
宝天曼固定样地林窗特征及与林窗边缘木的相关性研究
2012-07-14 02:49庄静静刘晓静叶永忠朱学灵
河南农业大学学报 2012年5期
庄静静,刘晓静,叶永忠,朱学灵,,3
(1.河南农业大学生命科学学院,河南郑州450002;2.国家林业局宝天曼森林生态定位研究站,河南内乡474350;3.河南宝天曼自然保护区管理局,河南内乡474350)
林窗有不同的定义,这是由于不同学者从各自的研究角度采取不同尺度而造成的[1].但总体来看,其内涵基本一致,即:森林群落中林冠层乔木的死亡必然导致森林中相对连续的林冠层面出现间断的现象,这种空间上的不连续现象,即为林窗.英国生态学家 WATT[1,2]首先提出林窗(gap) 的概念,RUNKLE[3]和 BARDEN[4]也通过对同一森林群落的进一步研究指出:“林窗就是丛林中单株树、树的某一部分或多株树死亡所形成的林冠空隙”[1,2,5].RUNKLE[3]深入研究后,对林窗的概念进行了扩充,将林窗分为狭义的林窗和广义的林窗[1~3,6].而 WATT 认为林窗主要是指森林群落中老龄树死亡或因偶然性因素(如干旱、台风、火灾等)导致成熟阶段优势树种的死亡,从而在林冠层造成空隙的现象[7,8].充分认识林窗基本特征,有助于揭示林窗对森林群落组成、结构和动态的影响及其维持机制[9~11].林窗的形成和变化构成了森林景观的流动镶嵌结构,对植被的正常更新具有重要作用,它是森林群落中经常发生的重要的小规模干扰[1,2],也是推动森林植被演替和更新的重要驱动力[8],已成为当前森林生态学研究的最活跃领域之一.张希彪等[12]对黄土丘陵区油松天然次生林林窗特征与更新动态进行了研究,王家华等[2]对林窗研究进展进行了详细的阐述.刘庆等[9]研究了玉龙雪山自然保护区丽江云杉林林窗特征.兰国玉等[13]分析研究了秦岭华山松种群格局规模与林窗特征.包维楷等[14]对瓦屋山中亚热带湿性常绿阔叶林的林窗形成特征进行研究.但目前并未有北亚热带向暖温带过渡地区林窗特征的相关研究报道,本研究通过对宝天曼固定样地内的林窗特征的调查,研究了保护区内各林窗的形状、大小等,以及林窗形成木、边缘木等其它相关因子,探讨其在中国第2级地貌阶梯向第3级地貌阶梯过渡的边缘林窗特征.
1 研究区概况
宝天曼自然保护区位于南阳市内乡县辖区内,属于河南省西南部,秦岭东段、伏牛山南坡[15],地理位置为北纬 33°25'~33°33',东经 111°53'~112°,总面积为 5 412.5 hm2[16].保护区地理位置独特,属于典型的北亚热带向暖温带过渡地区[17],保护区内最高峰海拔1 830 m,地形复杂,土层深厚,本区岩石主要是花岗岩,有少量的石灰岩和沙岩[18].该区为季风型大陆性气候[19],四季分明,全年降雨量为900 mm,多分布于6~8月份,年均相对湿度68%[20].研究地内可分为乔木层、灌木层及草本层,乔木层以锐齿槲栎(Quercus aliena)、华山松(Pinus armandi)、葛罗槭(Acer grosseri)、三桠乌药(Lindera obtusiloba)、膀胱果(Staphylea holocarpa)为优势群落类型,群落高15~20 m,盖度约90%[21];灌木层主要以海州常山(Clerodendrum trichotomum)、中华绣线菊(Spiraea chinensis)、接骨木(Sambucus williamsii)等组成为主,高度10 m,盖度达40%;草本层稀疏,高度20~60 cm,盖度为15% ~30%.
2 研究方法
2.1 林窗调查
调查于2011-07月进行.在大面积踏查的基础上,采用样方调查法[1,9,10],在林内西北点作为调查起点,沿罗盘仪所指方向,由北向南穿行,遇到林窗则停下来,调查林窗的长和宽、形状、方向,记录林窗内伐根的数量和种类,记录距林窗边缘周围3 m内立木的种类、树高、胸径等.调查项目[22]主要包括:海拔、坡度、坡向等生态因子;林窗的形状、大小、形成方式、边缘木的种类、株数,林窗长轴与短轴的长度;记录林窗形成木的形成方式并分枯立、干基折断、干中折断、人为活动、折枝、砍伐和掘根风倒.
2.2 数据处理
林窗面积大小一般采用椭圆形面积公式进行计算,椭圆形公式计算:S= πLW/4[3,23],式中:S 为林窗面积,L为林窗长度,W为林窗宽度,它指与林窗长度相垂直的最大直径[24].
3 结果分析
3.1 林窗形状分布特征
一般来说,从水平面上来看,林窗的形状是不规则的,有圆形、椭圆形、多边形、浑圆形等[3~5].但总体来说,其形状近似于椭圆形,从垂直剖面看,林窗像一个倒圆锥体[3,5,12,25],越往上开敞度越大,向下则逐渐变小,如果考虑到更新树种,则林窗的垂直剖面近似于鼓形.根据调查数据可知,样地内的林窗形状从水平面上来看,主要以多边形为主,占样地内林窗形状的83.5%.
3.2 林窗大小分布特征
林窗大小是表征林窗内生态环境和特征的重要指标之一[2,6,12,26,27],通常用其面积来衡量,它不仅反映了林窗内微环境因子的变动,而且提供了林窗内更新种类所能利用的空间资源.研究者一般把林窗范围确定为 4 ~ 1 000 m2[2,6,28,29],因小于 4 m2的间隙很难与林分中的枝叶间隙区分开,而大于1 000 m2的范围则一般当作林中空地处理.本研究样地内林冠林窗(CG)和扩展林窗(EG)的面积分别划分为4个等级,统计82个林窗在不同等级中的数量分布(表1)。从中表1看出CG最大面积的林窗为 49.46 m2,最小面积的为 4.71 m2,大于45 m2的林窗只有3个,小于45 m2的林窗共33个,其中 <15 m2的林窗数量最多,共有19个,占样地林窗总数的53%,其中以<15 m2的数量比最大,而30~40 m2的面积比最大,分别为53%和29%;EG最大面积的林窗为223.73 m2,最小面积的为85.76 m2,大于200 m2的林窗有6个,小于100 m2的林窗有10个,其中100~150 m2的林窗数量最多,占样地内林窗总数的48%,101~150 m2的数量比和面积比最大,分别为48%和42%.CG随着面积的变小,林窗数不断的增加,说明了样地内林窗的面积较小,并无大林窗,这不仅与保护区内林窗形成原因有重要关联,也与形成林窗的物种有重要关系;而EG在>200 m2时开始下降,充分说明了样地内大林窗数量少,小林窗数量多的特征.
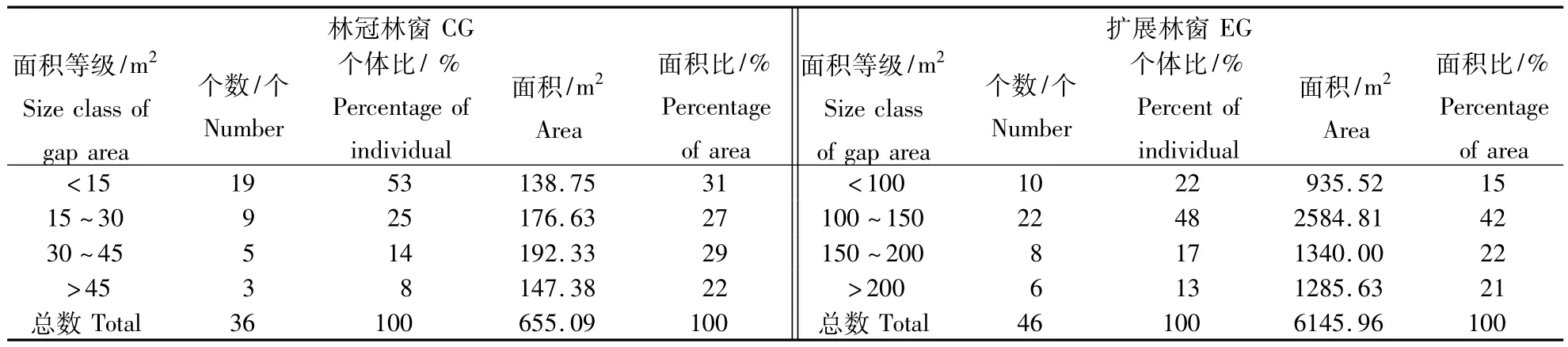
表1 样地内EG和CG大小结构Table 1 Size structure of gaps in plot
3.3 林窗形成木(GM)的数量及种类
林窗形成木(GM)的特征直接或间接地影响着林窗的各种特征和树种的更新[9,30].林窗形成木是创建林冠空隙的主林层树木,当主林层达到一定高度和年龄后,在外界因子的作用下死亡,形成林窗.从林窗内死亡木情况来看(表2),林窗形成木的种类有8种55株,主要组成树种有锐齿槲栎(Quercus aliena)36株(65.5%),所占比例最大,华山松(Pinus armandi)12 株(21.8%)、葛罗槭(Acer grosseri)2 株(3.63%)、崖樱桃(Cerasus scopulorum)1 株(1.82%)、三桠乌药(Lindera obtusiloba)1 株(1.82%)、马鞍树(Maackia hupehensis)1株(1.82%)、膀胱果(Staphylea holocarpa)1株(1.82%),灌木有四照花(Dendrobenthamia japonica)1 株(1.82%).
在不同森林中,形成林窗的形成木也是不同的,可能是一株形成木,也可能是多株形成木.在调查的样地内,每个林窗内的形成木数量都不相同,有些林窗是由1株形成木形成,而有的林窗由2株或多株形成木形成.统计结果表明(表2),林窗内林窗形成木的数量最多的有12株,最少的仅1株;1 株树形成的林窗占 57.1%;4,5,7,8,11,12 株形成木形成的各占7.1%,林窗形成木的数量较少是样地内林窗面积较小的主要原因.

表2 样地内林窗形成木的数量Table 2 The number of the gap formation wood in plot 个
3.4 林窗特征与林窗形成木的关系
样地林窗的形成方式主要有枯木、倒木、林中平地、灌木群以及岩石区.由于样地内林分多为成熟林或过熟林,树干心腐脆弱,易发生风倒、折干死亡[9],这成为林窗形成的主要原因,占78.57%;而林内状况和林内地理位置分别占4.76%和16.67%,表明外部环境和自身生理因素所形成的林窗形成木造成了大部分林窗,与人为因素联系较小.
在样地内锐齿槲栎(Quercus aliena)作为优势种,该群落层次较高,受冬季冰雪和强劲的风力的影响,使一些受病虫害侵扰和衰老的乔木发生根倒,从而形成林窗,一般主林层树木掘根风倒形成的林窗面积较大[31],从表3可知,在55株林窗形成木中,倒木与枯立木形成方式分别占60%和40%,随着林窗面积的减少,枯木的数量越来越多,所以倒木与枯立木是该群落林窗形成的重要方式,而枯立木形成的林隙一般较小。落叶松更能适应冬季雪压和冰挂的不利环境,降低机械损伤,而小乔木层因为树体矮小受损伤的机率更小,所以样地内小林窗多而大林窗较少.

表3 样地内林窗形成木的特征Table 3 The characteristics of the gap formation wood in plot
3.5 林窗边缘木的特征
3.5.1 林窗边缘木的数量及种类
林窗边缘木是指包围整个林窗的树木,它是衡量林窗特征的一个重要指标[32].以详细调查的林窗作为总体,统计其边缘木的数量及其比例(表4).形成林窗的林窗边缘木主要树种是锐齿槲栎(Quercus aliena)共170株,其次是华山松(Pinus armandi)51株,还有马鞍树(Maackia hupehensis)、千金榆(Carpinus cordata)、葛罗槭(Acer grosseri)、毛棶(Cornus walteri)、三桠乌药(Lindera obtusiloba)、膀胱果(Staphylea holocarpa)、崖樱桃(Cerasus scopulorum)、地锦槭 (Acer mono)、山樱桃(Cerasus cyclamina)、铁木(Ostrya japonica)、四照花(Dendrobenthamia japonica)、灯台树(Bothrocaryum controversum)等共14种植物,其中锐齿槲栎(Quercus aliena)在林窗边缘木中占的比例最大,占所有树种的53.8%,而将近10种只是出现不超过10株,所占的比例很少.
3.5.2 林窗边缘木的径级结构
林窗边缘木的径级结构是指林窗边缘木胸径的径级分布,是森林群落研究中最基本的林分结构[33].样地内林隙边缘木主要树种是锐齿槲栎(Quercus aliena),其胸径范围为20~50 cm.将本次调查的316株边缘木,按照胸径划分为8个径级(表5).由表5可以看出,胸径30~40 cm所占的比例最大,为105株,占总体的33.23%;胸径>70 cm的个体只有2株,占总体的0.63%.
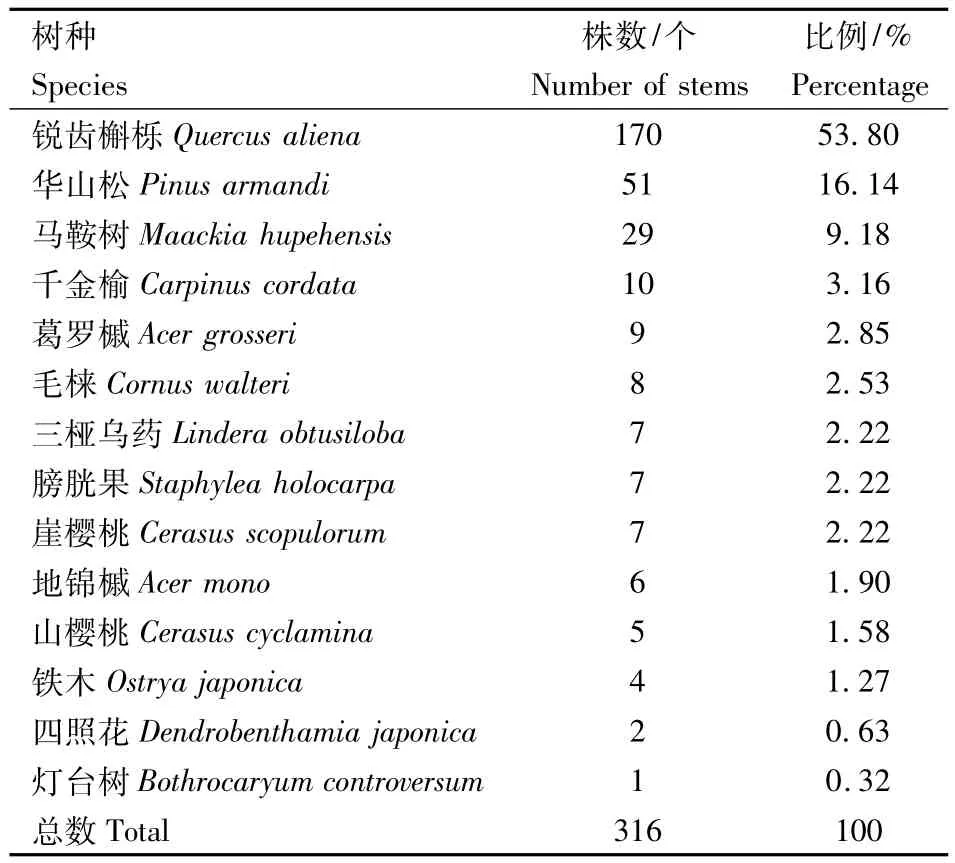
表4 林窗边缘木的组成结构Table 4 species composition structure of gap makers in forest
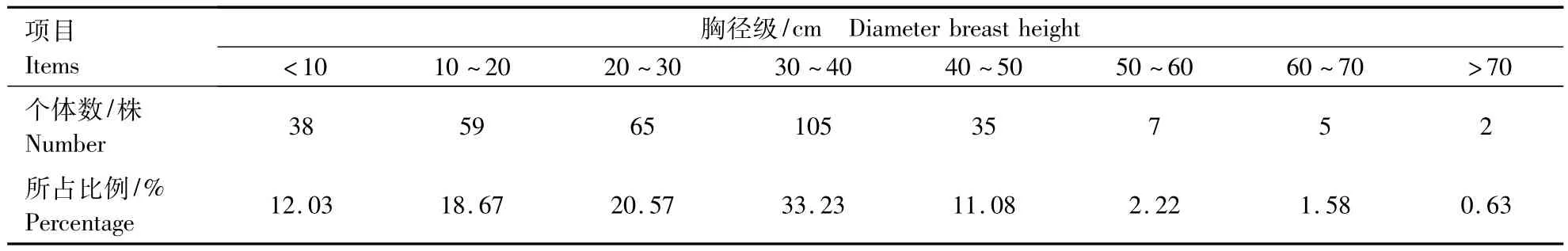
表5 林窗边缘木不同胸径级的个体数Table 5 The number of the different DBH in plot
3.6 林下及林隙内的乔木更新
从样地调查可以看出,整个样地中以锐齿槲栎(Quercus aliena)为主要优势种,其次是华山松(Pinus armandi),故选取这2个优势种来了解林下及林隙内的乔木幼苗更新情况.从图1可以看出,在林下锐齿槲栎(Quercus aliena)的萌发的幼苗与华山松(Pinus armandi)萌发的幼苗量多,而幼树数量却与幼苗数量呈负相关,这是因为锐齿槲栎(Quercus aliena)虽说在整个样地内作为优势种,种子库含量丰富,每年产生的种子数量多,又因自身种子生活力高的特性[34],故在林下萌发的幼苗数量较多,但在长期的演替过程中,由于环境或自身竞争的压力,使发育成为幼树的锐齿槲栎(Quercus aliena)幼苗较少;而作为第2位优势种的华山松(Pinus armandi)因幼苗属于耐荫性树种[35],除了可以在林下较好的生长外,华山松(Pinus armandi)因种子大,数量少,加之动物的取食,使其种内竞争减少,也是成为华山松幼树较多的原因之一;在林窗内锐齿槲栎(Quercus aliena)萌发的幼苗数量比在林下萌发的更多,这是因为锐齿槲栎(Quercus aliena)是非耐荫性树种,在林窗形成后,充足的光照条件,使锐齿槲栎(Quercus aliena)幼苗在林窗内快速萌发,华山松种子主要靠动物的传播,因林窗的形成影响动物的活动,从而华山松的幼苗量减少,而且在林隙内因为环境的适宜使锐齿槲栎(Quercus aliena)幼苗加速生长,发育成幼树来占领上层空间从而形成优势地位.但与形成的幼苗数量相比,生长成为幼树的锐齿槲栎(Quercus aliena)幼苗还是有限的,这可能与锐齿槲栎(Quercus aliena)种内竞争以及密度制约有关.
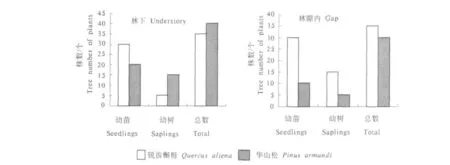
图1 林下、林隙内乔木幼苗株数Fig.1 Number of tree regeneration in understory and gap
4 结论与讨论
经调查,样地内的林窗形状从水平面上来看,由于样地内地形以及林窗形成木的形成方式不同,主要是以多边形为主,林窗大小分布表现出负指数分布,即小林窗多、大林窗少的特点[14].样地内共调查林窗数82个(CG和GE),CG林窗面积为4.71 ~49.46 m2不等,大林窗(面积 >45 m2)、中等林窗(面积15~45 m2)和小林窗(面积 <15 m2),各占8.33%、38.89% 和 52.78%.GE 林窗面积为85.76 ~223.73 m2,大林窗(面积 >200 m2)、中等林窗(面积100~200 m2)和小林窗(面积<100 m2),各占22%、65%和13%.样地中形成林窗最重要的方式是枯木和倒木,其次是林中平地.林窗内林窗形成木的数量最多的有12株,最少的仅1 株;1 株树形成的林窗占 57.1%;4,5,7,8,11,12株形成木形成的各占7.1%.样地内林窗形成木的种类共有8种55株,主要组成树种有锐齿槲栎(Quercus aliena)36 株(65.5%),所占比例最大,华山松(Pinus armandi)12株(21.8%)、葛罗槭(Acer grosseri)2 株(3.63%)、崖樱桃(Cerasus scopulorum)1 株(1.82%)、三桠乌药(Lindera obtusiloba)1株(1.82%)、马鞍树(Maackia hupehensis)1 株(1.82%)、膀 胱 果 (Staphylea holocarpa)1 株(1.82%),灌木有四照花(Dendrobenthamia japonica)1株(1.82%).林窗形成一般是由自然干扰和人为干扰两方面因子促成林窗的形成,主要有气候变化、土壤发育状况和地形特征等外因以及树木年龄和根系发育等内因。本研究发现样地林窗多是因为树木自身成熟林引起的倒木或枯木,由于宝天曼是自然保护区,人为干扰因素不多,即使偶然发生的火灾或人工砍伐,其面积也非常小,所以大多数林窗是由暴风雨、雷击大树、自身衰老等原因形成.
边缘木的特性影响林窗内物种的更新、外来物种的传播和环境因子的变化[32],因此本研究以10 cm为径级单位统计各径级的边缘木数量。可以看出在样地内锐齿槲栎(Quercus aliena)占相对优势的位置,林窗形成木以锐齿槲栎(Quercus aliena)所占的比例最大,且在统计过程中林窗边缘木也以锐齿槲栎(Quercus aliena)为主,胸径主要介于20~50 cm,其中30~40 cm最多,这是形成林窗最可能的阶段。在林窗形成初期,由于林窗内光照条件的变化,促使喜光物种萌发生长,使林窗内物种丰富度和多度增加,且林窗边缘物种的种类也发生了改变.作为样地先锋树种的锐齿槲栎(Quercus aliena),因为其种子产量大、活性高的特性,其幼苗更新大多数是以实生苗进行的[21],但其从幼苗到幼树的成活率极低,所以幼树极罕见,但是在林窗等较为开阔的地方,光照充足,其幼苗能迅速生长,并占领上层空间成为优势种.但从整个样地来看,锐齿槲栎(Quercus aliena)幼苗的更新还是面临着困难,除了环境和动物的影响外,还存在种内竞争的压力,使其更新艰难.华山松(Pinus armandi)在幼龄时处于优势状态,它种子较大,主要靠动物传播,林窗的形成影响了动物的活动,使种子传播困难,加上动物的取食,使华山松的幼苗数极速下降,但其形成幼树的成活率较高,使样地内华山松(Pinus armandi)幼树较多,但因针叶类树种与阔叶类树种相比一般处于劣势地位,特别是在中龄林中,这种现象更加明显.随着时间的演替,样地中的锐齿槲栎(Quercus aliena)还会处于优势地位,而华山松(Pinus armandi)的劣势地位会越来越明显.
[1] 张远彬,王开运.SEPPOKELLOMA K.针叶林林窗研究进展[J].世界科技研究与发展,2003,25(5):69 -74.
[2] 王家华,李建东.林窗研究进展[J].世界林业研究,2006,19(3):27 -30.
[3] RUNKLE J R.Patterns of disturbance in some old -growth mesic forests of eastern North America[J].Ecology,1982,63(5):1533 -1546.
[4] BROKAW N V L.The definition of treefall gap and its effect on measures of forest dynamics[J].Biotropica,1982,14(2):158 -160
[5] RUNKLE J R.Gap regeneration in some old - growth forests of the eastern united states[J].Ecology,1981,62(4):1041-1051.
[6] RUNKLE J R.Synchrony of regeneration,gaps,and latitudinal differences in tree species diversity[J].Ecology,1989,70(3):546 -547
[7] 周丹卉,贺红士,孙国臣,等.林窗模型及其在全球气候变化研究中的应用[J].生态学杂志,2007,26(7):1303-1310.
[8] 张远彬,王开运,鲜骏仁.岷江冷杉林林窗小气候及其对不同龄级岷江冷杉幼苗生长的影响[J].植物生态学报,2006,30(5):941 -946:.
[9] 刘 庆,吴 彦,吴 宁.玉龙雪山自然保护区丽江云杉林林窗特征研究[J].应用生态学报,2003,14(3):845-848.
[10] RUNNKLE J R,YETTER T C.Treefalls revisted:Gap dynamics in the southern Appalachians[J].Ecology,1987,68(2):417 -424
[11] SHUGART H H.A Theory of forest dynamics:the ecological implications of forest succession models[M].New York:Springer Verlag,1984
[12] 张希彪,王瑞娟,周天林,等.黄土丘陵区油松天然次生林林窗特征与更新动态[J].应用生态学报,2008,19(5)2103-2108.
[13] 兰国玉,雷瑞德,安 锋,等.秦岭华山松种群格局规模与林窗特征[J].生态学杂志,2006,25(4):652 -656.
[14] 包维楷,刘照光,袁亚夫,等.瓦屋山中亚热带湿性常绿阔叶林的林窗形成特征[J].应用生态学报,2001,12(5):485-490.
[15] 史作民,刘世荣.河南宝天曼种子植物区系特征[J].西北植物学报,1996,16(6):329 -335.
[16] 王文静,何雅蔷.宝天曼自然保护区青檀林结构特征与物种多样性研究[J].河南农业大学学报,2001,35(3):364-367.
[17] 史作民,程瑞梅,刘世荣,等.河南宝天曼药用植物资源及其保护对策[J].林业科学研究,2005,18(3):195-198.
[18] 史作民,刘世荣,程瑞梅.宝天曼地区栓皮栎林恢复过程中高等植物物种多样性变化[J].植物生态学报,1998,22(6):415 -421.
[19]闫东锋,何 静,郭 芳,等.宝天曼栎类天然次生林乔木层种间联结性研究[J].西北林学院学报,2009,24(4):26-30.
[20] 史作民,程瑞梅,刘世荣.宝天曼落叶阔叶林种群生态位特征[J].应用生态学报,1999,10(4):265 -269.
[21] 袁志良,庄静静,朱学灵,等.宝天曼锐齿槲栎-华山松林样地物种组成及空间分布格局[J].河南农业大学学报,2011,45(12):411 -418.
[22] 周 东,刘国彬.林窗对子午岭天然辽东栎群落林下植物多样性的影响[J].中国农学通报,2010,26(7):91-98.
[23] PLATT W J ,STRONG D R.Special feature:Gaps in canopy ecology[J].Ecology,1980,70(3):535 -546.
[24] DYNESIUS M,JOHNSON B G.Dating uprooted trees:Comparison and application of eight methods in a boreal forest[J].Canadian Journal of Forest Research,1991,21(5):655-665.
[25] LAWTON R O,PUTZ F E.Natural disturbance and gapphase regeneration in a wind-exposed tropical cloud forest[J].Ecology,1988,69(3):764 -777.
[26] STOEHR M U.Seed production of western larch in seedtree systems in the southern interior of British Columbia[J].Forest Ecology and Management,2000,130(1/3):7-15.
[27] 李永兵,王 辉,付 菁.子午岭辽东栎林林窗主要树种更新生态位[J].生态学杂志,2008,27(4):2062 -2066.
[28] 臧润国,刘艳静,董大方.林隙动态与森林生物多样性[M].北京:中国林业出版社,1999.
[29] TYRRELL L E,CROW T R.Structural characteristics of old-growth hemlock-hardwood forests in relation to age[J].Ecology,1994,75(2):370 - 386.
[30] WANG J X,XU J D,Hou G W,et al.Ecology and regeneration of cutted blank in alpine and plateau region of the upper reach of Yangtze River[M].Beijing:China Forestry Press,1995.
[31] 臧润国.林隙更新动态研究进展[J].生态学杂志,1998,17(8):50 -58.
[32] 谭 辉,朱教君,康宏樟,等.林窗干扰研究[J].生态学杂志,2207,26(4):587 -594.
[33] 杨晓伟,洪 伟,吴承祯.中亚热带台湾相思次生林林隙边缘木高径分布特征[J].西南林学院学报,2010,20(6):25 -28.
[34] 吴明作,刘玉萃,姜志林.栓皮栎种群生殖生态与稳定性机制研究[J].生态学报,2001,21(2):225 -230.
[35] 陈建平,李银梅.小陇山林区辽东栎和华山松天然更新规律初探[J].甘肃科技,2009(10):143-144.
猜你喜欢
林业资源管理(2022年6期)2023-01-18
科海故事博览(2022年28期)2022-11-25
现代园艺(2021年23期)2021-12-01
林业勘查设计(2020年1期)2021-01-18
新农业(2020年18期)2021-01-07
中国森林病虫(2020年1期)2020-01-18
新闻传播(2018年15期)2018-09-18
通信产业报(2016年44期)2017-03-13
河南科技(2014年24期)2014-02-27
雕塑(1999年2期)1999-06-28
