
不同砧木对核桃光合特性的影响
2013-04-04 05:43孟丙南张俊佩徐虎智王少明郭志民
经济林研究 2013年2期
孟丙南 ,张俊佩 ,裴 东 ,徐虎智 ,王少明 ,郭志民
(1.林木遗传育种国家重点实验室,北京 100091;2.中国林业科学研究院 林业研究所,北京 100091;3.河南省洛宁县林业局,河南 洛宁 471000;4.河南省国有洛宁县吕村林场,河南 洛宁 471000)
核桃是我国重要的木本粮油树种之一[1]。长期以来,我国核桃嫁接以实生核桃为主要砧木,使得培育的核桃苗木整齐度低、坚果质量一致性差[2],特别是早衰已成为制约早实核桃品种持续丰产的关键问题。而美国等核桃生产先进国家在核桃上应用了抗逆性强和生长势旺的黑核桃杂交种作砧木,增强了核桃树体的生长势和抗逆性,提高了其产品的品质,加以现代化栽培管理模式,实现了核桃持续丰产[3-4]。近年来,我国先后引进了美国黑核桃多个优良品种或类型,在此基础上,通过杂交、优选培育了一些生长势旺、抗逆性强的优良品系,这些品系在嫁接时表现出很好的亲和性。
光合作用是植物生长发育的基础,是产量和品质形成的决定性因素,光合产物构成了果树树体90%~95%的干物质,光合效率的高低可以反映出植物生长发育水平的高低[5]。果树砧木对接穗光合能力有较强的影响,这已在柑橘、苹果、梨和葡萄等果树上得到证实[6-11]。
本研究以大田栽培的4年生核桃杂交种中宁异、中宁强、中宁奇、黑核桃和实生核桃砧木嫁接核桃优系“上14”作试材,对嫁接苗的光合特性进行了研究和评定,以期为核桃砧木的合理选择与利用提供理论依据。
1 材料与方法
1.1 试验地与试验材料
试验在河南省洛宁县东宋乡核桃栽培示范园进行。
选用中宁异、中宁强、中宁奇、黑核桃和实生核桃为砧木材料。2009年春,选取粗细、生长势相对一致的苗木材料定植于大田,定植时中宁异、中宁强、中宁奇、黑核桃为1年生扦插苗,实生核桃为1年生实生苗。采用完全随机区组设计,每小区4株,3次重复,株行距为5 m×5 m。于2010年5月12日统一采用大方块芽接法进行嫁接,接穗为核桃优系“上14”。嫁接后对幼树进行常规的田间管理。
1.2 光合日变化曲线的测定
在2012年7月的晴天里,采用Li-6400光合测定仪(美国 LI-COR 公司生产)进行光合指标的测定。从当日6:00~18:00对5种砧木处理的净光合速率(Pn,μmol·m-2s-1)进行测定,每隔2 h测定1 次,即可得出其日变化曲线。每种砧木处理每重复随机选取1 株,共选择3 株生长良好的单株,每株选择树冠中部的枝条中部复叶的3片顶叶进行测定[12]。同时用仪器记录蒸腾速率(Tr,μmol·m-2s-1)、气孔导度(Gs,mol·m-2s-1)、细胞间 CO2浓度(Ci,μmol·mol-1)、环境 CO2浓度(Ca,μmol·mol-1)、叶片温度(Tair,℃)、光照强度(Par,lx)和环境相对湿度(RH,%)。
1.3 光响应曲线的测定
在晴天的上午8:30~11:30之间,对中宁异、中宁强、中宁奇、黑核桃和实生核桃5种砧木处理的光响应曲线进行测定,每种砧木选择3个单株,每株选择树冠南向中部枝条上的中部复叶的3片顶叶进行测定。测定时CO2浓度设定为恒值400 μmol·mol-1,人工光源控制光合有效辐射每5 min改变1 次,光合有效辐射设定值依次为0、20、50、100、200、400、600、800、1 000、1 200、1 400、1 600、1 800、2 000 μmol·m-2s-1。温度和相对湿度采用自然状态测定值。利用二次曲线方程对测定的Pn-Par曲线进行模拟,方程模型为:Y=b0+b1X+b2X2,计算光响应曲线方程的光饱和点(LSP),光补偿点(LCP)和最大Pn。
1.4 光合指标与环境因子间的相关性分析
对各光合指标的日变化值与环境因子进行相关性分析,同时利用逐步回归分析方法,计算Pn、Tr、Ci、Gs的回归方程,探讨环境因子对光合指标的贡献率。
当记者追问起提前储备的原因时,李中南表示,一是考虑到原料价格、环保以及人民币汇率波动等因素;二是今年的复合肥价格从7月底就开始一路上涨,参考去年,几乎也是同期开始上涨,并一直延续至今年的4月份,而且可以看出今年的涨价比去年更加扎实;三是依靠多年的从业经验分析得出,复合肥厂家比较偏向终端市场,一般不会让经销商吃亏。
1.5 数据分析
采用SPSS17.0软件数据处理系统对测定数据进行统计与分析。
2 结果与分析
2.1 光合指标的日变化曲线
测得的不同砧木处理核桃各光合指标值的日变化曲线如图1所示。图1表明,中宁异、中宁强、中宁奇、黑核桃和实生核桃5种砧木处理的净光合速率Pn日变化曲线均呈现单峰曲线,全天Pn的高峰值均出现在8:00,之后Pn值总体呈下降趋势。在光合高峰期,5种砧木处理的Pn从大到小依次是中宁奇(23.53μmol·m-2s-1)>中宁强(22.98μmol·m-2s-1)> 实生核桃 (20.85μmol·m-2s-1)> 黑核桃 (20.50μmol·m-2s-1)> 中宁异 (20.21μmol·m-2s-1),其中,中宁奇、中宁强的Pn值显著大于实生核桃,黑核桃、中宁异和实生核桃的Pn值之间不存在显著差异。胞间CO2浓度Ci表现为先下降后上升的日变化规律,8:00到16:00之间其变化不明显。气孔导度Gs和蒸腾速率Tr日变化曲线也呈现为单峰曲线,随着温度的升高和光照强度的增加,气孔导度的高峰值出现在10:00,蒸腾速率的高峰值出现在12:00。
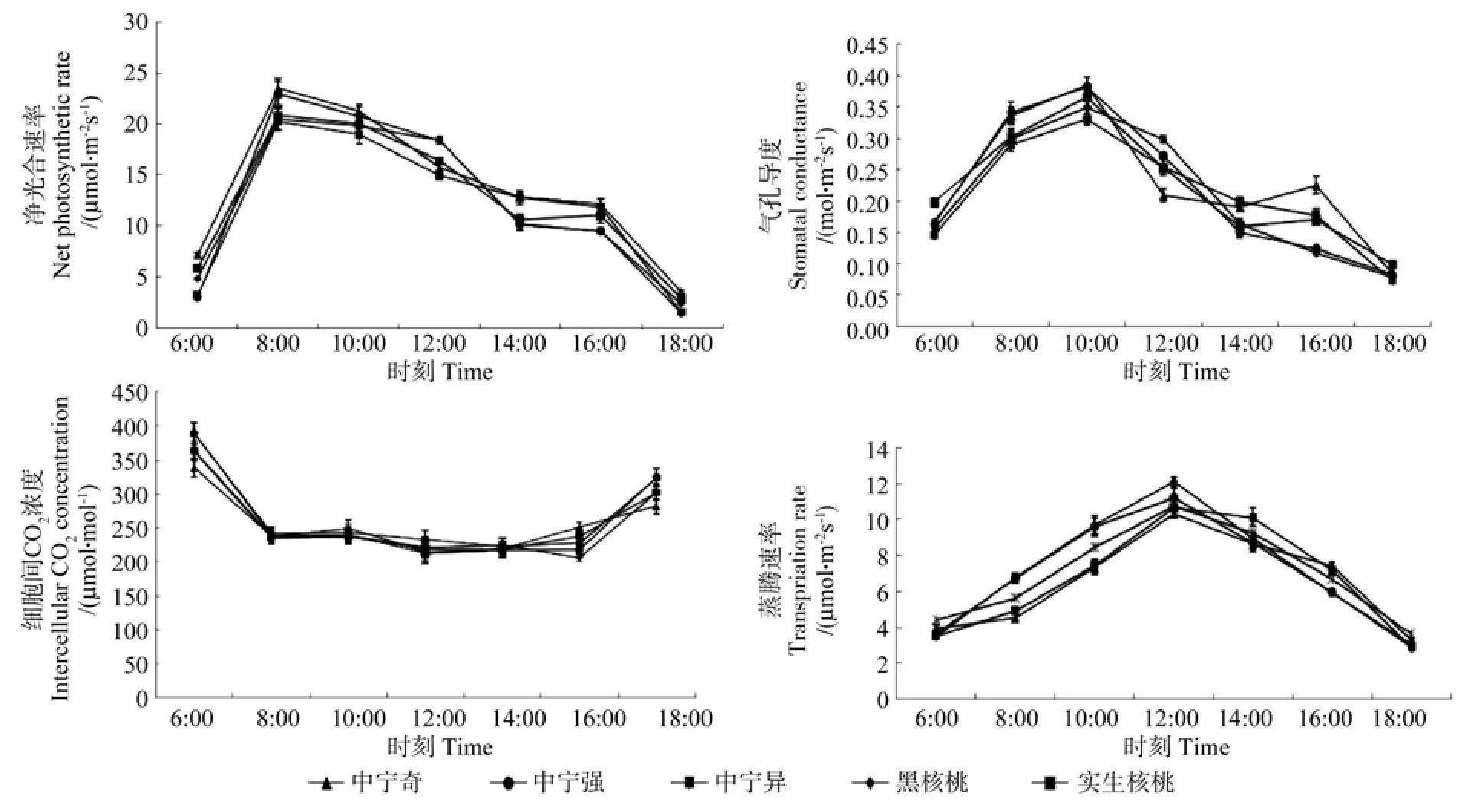
图1 不同砧木处理核桃各光合指标值的日变化Fig.1 Diurnal changes of photosynthetic indexes of walnut in different rootstock treatments
环境因子的日变化趋势如图2所示。图2 表明,环境因子中光照强度Par与叶片温度Tair的日变化曲线呈现为单峰曲线,随着光照强度的增加和温度的上升,Par的最大值(1 896.5 lx)出现在12:00,Tair的最大值(41.39 ℃)出现在 14:00。Ca与RH均表现为先下降后上升的变化趋势。

图2 不同砧木处理核桃光合有效辐射(Par)、空气温度(Tair)、空气CO2浓度(Ca)和空气相对湿度(RH)的日变化Fig.2 Diurnal changes of Par of walnut in different rootstock treatments, Tair, Ca and RH
2.2 光响应曲线
不同砧木处理核桃的光强-光合指标曲线如图3所示。由图3 可知,5种砧木处理的Pn-Par曲线均呈S型,当Par为0时,Pn为负值,随着Par增加,Pn值快速上升,当Par到达800 μmol·m-2s-1时,Pn值上升趋缓;5种砧木处理的Pn-Gs曲线和Pn-Tr曲线均呈缓慢上升趋势,随着Par的增加,Gs和Tr的值不断增加,当Par到达1 800 μmol·m-2s-1时,Gs和Tr的值开始下降;中宁奇和实生核桃砧木处理的Gs和Tr值一直较大,黑核桃和中宁异砧木处理的Gs和Tr值一直处于较低水平。5种砧木处理的Pn-Ci曲线则呈下降的变化趋势,Par到达800 μmol·m-2s-1以后,Ci处于平稳的变化趋势,5种砧木处理间Ci值的差异不显著。
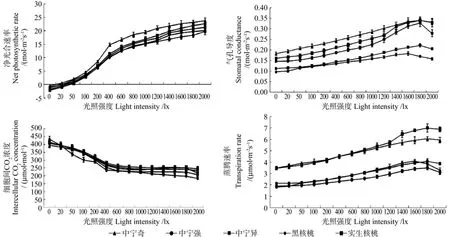
图3 不同砧木处理核桃的光强-光合指标曲线Fig.3 Curves of light intensity- photosynthetic indexes of walnut in different rootstock treatments
5种砧木处理的Pn-Par曲线模拟方程见表1。由表1可知,5种砧木处理的光饱和点(LSP)为1 706.23 ~ 1 648.55 μmol·m-2s-1,黑核桃砧木处理的LSP值最高(1 706.23 μmol·m-2s-1),中宁异砧木处理的的LSP值最低(1 648.55 μmol·m-2s-1)。在光饱和点处的最大光合速率,中宁奇砧木处理(23.12 μmol·m-2s-1)和中宁异砧木处理(22.38 μmol·m-2s-1)均显著高于对照的实生核桃(21.23 μmol·m-2s-1),中宁强砧木处理与实生核桃不存在显著差异。5种砧木处理的光补偿点(LCP)之间差异显著,实生核桃砧木处理的LCP值最高(36.41 μmol·m-2s-1),中宁奇砧木处理的LCP值最低,这表明中宁奇砧木处理在光照很弱的条件下就能进行光合作用。

表1 Pn-Par曲线模拟方程及最大净光合速率(Pnmax)、光饱和点(LSP)和光补偿点(LCP)Table 1 Pn-Par curves simulation equation, maximum photosynthetic rate (Pnmax), Light saturation point (LSP) and Light compensation point (LCP)
2.3 光合指标与环境因子的相关性分析
5种核桃砧木处理的各光合指标之间、各光合指标与各环境因子之间的相关系数见表2。由表2可知,Pn与Gs、Tr之间呈极显著正相关,而与Ci之间呈极显著负相关;环境因子Par与Pn、Gs、Tr之间呈极显著正相关,Par与Ci之间呈极显著负相关;Tair与Tr之间呈极显著正相关,Tair与Ci之间呈极显著负相关;Ca与Ci之间呈极显著正相关,Ca与Tr之间呈极显著负相关;RH与Ci之间呈极显著正相关,RH与Tr之间呈极显著负相关;Ci和Tr与所有环境因子的相关均达到极显著水平。
叶片的环境因子光强、温度、湿度、CO2浓度在时刻地发生变化,且各因子间又相互作用,共同影响各光合指标的变化。为确定各环境因子对光合特性的影响程度,进一步利用逐步多元回归分析方法。各光合指标与环境因子的逐步回归分析结果如表3所示。4个回归模型中,判定系数(R2)都达到了0.8以上。经F检验,因变量和自变量的相关性达到了极显著水平(P<0.01)。4个回归方程中,Pn的主要影响因子是Par和Tair,Gs的主要影响因子是Par和Tair,Ci的主要影响因子是Par和Ca,Tr的主要影响因子是Par和Tair。

表2 光合指标与环境因子的相关系数†Table 2 Correlation coefficients between photosynthesis indexes and environment factors

表3 光合指标的回归分析Table 3 Regression analysis of photosynthesis indexes
3 结论与讨论
自然条件下植物的光合作用日变化曲线多表现为单峰型或双峰型[13]。核桃光合日变化曲线一般呈明显的双峰型[12,14-17],本试验中5种砧穗组合下核桃光合的日变化曲线均呈单峰型,这与前人的研究结果存在差异,这可能是由于测定时环境条件不同所致。一方面,本试验光合测定当日10:00~18:00之间的叶片温度一直维持在35 ℃之上,高于光合作用的适宜温度范围。叶片温度高影响了酶的活性,进而影响了植物本身的生化反应和光合速率;另一方面,光照强度也影响光合日变化的进程,本试验在10:00~16:00之间进行,光照强度普遍大于5种砧木处理的光饱和点,从而抑制了核桃的光合作用,因此,试验中并未出现第2个光合高峰值。
光合作用的过程是极复杂的过程,既受植物自身结构的调节,也受外界环境因子的影响[18]。5种砧木处理核桃光合指标的日变化与各环境因子的相关分析和回归分析结果表明,相关系数极显著或显著的某些环境因子与回归方程中出现的环境因子不太一致,这可能与不同砧木对相同环境条件的敏感程度不同有关,也可能是简单的相关分析本身的片面性造成的,亦可能是没有考虑各因子间相互影响的间接作用所致,这些都有待进一步的研究。
试验结果表明,5种砧木处理的核桃叶片的光合速率、气孔导度和蒸腾速率等光合指标存在显著差异。前人的研究结果表明,砧木对接穗影响的差异表现在叶片上,可造成接穗叶片的叶绿素、自由水和束缚水含量、叶片矿质元素含量和营养物质含量的明显差异,进而影响叶片的光合作用。在相同的外界环境条件下,中宁奇处理的气孔导度值较高,可能是由于其根系发达,能为叶片提供充分的水分,从而表现出较高的净光合速率。光饱和点是反映最大光能利用率的重要光合指标。在光照饱和的情况下,中宁奇和中宁异表现了高的光合速率,表明这两种砧木更适合在光照强度高的地区栽培。砧木对接穗品种的光合特性的影响是一个受内外因子综合影响的非常复杂的生理过程,有关砧木影响果树光合作用的机理尚需进一步的研究。
通过分析5种砧木处理后核桃叶片的光合特性可知,中宁奇、中宁强和中宁异是在一定程度可以提高核桃接穗光合特性的砧木资源,但仍需进一步考察其生长量、树形、抗性等因子,以选择出具有生产应用价值的优良砧木资源。
参考文献:
[1] 谭晓风,马履一,李芳东,等.我国木本粮油产业发展战略研究[J].经济林研究,2012,30(1):1-5.
[2] 冯连芬,吕芳德,张亚萍,等.我国核桃育种及其栽培技术研究进展[J].经济林研究,2006,24(2):69-73.
[3] 陆 斌,宁德鲁,等.美国核桃产业发展综述及其借鉴[J].林业调查规划,2011,36(3):98-105.
[4] 裴 东,吴燕民,奚声柯.美国黑核桃的栽培及在我国的发展前景[J].河北林果研究,2000,15(1):95-100.
[5] 李天忠,张志宏.现代果树生物学[M].北京:科学出版社,2008.
[6] 张建光,刘玉芳,施瑞德.不同砧木上苹果品种光合特性比较研究[J].河北农业大学学报,2004,27(5):31-33.
[7] 胡利明,夏仁学,周开兵,等.不同砧木对温州蜜柑光合特性的影响[J].园艺学报,2006,33(5):937-941.
[8] 姜卫兵,俞开锦,高光林,等.梨不同砧穗组合光合特性[J].园艺学报,2002,29(6):569-570.
[9] 李小红,周 凯,陶建敏.不同葡萄砧木对矢富罗莎葡萄嫁接苗光合作用的影响[J].果树学报,2009,26(1):90-93.
[10] Jover S, Martinez-Alcantara B, Rodriguez-Gamir J. Influence of Rootstocks on Photosynthesis in Navel Orange Leaves: Effects on Growth, Yield, and Carbohydrate Distribution[J]. Crop Science, 2012,52 (2):836 - 848.
[11] Tsipouridis C, Thomidis T. Effect of 14 peach rootstocks on the yield, fruit quality, mortality, girth expansion and resistance to frost damages of May Crest peach variety and their susceptibility on Phytophthora citrophthora [J]. Scientia Horticulturae, 2005,103: 421 - 428.
[12] 王红霞,张志华,王文江,等.田间条件下核桃光合特性的研究[J].华北农学报,2007,(2):125-128.
[13] 董 然,王 莹,赵国禹,等.长白山5种橐吾光合生理日变化及光响应特征研究[J].中南林业科技大学学报,2011,31(9):49-54.
[14] 贺 奇,王 贵,常月梅,等.早实核桃光合特性的初步研究[J].山西农业大学学报:自然科学版),2010,(3).197-200
[15] 刘 鹏,刘庆忠,赵红军,等.核桃光合作用特性的初步研究[J].落叶果树,2003,4:1-3.
[16] 张俊佩,王红霞,高 仪,等.核桃(Juglans regia)光合影响因子的研究[J].河北农业大学学报,2008,(3):33-36.
[17] Le Roux X, Dreyer E. Parameterization and testing of a biochemically based photosynthesis model for walnut (Juglans regia) trees and seedlings [J]. Tree Physiology, 1999, 19(8):481-492.
[18] Kim S H, Gitz D C, Richhard C,et al. Temperature dependence of growth, development, and photosynthesis in maize under elevated CO2[J]. Enviromental and Experimental Botany, 2007,61: 224 - 236.
猜你喜欢
今日农业(2021年19期)2021-11-27
落叶果树(2021年6期)2021-02-12
河北果树(2020年2期)2020-05-25
基层中医药(2018年2期)2018-05-31
现代园艺(2017年21期)2018-01-03
新农业(2017年3期)2017-05-17
陕西画报(2016年1期)2016-12-01
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
少儿科学周刊·儿童版(2015年10期)2015-11-07
