
室内养殖松鼠秋季换毛序及新生被毛部分性状
2013-08-29 09:13荆璞张伟
东北林业大学学报 2013年7期
荆 璞 张 伟
(东北林业大学,哈尔滨,150040)
在哺乳动物被毛的研究中,有关动物换毛序的研究并不多见[1-3],而动物换毛序与环境关系的研究是动物生态学研究的重要方面。以往研究主要采用染色法、连续两年活体观察法、毛层毛长测量相结合方法及脱毛序、生毛序、长毛序的图式法等对动物的换毛序进行研究[4-7]。虽然有关松鼠东北亚种(Sciurus vulgaris manchuricus)秋季换毛期皮板颜色变化及被毛长度变化的研究已有一些报道[8],但其多为概述性研究,涉及秋季换毛期具体时间段内被毛生长程度的研究甚少。本研究以哈尔滨室内养殖的松鼠东北亚种为对象,采用不定期取皮方法对松鼠换毛序进行观察,细化换毛期内被毛各部位的总体生长规律。通过对秋季换毛过程中被毛部分性状的系统研究,加深了对松鼠东北亚种秋季换毛序、新生被毛生长情况的认知,量化了冬季被毛鳞片所占比例,旨在为哈尔滨野外条件下松鼠的换毛序、换毛时间、具体时间内的被毛生长程度提供理论参考。
1 材料与方法
1.1 材料的选取
在2011年9月20日至12月15日期间,利用不定期采样方法对30只(初生日期相同的15只雄性个体,15只雌性个体)哈尔滨室内养殖松鼠进行取样,每次采集松鼠皮3张,同时在每张松鼠皮的头部、背中部、臀部分别采集直针毛和绒毛各15根,左右耳部簇毛各3根。用以比较换毛期不同时间段内,耳部簇毛长度、各部位新生被毛长度和毛根出现无髓段比例,以此衡量松鼠东北亚种新生被毛生长速率和结束生长的比例。此外对于12月15日采集的3张松鼠皮,分别在每张背中部多采集直针毛和绒毛各5根,将在12月15日采集的所有背中部直针毛和绒毛用以作为松鼠被毛鳞片的观察。
1.2 方法
选取一小块带毛的皮板,用沸水煮至变软,用尖镊子将完整的直针毛和绒毛从皮板中拉出,用V(乙醚)∶V(95%的乙醇)=1∶1的脱脂液脱脂20 min,再用无水乙醇清洗10 min,将处理后的毛放到2~3 mm厚的无色有机载片上,在其上下覆以无机玻璃片,并用铁夹固定。置于120℃的DGG-9053A型电热恒温干燥箱中加热2 h,待样本冷却后,卸下铁夹及无机载片,此时毛附着于有机载片上,用窄透明胶带将整根毛粘起并平行贴于距毛鳞片印痕1~2 mm处,即制成毛的鳞片压膜片。
1.3 数据测量
使用精度为毫米的钢直尺测量毛的伸直长度,毛伸直长度是将毛拉直后毛尖至毛根的距离。将已制好的鳞片压膜片放置10倍物镜视野的显微镜下,观测是否毛根出现无髓段。40倍物镜视野中,利用显微测微尺测量自毛尖至毛根各鳞片类型起点与终点之间的水平距离,即某种鳞片类型的总长度。
1.4 数据处理
在换毛期不同时间段内,分别比较耳部簇毛长度和各部位新生被毛长度的生长变化。分别记录每次取样时,各部位出现无髓段新生毛的数量,统计各部位无髓段新生毛占取样新生毛总量的比例。根据测量值,整合各类鳞片类型的平均长度及所占毛的比例。所有统计分析均在SPSS17.0中进行,数据均以mean±SD表示。
2 结果与分析
2.1 皮板颜色变化
秋季换毛期被毛的长毛序为从后往前[8-9],且新生被毛完成生长的顺序为:臀部—背部—颈部—头部—体侧、后肢—前肢,其中,四肢的外侧要略先于四肢内侧完成生长(图1)。
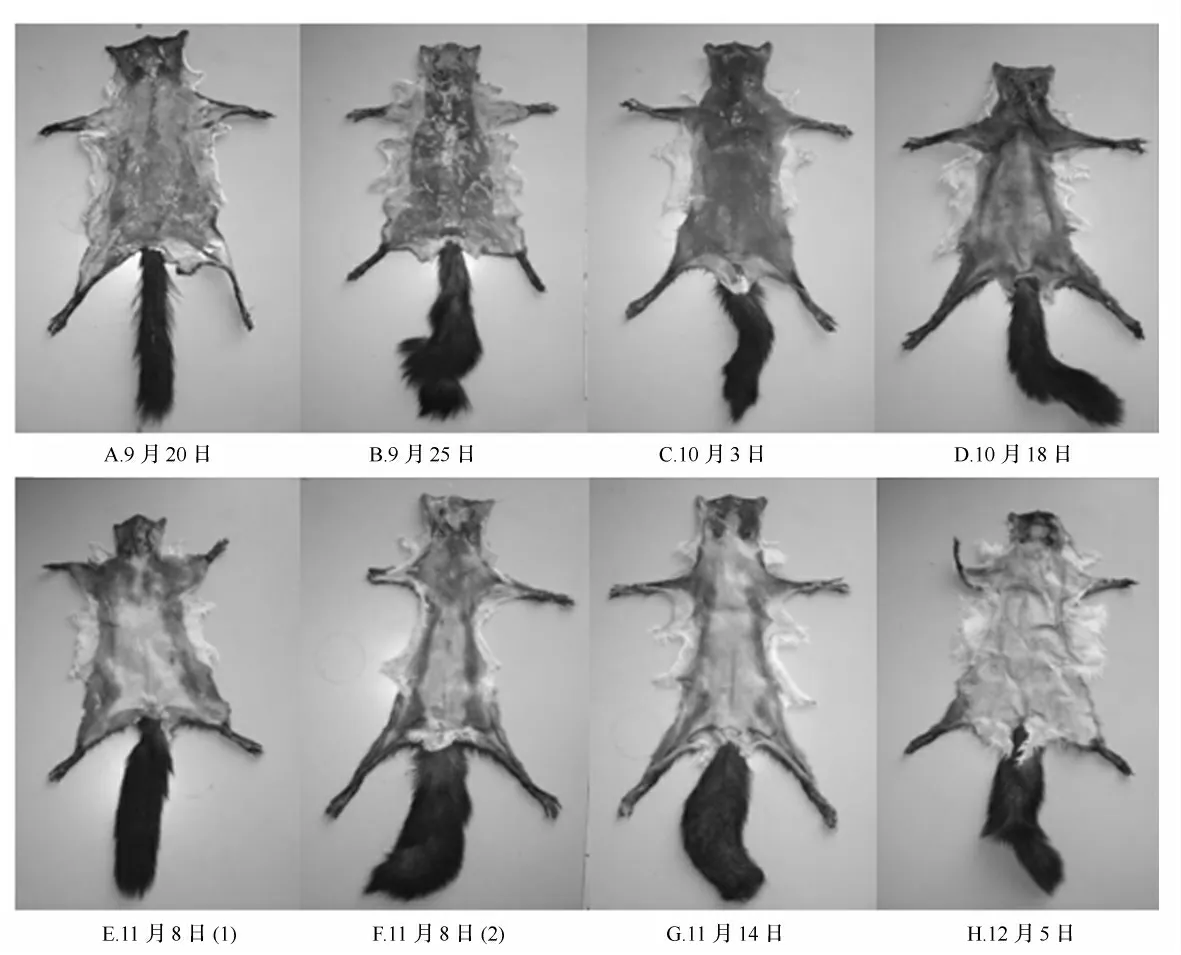
图1 秋季换毛期皮板颜色的变化
松鼠受光周期的变化启动秋季换毛,由于全身大部分皮肤内毛囊的冬毛正在孕育阶段,此时皮肤中富含黑色素的毛圆锥大量形成,皮肤呈青黑色,俗称“黑条子”(图1C)。松鼠只有已经长出冬毛的后臀部的皮肤由青黑色变成淡白色,俗称“小花板”(图1D)。松鼠正处于冬毛生长旺期,只有脊背上部、颈头部和2条前腿处的皮板呈青黑色,臀部毛已长平齐,其他大部分新生冬毛的长度为成熟冬毛长度的一半,俗称“大花板”(图1F)。松鼠仅头部和颈部两侧皮板呈青黑色,两个前腿处的毛绒尚短,全身冬毛大部分长出,只是略显短、稀,接近成熟,俗称“雁脖”(图1G)。冬季松鼠周身毛绒已经成熟,头部毛绒略欠丰厚,除头部皮板呈青黑色外,全身均为白色,俗称“黑头”。松鼠正冬季全身皮板均为白色,富有油性,毛被完全成熟,俗称“冬白板”(图1H)。
2.2 秋季换毛期被毛长度的变化
2.2.1 秋季换毛期耳部簇毛的长度变化
表1显示,室内饲养的雄性松鼠9月20日出现耳部簇毛,9月25日雌、雄个体左右耳簇毛长度为0.49~0.61 cm。随着时间的推移,耳部簇毛长度呈不断增长的趋势,12月5日簇毛长度为5.55~5.90 cm。其中雄性个体耳簇毛的出现和生长进度要略先于雌性个体。
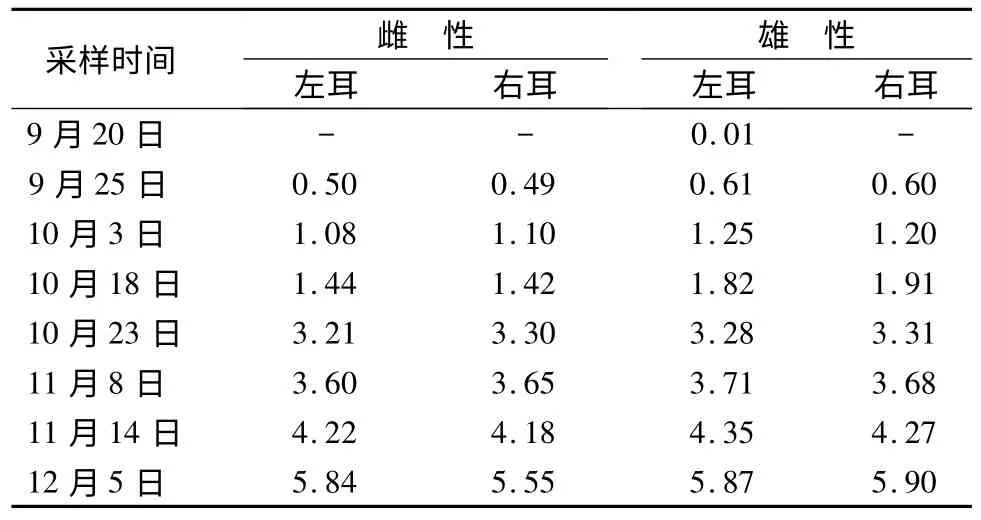
表1 不同时间段松鼠耳部簇毛平均长度 cm
2.2.2 不同采集时期各部位新生直针毛和绒毛的长度变化
从表2看出,头部、背中部、臀部直针毛和绒毛分别在10月18日、10月3日、9月25日长度最短,且各部位直针毛和绒毛的最短长度分别为(4.35±2.21)、(3.77±1.22)mm,(6.46±1.39)、(4.16±1.62)mm,(5.65±1.88)、(4.46±0.64)mm。在12月15日各部位直针毛和绒毛都达到最大伸直长度,分别为(21.32±2.56)、(13.07±1.25)mm,(26.25±3.06)、(13.88±1.76)mm,(28.07±2.37)、(15.00±1.27)mm。9月25日—11月14日比11月14日—12月15日期间内各部位直针毛和绒毛长度变化明显,前期被毛生长速度较快。在换毛期导致每个时间段内被毛长度臀部>背中部>头部的原因,主要是由于松鼠秋季换毛序的原因所致。
2.3 不同采样时期各部位直针毛和绒毛毛根出现无髓样本的比例
根据试验观察松鼠的毛都是有髓毛。由于毛在发生时,首先形成的是鳞片细胞和皮质细胞,因而毛尖端均无髓质,而当毛生长接近成熟时,首先停止分生的是髓细胞,所以毛根下部亦无髓质。因此可以根据一根毛的毛根下部是否无髓,判别这根毛是否已经结束生长[7]。由表3可见,整体上看,随着时间的推移,直针毛和绒毛毛根出现无髓段样本的比例是逐渐增长的,直至12月15日3个部位毛根无髓样本比例都为100%。头部、背中部和臀部被毛毛根出现无髓段即毛的生长完全结束的日期分别为11月8日、10月18日、10月3日,这是由于松鼠秋季换毛序是从臀部向头部的方向扩展。同一采集时间的3个部位绒毛出现毛根无髓样本的比例均大于直针毛,且11月25日臀部绒毛毛根无髓比例先达100%,说明绒毛结束生长的时期早于直针毛。
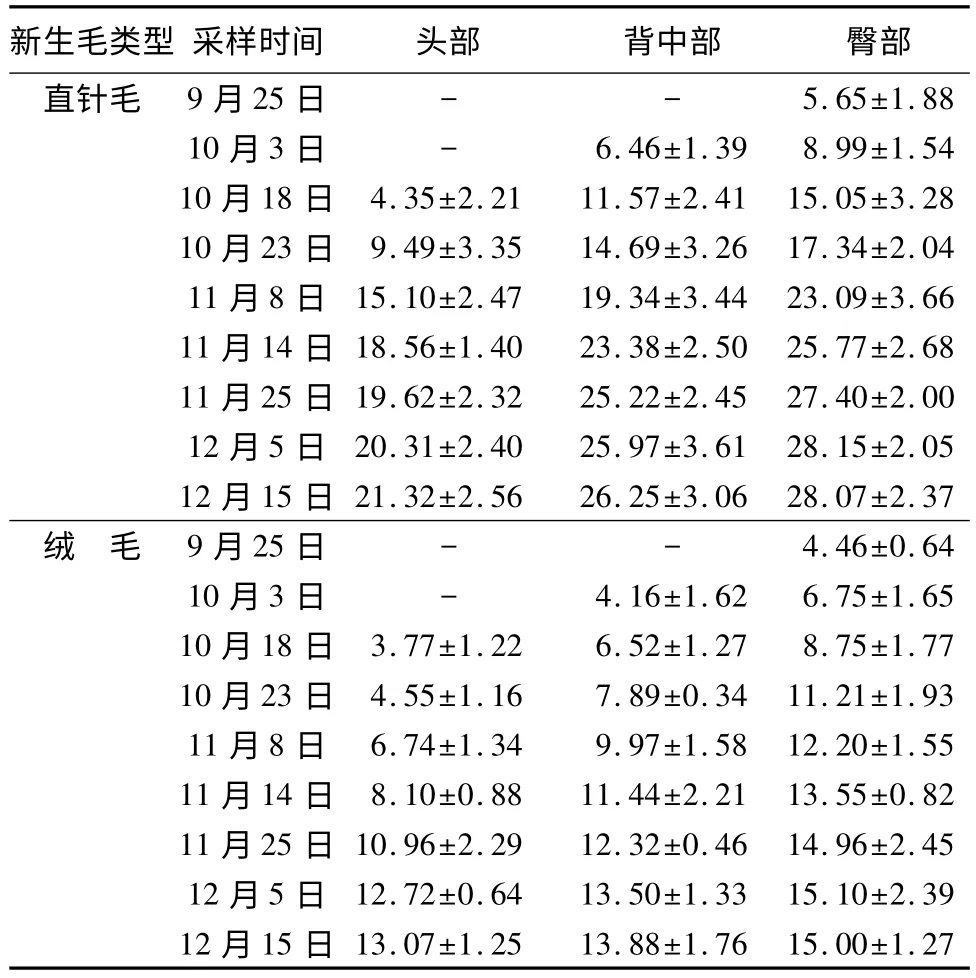
表2 不同采集日期各部位新生毛平均长度 mm

表3 各时间段新生直针毛和绒毛毛根无髓段比例%
2.4 冬季松鼠被毛背中部直针毛、绒毛鳞片类型
由表4统计结果显示:松鼠背部直针毛从毛尖到毛根的鳞片排列顺序依次为冠状型+杂波型+规则扁平型、过渡型1(由扁平型向宽杂瓣型过渡)、宽杂瓣型、过渡型2(由宽杂瓣型向长瓣型过渡)、长瓣型、类扁平型,其中冠状型+杂波型+规则扁平型鳞片长度和比例均为直针毛各鳞片类型的最大值,分别为(13293±957)μm、(44.88±3.23)%,而类扁平鳞片长度和比例皆最小,分别为(417±98)μm、(1.41±0.33)%。直针毛各鳞片类型长度和所占的比例排序为:冠状型+杂波型+规则扁平型>长瓣型>宽杂瓣型>过渡型2>过渡型1>类扁平型。背中部绒毛从毛尖到毛根的鳞片排列顺序依次为冠状型、过渡型(由冠状型向长瓣型过渡)、长瓣型、类扁平型,其中冠状型鳞片长度和比例均为绒毛各鳞片类型的最大值,分别为(6446±536)μm、(45.23±3.76)%,类扁平型鳞片长度和比例都是最小值,分别是(210±43)μm、(1.47±0.30)%。绒毛各鳞片类型长度所占的比例排序为:冠状型>长瓣型>过渡型>类扁平型。
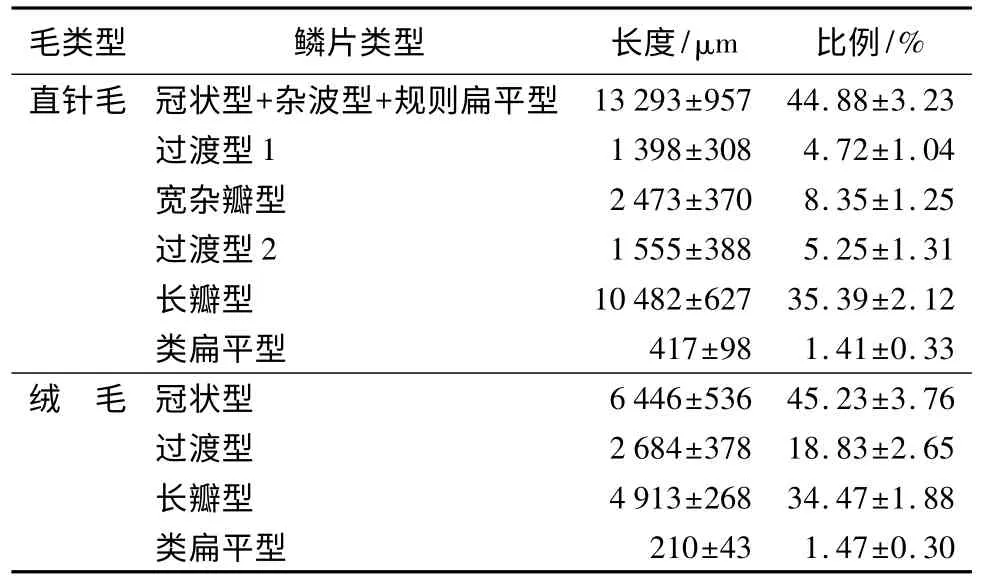
表4 冬季松鼠背中部直针毛鳞片类型、长度及其比例(从毛尖到毛根)(n=60)
3 结论与讨论
松鼠的换毛期因产地纬度变化而异,光照和温度是影响换毛的主要因素,纬度越高,换毛起始时间就越早。大兴安岭漠河地区的秋季换毛期约8月底即换冬毛[9],小兴安岭伊春地区9月初松鼠被毛尚无脱换现象[8],随地区纬度的差异有迟早之别,先后换毛起始时间相差半月之久。本研究地点为哈尔滨地区,因纬度比大小兴安岭纬度都低,所以换毛起始时间较前二者都晚。又由于试验采用室内饲养的松鼠,尽管在饲养中力求模拟野外饲养条件,但因试验条件有限,致使室内条件(光照、温度、湿度等)与室外条件略有不同,从而导致室内养殖的松鼠较野外松鼠换毛进度略慢。研究所得耳部簇毛的出现时间为9月20日,而耳部簇毛的出现是作为松鼠秋季换毛起始时的标记,这也是松鼠作为季节性换毛动物所特有的换毛特点。
松鼠东北亚种不属于雌、雄二型性动物,换毛过程中雌、雄个体皮板颜色、毛长度无明显的差异,而唯有耳簇毛雄性比雌性出现的时间略早,表明松鼠雄性比雌性换毛的起始时间略早,因换毛序包括脱毛序、生毛序和长毛序,所以松鼠雌、雄性差异并不影响其换毛期新生冬毛长度的研究结果。试验中之所以能及时地在不同的时间段上观测到松鼠各部位被毛的长度和出现结束生长的被毛的比例程度,取决于松鼠换毛序的特点。秋季换毛期松鼠的新毛在旧毛层下局部生出,边生边长向前萌发扩展,就是说,生毛序、长毛序与脱毛序三者是在一个区段上相重叠,松鼠东北亚种换毛序既不像岩松鼠那样三者几乎在一条线上进行,又不似黄鼬那般三者几乎全身性地在大片面积上相重叠[7]。
总体上看,冬季松鼠背中部直针毛和绒毛的冠状型、过渡型、长瓣型、类扁平型鳞片所占毛长度的相对比例大致相同。其中对于直针毛将冠状型、杂波型、规则扁平型合为一起观测是因为毛尖处的冠状型鳞片和薄的扁平型鳞片的局部脱落都是为了防止被毛离体而起到的机体保护功能,而杂波型鳞片为规则扁平型鳞片受到磨损后所形成的。因此,被毛受到或多或少的磨损后无法将3种鳞片进行具体的划分。松鼠直针毛和绒毛的长瓣型鳞片主要分布在近毛根处,所占毛鳞片的比例都在32%~37%范围内,而江獭直针毛和绒毛的长瓣型鳞片比例分别为60%、69%[7],是松鼠的2倍,那是因为江獭是半水生哺乳动物,长瓣型鳞片所占毛鳞片比例越大,就会使针毛与针毛间、针毛与绒毛间的相对摩擦力增加,从而导致体被层的稳定程度大大加强,加之江獭毛短毛密的特点,因此使其更好地适应寒冷的江河生境。相比较而言,松鼠作为陆生的哺乳动物,生境环境并非极寒冷,体被层无需过度的稳定与保温,因此直针毛和绒毛的长瓣型鳞片所占比例无需过大。
虽然本研究已对换毛期新生毛部分性状做量化研究,但关于换毛期被毛保温性能的变化趋势及影响保温性能的生态因子、体能参数(如皮下脂肪和皮张厚度)及被毛结构变化等研究并未涉及,有待在以后的工作中继续深化。
[1]Hoffmann K.The influence of photoperiod and melatonin on testis size,body weight,and pelage colour in the Djungarian hamster(Phodopus sungorus)[J].Journal of Comparative Physiology A:Neuroethology,Sensory,Neural,and Behavioral Physiology,1973,85(3):267-282.
[2]Allain D,Rougeot J.Induction of autumn moult in mink(Mustela vison Peale and Beauvois)with melatonin[J].Reproduction,Nutrition Développement,1980,20(1A):197-201.
[3]Daniel R G,Jemison L A,Pendleton G W.Molting phenology of harbor seals on Tugidak Island,Alaska[J].Marine Mammal Science,2003,19(1):128-140.
[4]杨圭璋.黄鼬季节性换毛的规律[J].毛皮动物饲养,1980(2):6-8.
[5]朱靖,王家义.黃鼬(Mustela sibirica Pall.)换毛序[J].动物学杂志,1960(7):293-295.
[6]朱靖.兽类换毛序研究方法的讨论:附毛层毛长测定法[J].动物学杂志,1960(6):241-244.
[7]杨树勋,潘文石.岩松鼠换毛序[J].动物学杂志,1985(4):6-8.
[8]朱靖,杨树勋,王家义.灰鼠(Sciurus vulgaris Linné)换毛序[J].动物学杂志,1960(6):244-249.
[9]景松岩,张伟,徐艳春.毛皮学[M].哈尔滨:东北林业大学出版社,1993:41-84
猜你喜欢
作文小学中年级(2022年5期)2022-06-02
作文大王·低年级(2020年4期)2020-04-19
阅读与作文(小学低年级版)(2019年12期)2019-12-26
中成药(2018年10期)2018-10-26
短篇小说(2017年1期)2017-09-22
儿童与健康(幼儿教师参考)(2017年5期)2017-06-05
短篇小说(原创版)(2017年1期)2017-02-24
国际妇产科学杂志(2016年2期)2016-06-16
中国塑料(2015年8期)2015-10-14
