寒,喜温凉湿润气候的五角枫和硕桦以及油松和硕桦,耐寒喜光向阳的华北落叶松和蒙古栎以及蒙古栎和棘皮桦都表现出显著正关联;生长在干燥阳坡的山杨和生长在阴坡喜温凉湿润气候的五角枫则表现为显著负关联。其它物种间呈显著正相关或是显著负相关也都是由于两物种的生物学特性相似或不同所致。当然,还有很多因素会影响种对之间的关系,这其中的具体原因还有待于进一步研究。
种间连结和关联的计算是研究种间关系的重要方法,但是利用种间关联指数进行种间关系的研究也存在着一些缺陷:①测定结果受样方面积大小和样方数量多少的影响[6]。种间关联研究中应该用一个什么指标来衡量样地大小对种间关联所带来的影响;样方选取的科学性以及数量,这些都是今后研究中应该努力解决的问题,因为种间关系的研究更关注的是物种的空间布局,而不仅仅是种类和数量。②仅能测定群落中两个物种间的关系,而群落中种群间的关系不局限于两两之间[21]。为了全面检测多物种之间的关系需借助于多物种间联结,多物种间联结可检测3,4,…,n个物种间的相互关系[22]。③种间关联指数所计算的结果只是物种现有的状态,它可能是群落逐步演替的结果,也可能是偶然的一些原因造成的,因此要对种间关系有更全面和深刻的研究,就必须对一个区域的物种进行动态的定点观测和研究,并需要借助其它生态学方法,这将有助于揭示种间联结的内在机制。
5 结论
采用χ2检验、Ochiai关联指数、Pearson相关分析和Spearman秩相关分析对北沟林场天然次生林11种乔木的种间联结和相关性的研究结果表明:1)群落中物种间的总体关联性较弱,多数种对间关联程度未达到显著水平,说明林场多数乔木树种间关系不紧密,未形成明显的总体联结性,群落处于相对稳定状态;2)利用χ2检验、Ochiai关联度指数、Pearson相关指数和Spearman秩相关指数计算种间关联和联结得到的结果有很大的相似性,但是,Pearson相关检验在本研究中有较大偏差,Spearman秩相关检验的灵敏度要大于χ2检验和Pearson相关检验;3)依照χ2检验、Ochiai关联度指数、Pearson相关分析和Spearman秩相关分析结果,结合植物种群的生物学和生态适应特征,将北沟林场天然次生林群落中11种乔木树种划分为3个生态种组。
[1]Greig-Smith P.Quantitative Plant Ecology[M].3rd ed.Oxford:Blackwell scientific publications,1983:23-245.
[2]尹林克,李涛.塔里木河中下游地区荒漠河岸林群落种间关系分析[J].植物生态学报,2005,29(2):226-234.
[3]张丽霞,张峰,上官铁梁.芦芽山植物群落种间关系的研究[J].西北植物学报,2001,21(6):1085-1091.
[4]王正文,祝廷成.松嫩草原主要草本植物种间关系及其对水淹干扰的响应[J].应用生态学报,2003,14(6):892-896.
[5]吴东丽,上官铁梁,薛红喜,等.滹沱河湿地植物群落的种间关系研究[J].山西大学学报:自然科学版,2003,26(1):71-75.
[6]胡理乐,闫伯前,刘琪琮.南方丘陵人工林林下植物种间关系分析[J].应用生态学报,2005,16(11):2019-2024.
[7]张金屯.植被数量生态学方法[M].北京:中国科学技术出版社,1995:79-87.
[8]张震云,周伶,上官铁梁,等.半干旱区锦鸡儿群落优势种种间关系[J].生态学杂志,2011,30(9):1868-1874.
[9]刘喆,岳明.太白山独叶草及其伴生种的种间关联研究[J].武汉植物学研究,2007,25(5):445-450.
[10]孙勃,张金屯.天龙山木本群落种间关系的研究[J].西北植物学报,2004,24(8):1457-1461.
[11]Legendre L,Legendre P.Numerical Ecology[M].Amsterdam:Elsevier Scientific Publishing Company,1998:139-144,194-198,216-222.
[12]李晶,上官铁梁.山西北部沙棘群落优势种种间关系[J].山地学报,2006,24(2):222-227.
[13]Mueller-Dombois D,Ellenberg H.植被生态学的目的与方法[M].鲍显诚,译.北京:科学出版社,1986.
[14]周先叶,王伯荪,李鸣光,等.广东黑石顶自然保护区森林次生演替过程中群落的种间联结性分析[J].植物生态学报,2000,24(3):332-339.
[15]邓贤兰,刘玉成,吴杨.井冈山自然保护区栲属群落优势种群的种间联结关系研究[J].植物生态学报,2003,27(4):531-536.
[16]杜道林,刘玉成,李睿.缙云山亚热带栲树林优势种群间联结性研究[J].植物生态学报,1995,19(2):149-157.
[17]王琳,张金屯.历山山地草甸优势种的种间关联和相关分析[J].西北植物学报,2004,24(8):1435-1440.
[18]赵则海,祖元刚,杨逢建,等.东灵山辽东栎林木本植物种间联结取样技术的研究[J].植物生态学报,2003,27(3):396-403.
[19]王乃江,张文辉,陆元昌,等.陕西子午岭森林植物群落种间联结性[J].生态学报,2010,30(1):67-78.
[20]邓福英,臧润国.海南岛热带山地雨林天然次生林的功能群划分[J].生态学报,2007,27(8):3240-3249.
[21]胡理乐,李新,江明海,等.宣恩七姊妹山珙桐群落种间联结分析[J].武汉植物学研究,2003,21(3):203-208.
[22]Dale M R T.Spatial pattern analysis in plant ecology[M].New-York:Cambridge University Press,1999:147-167.
猜你喜欢
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
建筑与预算(2022年6期)2022-07-08
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
农村实用技术(2020年1期)2020-12-16
河南科学(2020年3期)2020-06-02
蔬菜(2018年5期)2018-05-17
现代园艺(2018年14期)2018-01-18
杂草学报(2015年2期)2016-01-04
草业科学(2015年9期)2015-12-11
