
神农箭竹更新苗生物量龄级分配模型
2012-11-21 07:36王玲孟银萍赵丽娅李兆华
湖北大学学报(自然科学版) 2012年2期
王玲,孟银萍,赵丽娅,李兆华
(湖北大学资源环境学院,湖北 武汉430062)
竹子是集经济、生态和社会效益于一体的重要克隆植物,在区域经济社会发展和环境保护中发挥着越来越重要的作用,被誉为21世纪最有发展前景的植物类型[1].神农箭竹(Fargesiamurielae(Gamble) Yi)原产于我国神农架地区,地下茎合轴型,笋期5月[2].本种在神农架是垂直分布最高的一种竹子,生于海拔2 800~3 000 m的冷杉林下或灌木丛中,在欧洲庭园中普遍栽培,是欧洲引种我国高山竹类最成功的一种,同时也是珍稀哺乳动物大熊猫的重要主食竹种之一,对山地水土保持、减缓地表径流、涵养水源、调节小气候环境、促进农业稳产丰产等都起着不同程度的有利作用.神农箭竹种源非常稀少,1996-2000年神农架自然保护区内的神农箭竹大面积开花死亡[3-4],2004-2005年,该保护区内金猴岭地区又有大量竹子开花死亡.而目前对神农箭竹的研究主要集中在种群[5]、 群落的海拔多样性[6]、 植被的更新[7]、 种子的萌芽和分布[8-9]等方面.
生物量研究不仅是林业科学研究的一项重要内容[10],也是植物群落最重要的特征之一,其值的大小表明植物固定和积累空间资源的能力[11].生物量作为生态系统研究的本底数据,是评价植物生长状况和生态系统功能的重要指标[12].关于竹类植物的生物量结构和生物量模型,近年来我国的学者做了大量研究,主要集中在很多优良的经济竹种上,地域主要集中分布在浙江、福建等地,如毛竹(Phyllostachysedulis)、麻竹(Dendrocalamuslatiflorus)、苦竹(Arundinariaamarus)、雷竹(Ph.violascens)等[13-17].而关于竹类植物中的高山竹种生物量结构和生物量模型的研究报道比较少,迄今可见的高山竹种仅有缺苞箭竹(Fargesiadenudata)、筇竹(Chimonobambusatumidinoda)、巴山木竹(Arundianariafargesii)等几篇报道[18-20],对神农箭竹生物量结构的研究较为缺乏.本研究旨在通过对神农箭竹种群生物量结构的探索,以期掌握其生长的基本规律,为进一步研究神农箭竹觅养机制及生态保护工作提供理论依据.
1 研究地区与研究方法
1.1研究地概况本项研究实验地位于神农架西南部神农顶旅游区凉风垭(N31°26′55.7″,E110°14′23.2″),属于中纬度北亚热带季风区,受大气环流控制,其地理区位见图1.海拔高2 773 m,坡度20~30度左右,年平均气温在0℃以上,有的地方可达5 ℃,最热月七月的平均气温可达18 ℃,无霜期100~120 d,≥10 ℃的积温小于1 650 ℃,干燥度0.21.年降雨量在顶峰附近可达2 500 mm,云雾多,湿度高,风较大,一年中有相当长的冰冻期,冬季积雪厚度可达1~2 m以上,低温冻融交替和高温,对岩石风化、植被类型与土壤形成有重要影响.该地区植被为巴山冷杉林纯林或箭竹—冷杉林,土壤为草甸暗棕壤,土层较厚,地表有竹根密织形成毡毛层或草甸层,腐殖质层也较厚,属典型的亚热带季风性湿润气候,常年大风.
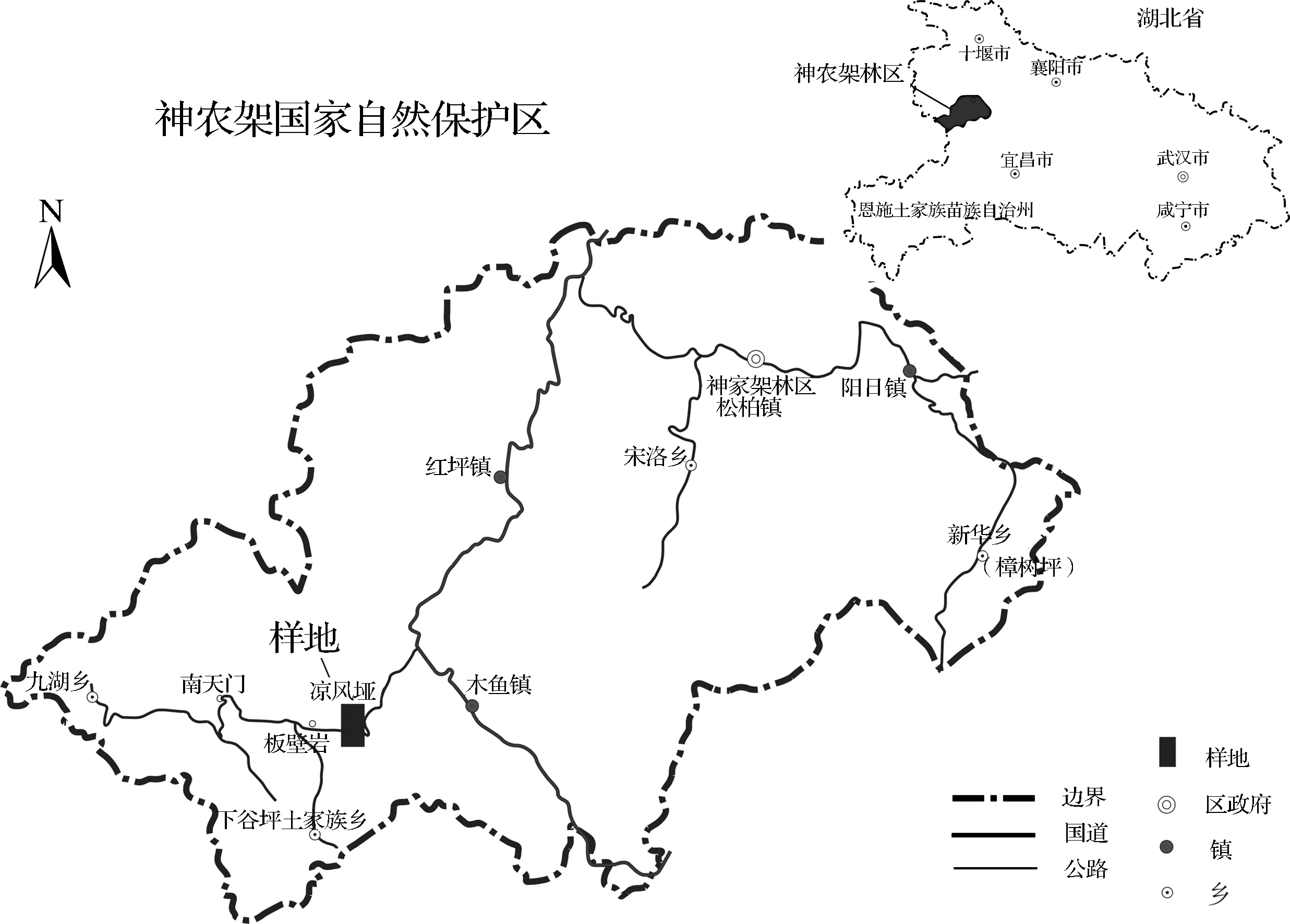
图1 研究样地地理区位图
1.2材料与方法凉风垭神农箭竹为野生状态,但成片分布,且为神农箭竹纯林,上部无其他灌木、乔木,只有少许草被层,人为干扰较少.试验于2011年8月进行,在凉风垭神农箭竹林中随机选取1、2、3、4、5、6 a及6 a以上神农箭竹无性分株各10株,并深入土壤30 cm连根完整挖出,洗净根部土壤,之后分别进行地上部分秆胸径、基径、秆高、地下茎长度、根径等数据的测量及记录.之后按秆、枝叶、地下茎等几部分分装带回实验室,用烘箱于80 ℃恒温下烘干24 h,冷却后在电子天平(精确到0.000 1 g)上称重,计算各器官含水率和生物量.
将数据收集输入计算机,运用SPSS Statistics 17.0对所测数据进行统计分析.
2 结果与分析
2.1神农箭竹地上部分不同年龄分株生物量分配规律竹类是不同年龄立竹构成的异龄林,由于生长年限的不同,各年龄的竹株所进行的光合作用的时间有所差异,因此其生物量及其分配必然存在差异[21],生物量在不同年龄竹的分配是不同的[22-23].将收集到的生物量数据进行龄级分析,得到不同龄级分株生物量的分配规律.由于神农箭竹每节分枝较多,且分枝纤细短小,叶片繁多密集,枝叶混杂不易分离,因此将地上部分分为秆、枝叶两部分进行测量分析.神农箭竹地上部分各器官生物量分配见表1.
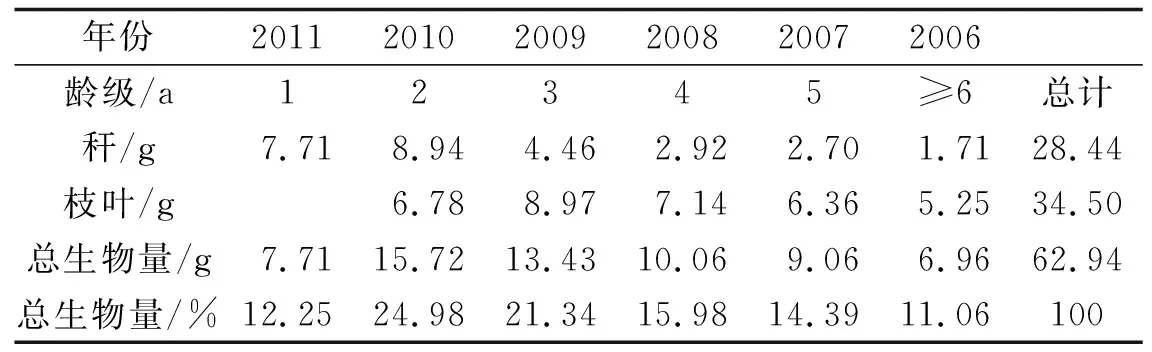
表1 神农箭竹地上部分各器官生物量分配
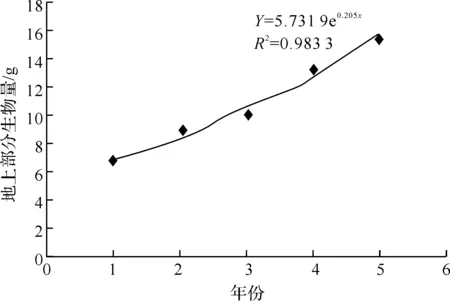
图2 神农箭竹不同龄级地上部分生物量分布图
将神农箭竹地上部分生物量数据绘制散点图,并添加趋势线(如图2所示).可以看出地上部分生物量随时间变化规律符合指数函数回归模型,拟合方程为:
Y=5.731 9 e0.205 4x
其中x以2006年为基点取值为1,2007年取之为2,依此类推.R2=0.983 3,趋近于1,说明此指数回归模型的拟合优度很高.
由表1 看出,神农箭竹地上部分生物量在龄级上的分配为2龄>3龄>4龄>5龄>1龄>6龄及以上.1龄竹的地上部分生物量低于2龄竹,不符合上述规律,是由于本样竹采于2011年8月1日,距神农箭竹的出笋期5月仅3个月左右,1龄竹还未完成高生长,因此所测1龄竹为竹笋,还未抽枝展叶,生物量较低,为减小误差,分析时以2龄竹为基点进行分析.地上部分生物量在各龄级中的分配以2龄竹为最多,占24.98%;3龄以下竹占58.57%;4、5、≥6龄竹所占比例依次减少,分别为15.98%、14.39%、11.06%.竹秆生物量基本符合随龄级增大而逐渐下降的趋势,与总生物量的变化规律一致,说明竹秆生物量在各器官生物量分布中占主导地位,且竹秆生物量的变化较稳定.根据植物生长规律,植物的生物量增长应符合一般生长曲线,从图1可以看出神农箭竹地上部分生物量随时间呈现指数增长的趋势,这是因为本研究采集神农箭竹样本处于其克隆生长周期的前期阶段,即“S”型曲线的“J”形增长阶段.
2.2秆高、胸径、基径、地下茎直径、地下茎长与各器官生物量的相关性分析遗传的相对稳定性使得同种竹类植物个体的不同器官间必然存在结构上的联系,表现在外形上的同种相似性和个体稳定性,并随着环境因子的改变相应发生不同程度的变异.而竹类个体不同器官之间的联系反映着竹子个体的生长规律[24].通过以往对竹林生物量模型的研究,可以看出竹类植物是一个有机的生命体,各组织相互联系,各个器官的生物量也存在着密切的关系[25].神农箭竹各器官生物量及度量指标的相关分析结果见表2.

表2 神农箭竹各器官生物量及部分特征因子的相关性
*,在 0.05 水平(双侧)上显著相关;**,在 0.01 水平(双侧)上显著相关.
由表2可以看出神农箭竹地下茎直径与秆重、胸径、地下茎长、基径的相关性在0.01水平上呈现显著相关,相关系数分别为0.949、0.841、0.890、0.927,胸径与秆重、基径、地下茎直径呈现极显著相关,相关系数分别为0.903、0.897、0.841,同时与秆高、干重、枝叶重、地下茎长也呈现显著相关关系.这与在野外的实际调查是一致的,即地径和胸径越粗,秆高及秆和枝叶质量就越大.通常竹类植物的胸径为比较容易获得的数据,且胸径与各构建因子均呈现不同程度的相关关系,因此,可以用神农箭竹立竹胸径对各器官和地上部分总生物量进行估算.
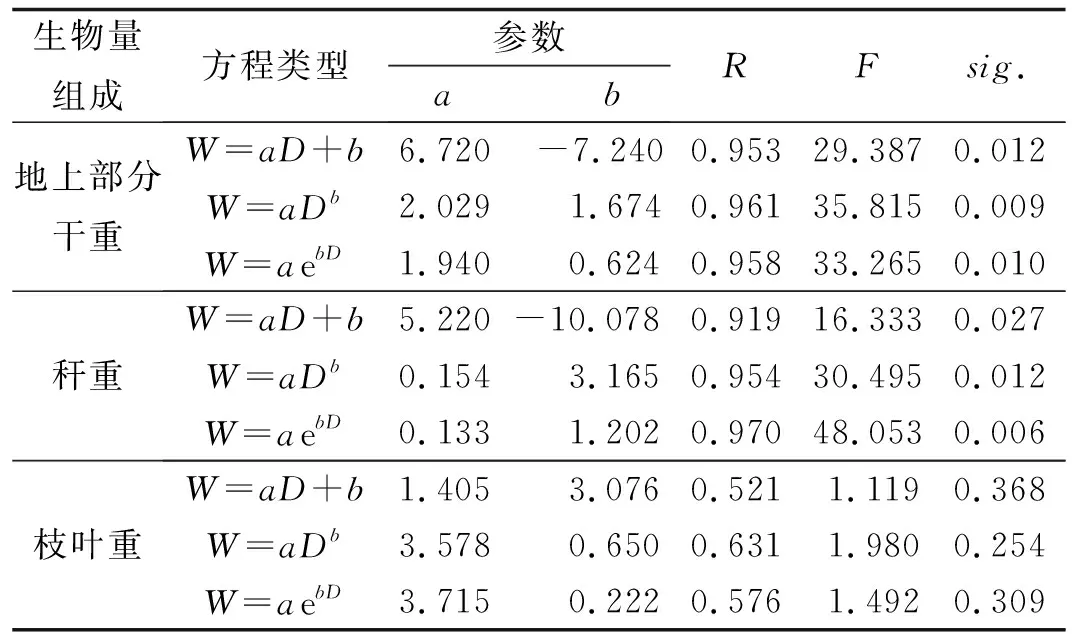
表3 各器官生物量与胸径的回归方程
2.3各器官生物量与胸径相关性及回归模型为了便于估算神农箭竹生物量的大小, 对神农箭竹秆质量、枝叶重量和地上部分干重与胸径(DBH)的相关模型进行拟合, 并选择一元线性函数、幂函数和指数函数模型分别进行曲线拟合,建立以胸径估测其他各器官质量的数学模型,胸径拟合结果见表3.
从表3可以看出,地上部分干重、秆重与胸径的3种拟合模型的sig.值均小于0.05,说明3种方法的拟合模型都是适用的,且具有显著的统计学意义.3种方法的相关系数(R)均在0.90以上,表明地上部分干重、秆重与胸径之间的密切程度很高.相反,枝叶重与胸径的拟合模型效果较差,说明枝叶重与胸径的相关关系并不显著,与表2结果相同.地上部分干重与胸径的3种拟合模型中,根据各指标对比显示幂指数模型最适用,方程式为W=2.029×D1.674.秆重与胸径的指数模型最适用,方程式为W=0.133×e1.202D.
3 结论与讨论
神农箭竹原产于湖北神农架地区,是典型的高山小径竹类,对高山防风护坡及水土保持具有重要的生态价值.据研究报道,神农箭竹母代已开花死亡,子代开始定居生长,进入到新一轮的克隆生长阶段,这一过程将持续110年之久[26].此次研究样地的神农箭竹均为1996-2000年开花死亡[3-4]后有性繁殖而成,整个群落正处于其生长周期的早期定居阶段.通过对神农箭竹更新苗生物量龄级分配模型的研究,得到以下结论:(1)神农箭竹地上部分生物量随时间变化规律符合指数函数回归模型,拟合方程为:Y=5.731 9 e0.205 4x,且R2=0.983 3,竹秆生物量与总生物量的变化规律一致,基本符合随龄级增大而逐渐下降的趋势;(2)地下茎直径、胸径与各器官生物量都存在不同程度的相关关系,地上部分干重与胸径的最优拟合模型为幂函数模型,方程式为W=2.029×D1.674,秆重与胸径的指数模型最适用,方程式为W=0.133×e1.202D.
生物量值的大小表明植物固定和积累空间资源的能力[11].老龄竹在其生长旺盛期,整个种群环境条件不稳定,因此需要不断调整自身生长机能以适应外部环境,而幼龄竹的生长环境相对而言已趋于稳定,受内外干扰影响较少.此外老龄竹自身活力降低,缺枝少叶,生物量积累弱.总体而言,凉风垭样地的神农箭竹种群结构比较年轻化,幼龄竹生理整合能力高于老龄竹,能更好利用生境资源促进自身的营养生长,而老龄竹生理更新能力老化,种群整体处于快速增长的阶段,因此地上部分生物量随时间呈现指数增长的趋势.生物量在各器官的分配比例因树种的不同而异, 反映植物对环境资源的不同利用方式[27].2、3、4、5、6 a及6 a以上神农箭竹枝叶生物量占地上部分总生物量的比例分别为43.13%、66.79%、70.97%、70.20%、75.43%,较材用竹大[13-17],说明神农箭竹枝叶生长较为茂盛,这对竹种的生长较为有利, 但用材效率则会降低.在神农箭竹生长周期的早期定居阶段,母竹通过自身的觅养机制促进克隆体朝向延续种群稳定的方向生长,随着龄级的年轻化,神农箭竹种群生长的稳定性增强,枝叶生物量比例逐渐降低.
由于本次生物量研究的调查范围与采样力度的限制,没有足够的实测值与理论值比较,因此无法检验生物量拟合模型的可靠性,评估模型应用效果的准确性.如要在生产实践中应用,还需进一步的调查研究.此外,本研究的采样是随机抽样,其结果未能全面反映神农箭竹无性系种群生物量结构动态.神农箭竹无性系种群生物量结构是否在不同密度、不同立地条件(海拔、坡向、坡位、土壤、枯落物)等诸多因素下有差异,还有待研究.
[1] 郭子武,李迎春,杨清平,等.花吊丝竹立竹构件与生物量关系的研究[J].热带亚热带植物学报,2009,17(6):543-548.
[2] 易同培,史军义,马丽莎,等.中国竹类图志[M].北京:科学出版社,2008.
[3] 李兆华,赵丽娅,卢进登.“易根”:竹子群体开花的生态诠释[J].世界竹藤通讯,2004,2(4):21-23.
[4] Li Z H, Denich M, Borsch T. Effects of bambooFargesiamurielaeon plant diversity in fir forest on Mountain Shennongjia[J].Forest S tud China, 2004, 6 (4):17-22.
[5] Li Z H, Deng L, Zhao B Y,et al. Species and distribution of mountain bamboos in Shennongjia, Central China[J].J For Res, 2003, 14 (1):35-38.
[6] Li Z H, Denich M. Elevational diversity of arrow bamboo (Fargesiaspathacea) communities on Mount Shennongjia in Central China[J].J For Res, 2002, 13 (3):171-176.
[7] Li Z H, Denich M. Postfire regeneration of umbrella bamboo(Fargesiamurielae) on Mount Shennongjia in Central China[J].Ecol Env & Cons, 2002, 8:201-205.
[8] Li Z H, Deng L, Zhao B Y,et al. Shoot sp routing of arrow bamboo Fargesia spathacea along an altitudinal gradient in Central China[J].B am J,2003,20:1-11.
[9] Li Z H, Denich M, Deng L. Seed production and dispersal of umbrella bambooFargesiamurielaeaftermass flowering in central china[J].B am J,2004, 21:8-17.
[10] 郑景明,张育红.林分生物量研究综述[J].辽宁林业科技,1998(4):43-45.
[11] 徐雪娇,刘济明,徐国瑞,等.不同小生境中小蓬竹的含水率及生物量分配规律[J].贵州农业科学, 2010, 38(10):163-166.
[12] 杨春花,周小平,王小明.卧龙自然保护区华西箭竹地上生物量回归模型[J].林业科学, 2008, 44(3):114-123.
[13] 黎曦,鲍雪芳,王福升.赣南毛竹生物量研究[J].安徽林业科技,2007,131:9-11.
[14] 梁鸿桑,陈学魁.麻竹单株生物量模型研究[J].福建林学院学报,1997,18(3):260-262.
[15] 郑容妹,郑郁善,丁闽锋,等.苦竹生物量模型的研究[J].福建林学院学报,2003,23(1):61-64.
[16] 蒋俊明,费世民,李吉跃,等.苦竹各器官主要营养元素分布及采伐的养分输出[J].四川林业科技,2007,28(2):15-19.
[17] 金爱武,周国模,马跃,等.雷竹各器官生物量模型研究[J].浙江林业科技,1999,1(2):7-9.
[18] 邵际兴.箭竹地上部分鲜生物量的测定[J].甘肃林业科技,1986,3:33-36.
[19] 董文渊,黄宝龙,谢泽轩,等.筇竹无性系种群生物量结构与动态研究[J].林业科学研究,2002,15(4):416-420.
[20] 王太鑫,丁雨龙,李继清,等.巴山木竹种群生物量结构研究[J].竹子研究汇刊,2005,24(1):19-24.
[21] 林益明,林鹏,叶勇.绿竹种群生物量结构研究[J].竹子研究汇刊,1998,l7(2):9-13.
[22] 郑郁善,洪伟.毛竹丰产年龄结构模型与应用的研究[J].林业科学,1998,34(3):325-333.
[23] 薛萍,汪晓萍,李锡泉.湖南衡阳紫色页岩地区芦竹生物量的动态规律[J].中南林学院学报,2006,26(3):11-14.
[24] 熊壮,董文渊,刘时才,等.梁山慈竹秆形结构和地上生物量结构研究[J].林业科技开发,2010,24(3):81-83.
[25] 张鹏,黄玲玲,张旭东,等. 滩地硬头黄竹生物量结构及回归模型的研究[J].竹子研究汇刊,2009,28(3):25-28.
[26] Li Z, Denich M, Borsch T. Simultaneous flowering of umbrella bamboo (Fargesiamurieliae, Poaceae) at its native home in Central China[J].J For Res, 2006, 17(4):293-297.
[27] Tilman D. Plant strategies and the structure and dynamics of plant communities[J].Princeton:Princeton University Press,1988:52-97.
猜你喜欢
湖南生态科学学报(2022年4期)2022-11-11
河南农业大学学报(2020年2期)2020-05-22
山西林业科技(2019年1期)2019-05-27
小天使·二年级语数英综合(2019年2期)2019-01-12
幼儿画刊(2018年5期)2018-06-05
东北林业大学学报(2014年8期)2014-08-02
河南科技(2014年5期)2014-02-27
茶叶通讯(2014年4期)2014-02-27
江西农业大学学报(2013年3期)2013-11-05
