
甜瓜重组自交系群体第1雌花开花期遗传分析
2012-08-08 01:23高美玲栾非时朱子成
中国蔬菜 2012年22期
高美玲栾非时朱子成
(1齐齐哈尔大学生命科学与农林学院,齐齐哈尔大学遗传工程重点实验室,黑龙江齐齐哈尔161006;2东北农业大学园艺学院,黑龙江哈尔滨 150030)
甜瓜(Cucumis meloL.)是世界十大水果之一,早熟性是甜瓜育种的重要目标之一,早熟甜瓜可以提高经济效益,以往主要用开花到果实成熟的天数代表早熟性,但第 1雌花开花期(DFF)也是早熟性状构成因素之一,对其深入研究较少。Tyagi等(2003)研究表明,丝瓜第1雌花开放时间是由非加性基因效应决定的。Monforte等(2004)用从定植到收获的天数代表甜瓜早熟性(EA),利用 F2和 DH群体研究了早熟性的遗传并进行了 QTL定位,在两个群体中EA表现为连续变异,认为是数量性状遗传,并检测到了9个贡献率大于10%的QTL位点,但在不同的试验中QTL检测的重复性较低。利用重组自交系(RIL)群体进行遗传分析,可进行不同环境的重复试验,本试验以重组自交系群体为材料,在不同季节采用 RIL群体主基因+多基因混合遗传模型分析法(盖钧镒 等,2003),研究第1雌花开花期的遗传特点和不同季节第1雌花开花期的遗传稳定性,为提高甜瓜早熟育种效率和充分利用甜瓜早熟资源提供理论依据。
1 材料与方法
1.1 试验材料及重组自交系群体的构建
以美国威斯康星大学瓜类分子育种实验室提供的纯雌性早花厚皮甜瓜品系WI998为母本,东北农业大学园艺学院西甜瓜分子育种研究室特有的雌雄异花同株晚花薄皮甜瓜品系3-2-2为父本,父母本分枝、结果等习性差异显著,具体参见Luan等(2010)的研究结果。
于2007年3~6月配制F1种子,2007年7~10月从F1的1个瓜中取出300粒种子进行单粒传,自交得到F2,2008年3~6月自交得到F3,2009年2~8月自交得到F4,2009年10月至2010年4月在海南自交得到F5和F6(185个家系)。构建群体过程中在幼苗两叶一心期喷洒300 mg·kg-1硝酸银3次,每隔3 d喷1次,注意硝酸银喷洒要在光线较弱的早上或傍晚进行,诱导纯雌株产生完全花以保证自交传代。
1.2 田间试验和性状调查
2010年春秋两季分别种植P1、P2、F1、185个F6:7家系。试验采用随机区组设计,P1、P2、F1每个重复各12株,3次重复;185个F6∶7家系,3次重复,每次重复5株,株距0.4 m,行距0.6 m,自然生长不整枝。调查子蔓第1雌花开花期(DFF),即每个单株从播种到第1雌花完全开放的天数,每个重复内的所有单株取平均值。
1.3 遗传数据分析方法
将DFF数据用盖钧镒等(2003)提供的“RIL群体主基因+多基因混合遗传模型分离分析方法软件”进行分析,其方法是用IECM算法估计各种遗传模型的极大对数似然函数值和AIC值,从这些模型中选择AIC值较低的模型进行适合性检验,适合性检验参数达到显著差异个数最少的模型即为最优模型,然后根据最优模型估计一阶遗传参数和二阶遗传参数,一阶遗传参数由软件自动生成,二阶遗传参数估计方法如下:
在混和遗传模型中,表型值(p)表示为群体平均数(m)、主基因效应(g)、多基因效应(c)和环境效应(e)之和,即p=m+g+c+e,因此,表型方差()可表示为主基因方差()、多基因方差()和环境方差()之和,即群体的主基因遗传方差()等于群体表型方差减去纯合主基因成分分布方差,群体的多基因方差()等于该群体纯合主基因成分分布方差减去误差方差(),其中(n为RIL家系的观测个体数,δ2为P1、P2估算的方差),为RIL群体的表型方差,由群体家系观察值间的变异估计。群体的主基因遗传率;群体的多基因遗传率。
2 结果与分析
2.1 亲本及F1第1雌花开花期比较
春秋两季不同环境条件下,父母本及 F1DFF的表型值如表 1所示,春秋两季 WI998的 DFF均显著早于 3-2-2;F1的 DFF平均值偏向父本,即第1雌花开花期晚对第1雌花开花期早接近显性。

表1 不同季节父母本及F1 DFF比较分析 d
表2 不同季节RIL重组自交系群体DFF分析
2.2 RIL群体第1雌花开花期次数分布特征
从表 2可以看出,甜瓜 DFF性状在 RIL群体中,春秋两季均表现出明显分离,春季和秋季2次调查结果显示在RIL群体中分布的偏度均小于1,峰度小于 3,说明其为数量性状遗传的典型分布。DFF在重组自交系群体中的频次分布亦验证了相同的结论(图1)。
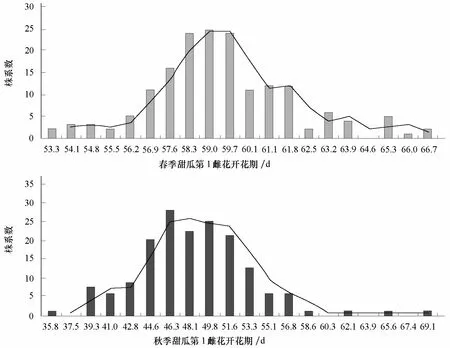
图1 不同季节甜瓜DFF性状在RIL群体中的次数分布
2.3 最优模型的选择、检验
将亲本及RIL群体不同季节DFF数据按照盖均镒等(2003)提出的有关RIL群体主基因+多基因混合遗传模型数量性状分离分析方法来选择最优遗传模型,计算了65种假设遗传模型的最大似然函数值和AIC值,DFF性状存在一些模型所对应的基因效应,结果见表3。
从表3中可知,春季DFF性状对应的模型E_1_3、I_1、I_2、I_4的AIC值空白,表明春季RIL群体内DFF性状不存在这些模型所对应的基因效应。模型E_1_7、E_1_8、E_1_9和G_3的AIC值相对其他模型较低(表3),作为备选模型,依据统计量的均匀性检验,nW2的 Smirnov检验及Dn的Kolomogorov检验对分布进行适合性检验(表4),4个模型均有7个统计量达到显著差异,而前3个模型的AIC值也相同,再根据主基因间作用方式简单化原则,E_1_8模型(2对重叠性主基因+加性多基因模型)可以作为春季DFF遗传的最适模型。
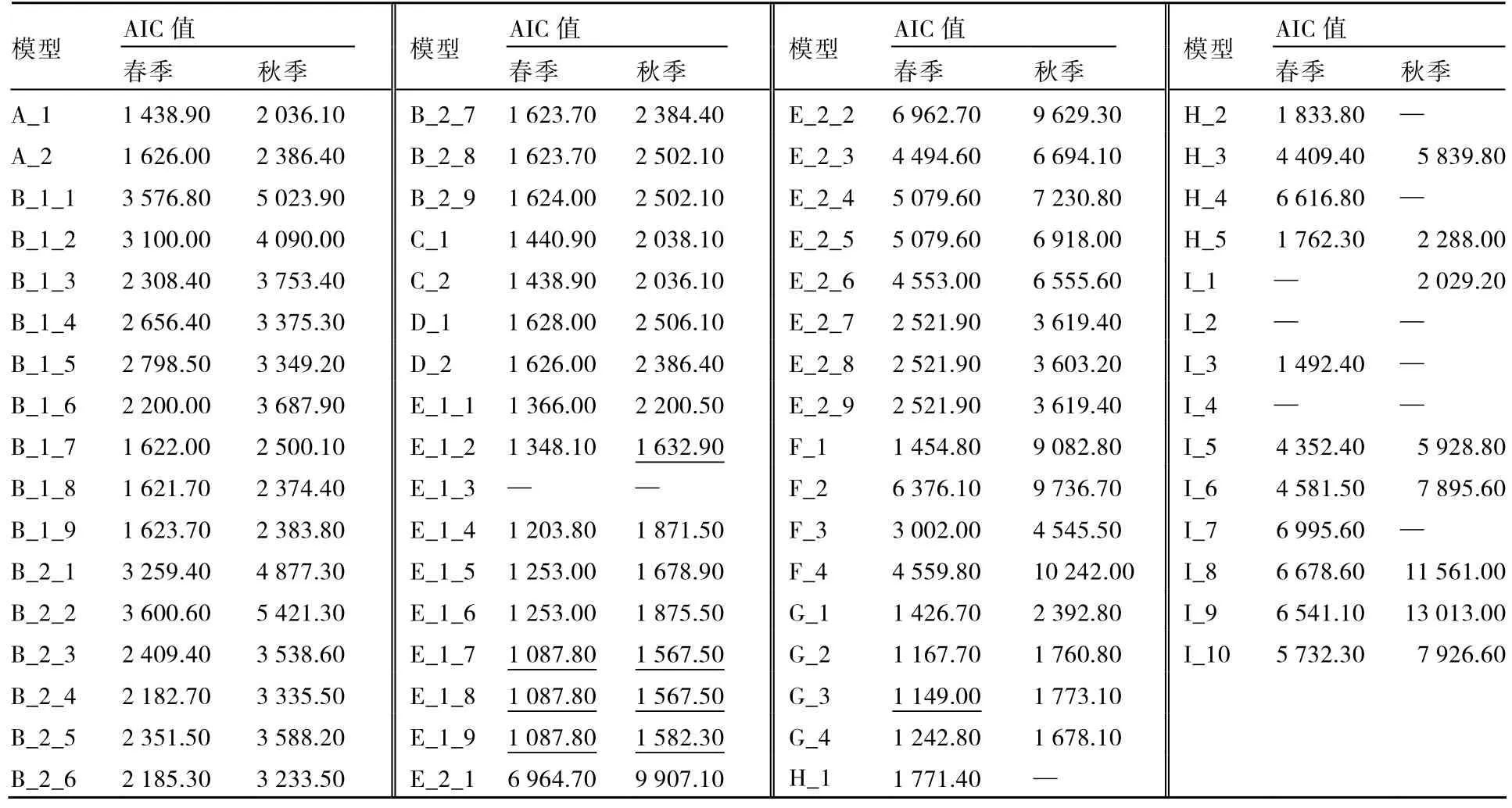
表3 RIL群体中DFF不同遗传模型的AIC值
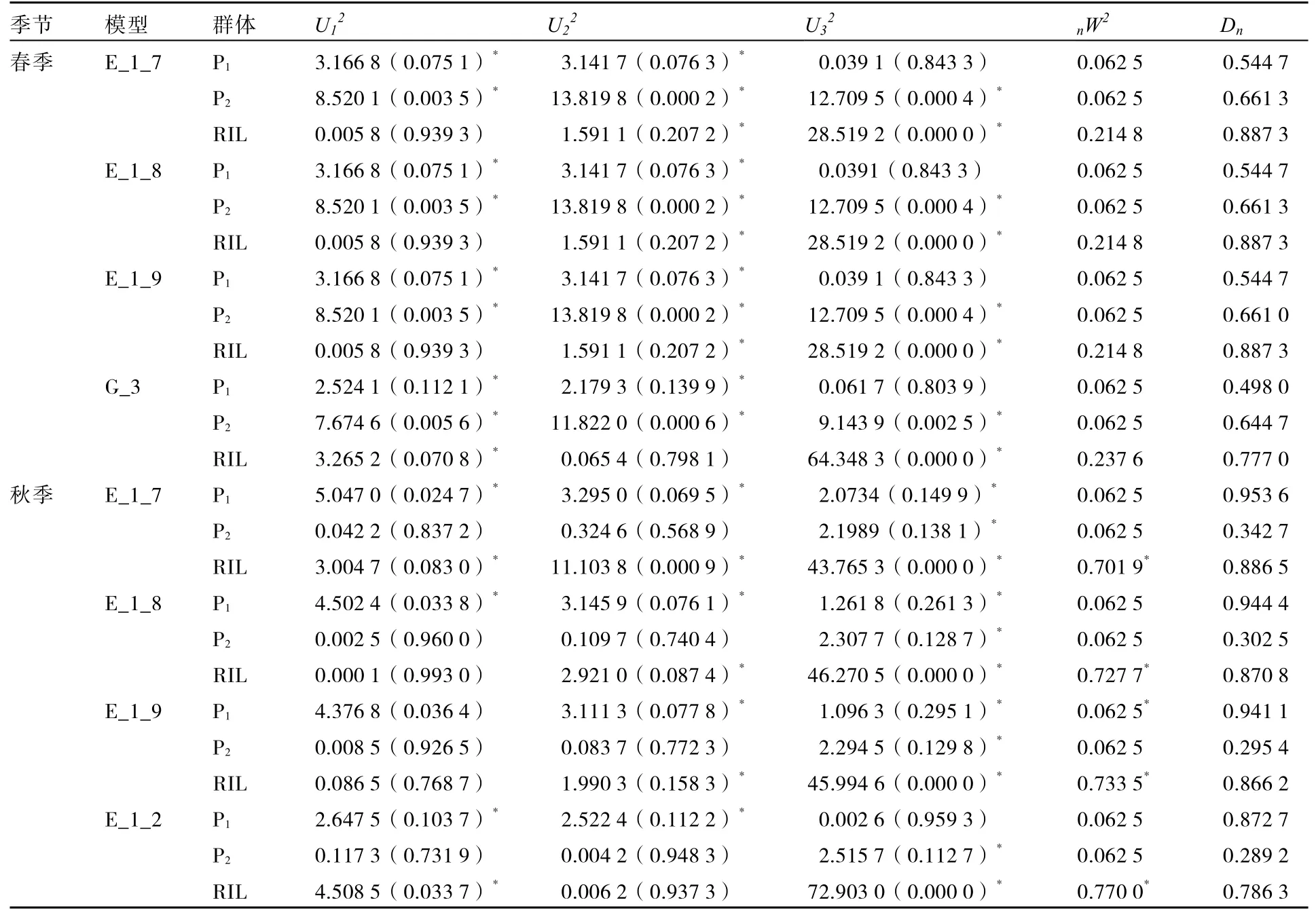
表4 DFF性状备选模型的适合性检验
从表 3中可知,秋季 DFF性状对应的模型 E_1_3、H_1、H_2、H_4、I_2、I_3、I_4、I_7的AIC值空白,表明秋季RIL群体内DFF性状不存在这些模型所对应的基因效应。模型E_1_7、E_1_8、E_1_9、E_1_2的AIC值相对其他模型较低,且4个模型间AIC值与极大似然函数值相比较,差异不大。依据统计量的均匀性检验,nW 2的Smirnov检验及Dn的Kolomogorov检验对分布进行适合性检验(表 4),E_1_8和和 E_1_9均有 7个统计量达到显著水平,E_1_7和E_1_2模型各有8个和6个统计量达到显著水平,E_1_8与E_1_7的AIC值最小,但E-1-8达显著水平的统计量数值较E_1_7少,所以E_1_8模型可以作为秋季DFF遗传的最适模型。
2.4 最优模型的遗传参数估计
利用软件分析结果得到春季DFF模型(E_1_8)、秋季DFF模型(E_1_8)的分布参数估计一阶遗传参数值(表5),并进一步估计二阶参数(表6)。
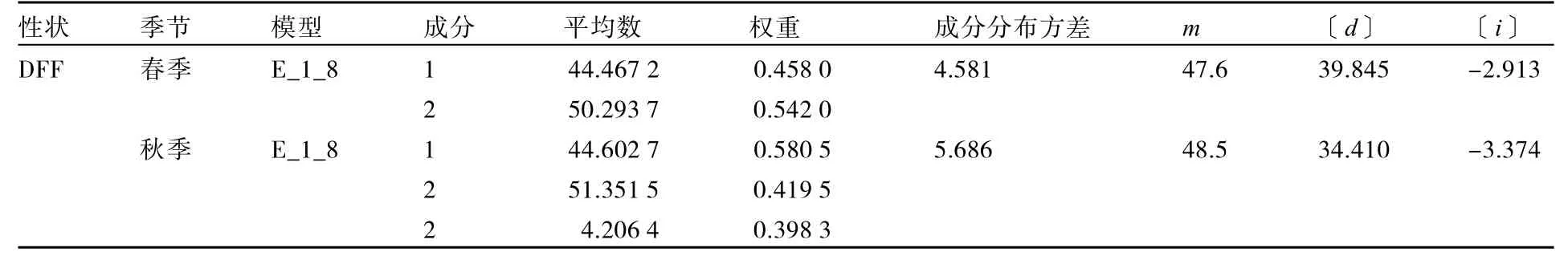
表5 春秋两季DFF的一阶遗传参数估计值
春季控制DFF遗传的2对主效基因表现为重叠性效应,多基因以加性效应为主(〔d〕=39.845)(表5),主基因遗传率较高(77.05%),多基因遗传率较低(22.44%)。秋季控制 DFF遗传的2对主效基因亦表现为重叠性效应,多基因以加性效应为主(〔d〕=34.410),主基因遗传率较高(72.38%),多基因遗传率较低(27.52%)。可以看出春秋两季DFF的遗传比较稳定,均受2对重叠性主基因和加性多基因控制。
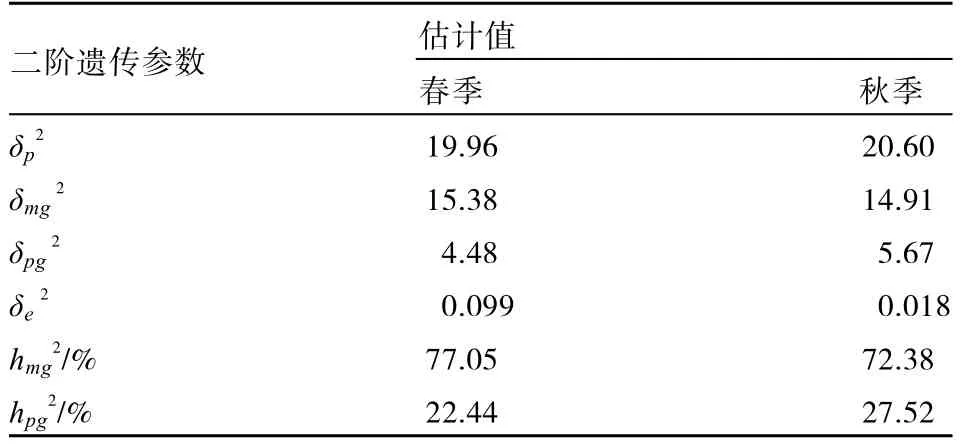
表6 春秋两季DFF二阶遗传参数估计值
3 结论与讨论
近年,研究者利用多世代群体(P1、P2、F1、BC1P1、BC1P2、F2)开展了不同性状遗传特性的研究(陈学军 等,2006;苏小俊 等,2007;张红 等,2009),而此群体为临时群体,无法进行多年多点试验,难以获得稳定的 QTL。郭绍贵等(2005)指出尽可能多的聚合与环境不发生互作的增效主基因或QTL是分子育种的重要目标,因此,进行多年或多点试验确定基因位点的遗传稳定性是十分必要的,重组自交系群体作为永久性群体可以进行多年或多点的重复试验,郭绍贵等(2005)利用重组自交系群体在新疆和北京不同生长环境中对果实可溶性固形物含量进行比较分析,得出了可能是控制可溶性固形物含量性状表达的主要 QTL。另外,有研究者利用RIL群体研究了不同性状的遗传特性(李侠 等,2009;薄凯亮 等,2011;马政 等,2011)。本试验利用重组自交系群体在不同环境(春季和秋季)下,对甜瓜第1雌花开花期遗传做了重复试验,不同季节得出了同一个遗传模型(E_1_8),说明第1雌花开花期在不同环境(季节)遗传稳定,环境对第1雌花开花期的遗传影响不大,可以进行早代选择。
有关甜瓜早熟性状构成因素遗传研究报道较少,尚未见甜瓜第1雌花开花期遗传的报道。本试验利用RIL群体主基因+多基因模型分析法,对甜瓜早熟性状构成因素之一,第1雌花开花期的遗传特点进行了研究,不仅检测了多基因效应,而且鉴别了主基因的存在并估计其遗传效应。
本试验采用主基因+多基因混合遗传模型分析法,对重组自交系群体第1雌花开花期进行了初步研究,结果表明第1雌花开花期由2对具有重叠效应的主基因控制,同时受加性多基因的控制,这一试验结果将有利于目标基因的定位。一般利用分子标记进行农艺性状的基因定位,如果对目标性状的遗传模式不清楚,则需构建覆盖全基因组的标记连锁图。然而,根据本试验结果,可以在采用BAS法筛选到与2个主基因连锁的分子标记的基础上,在目标染色体区段加大分子标记密度建立局部饱和的标记连锁图,实现目标基因的快速定位。匡勇等(2011)利用重组自交系群体研究了水稻穗数的遗传,认为在基因定位之前进行目标性状的遗传模型分析,将使基因定位具有明确的方向性,既可加快基因定位进程,也可减少工作量和降低成本。
薄凯亮,沈佳,钱春桃,宋慧,陈劲枫.2011.“北京截头”×西双版纳黄瓜重组自交系群体重要农艺性状的遗传分析.南京农业大学学报,34(11):20-24.
陈学军,陈劲枫,方荣,程志芳,王述彬.2006.辣椒始花节位遗传研究.园艺学报,33(1):152-154.
盖钧镒,章元明,王建康.2003.植物数量性状遗传体系.北京:科学出版社:120-126.
郭绍贵,许勇,张海英,宫国义,王永健.2005.不同环境效应对西瓜果实可溶性固形物含量的影响.中国蔬菜,(s):13-16.
匡勇,罗丽华,周倩倩,何云丽,范锡麟,肖应辉.2011.水稻籼粳交自交系群体穗部性状的相关和遗传分析.华北农学报,26(3):72-78.
李侠,常玮,韩英鹏,滕卫丽,李文滨.2009.大豆种子脂肪酸含量的遗传分析.大豆科学,28(3):404-407.
马政,薄凯亮,冀刚,钱春桃,陈劲枫.2011.西双版纳黄瓜RIL 群体叶绿素含量的遗传分析.中国瓜菜,24(6):11-13.
苏小俊,徐海,袁希汉,陈劲枫.2007.普通丝瓜第1雌花节位遗传研究.福建农业学报,22(2):154-157.
张红,王怀松,贺超兴,张志斌,张显,伊鸿平,吴明珠.2009.甜瓜糖酸性状的遗传研究.园艺学报,36(7):989-996.
Luan Feishi,Sheng Yunyan,Wang Yuhan,Staub J E.2010.Performance of melon hybrids derived from parents of diverse geographic origins.Euphytica,173:1-16.
Monforte A J,Oliver M,Gonzalo M J,Alvarez J M,Dolcet-Sanjuan R,Arus P.2004.Identification of quantitative trait loci involved in fruit quality traits in melon (Cucumis meloL.).Theor Appl Genet,108(4):750-758.
Tyagi S V S,Khandelwal R C,Maurya I B.2003.Combining ability analysis over environments for fruit yield and other traits in ridgegourd(Luffa acutangulaRoxb.).Scientific-Horticulture,(8):149-157.
猜你喜欢
中国农学通报(2022年29期)2022-11-25
作物研究(2020年5期)2020-12-08
北京农学院学报(2020年1期)2020-03-09
北京农学院学报(2019年1期)2019-02-22
农业知识(2018年18期)2018-05-28
现代农业科技(2018年22期)2018-01-15
绿色科技(2017年7期)2017-05-12
江苏农业科学(2016年12期)2017-04-05
创新作文(3-4年级)(2016年9期)2016-12-22
农民致富之友(2016年5期)2016-10-21
