
杜氏盐藻糖原合成酶激酶3 cDNA片段的克隆及其在鞭毛再生中的功能*
2013-11-21 01:36毛丽红李庆华王瑞莉龚方华薛乐勋1
郑州大学学报(医学版) 2013年2期
毛丽红,李庆华,韩 康,王瑞莉,龚方华,薛乐勋1,#
1)郑州大学生物工程系细胞生物学研究室 郑州 450001 2)郑州大学第一附属医院细胞生物学研究室 郑州 450052
糖原合成酶激酶3(glycogen synthase kinase 3,GSK3)是一种多功能的丝/苏氨酸蛋白激酶,它可以抑制糖原合成酶的活性,导致胰岛素抵抗,从而调节体内糖代谢水平[1]。GSK3还在Wnt信号通路中发挥重要作用,它通过与APC、Axin、CK1和β-catenin形成复合体,使β-catenin发生磷酸化,从而影响下游转录因子[2]。它还参与Hedgehog、Notch、mTOR等其他信号转导途径,并对各种细胞过程进行调控,包括细胞增殖、细胞凋亡、神经元功能及胚胎发育等[3-4]。鞭毛是广泛存在于多种细胞表面、结构保守的细胞器,它在介导细胞运动、感知外界环境及胞内信号传递的过程中发挥重要作用。鞭毛的组装是一个双向的、动态的组装过程,非常有利于鞭毛维持在合适的长度[5]。然而一旦这个过程发生异常,鞭毛出现缺陷或丢失,就有可能引发多种疾病,如多囊肾、癌症等。因此研究鞭毛长度调控机制对了解这些疾病的发生具有重要的意义。目前已有研究[6]证明,采用GSK3的抑制剂LiCl能够使衣藻鞭毛延长,表明衣藻中GSK3对鞭毛组装和维持鞭毛长度具有一定的调控作用。杜氏盐藻作为一种高度耐盐的单细胞绿藻,具有一对等长的鞭毛,且易于观察研究。该研究克隆杜氏盐藻GSK3 cDNA序列,并通过实时荧光定量的方法初步探讨杜氏盐藻GSK3与鞭毛的关系。
1 材料与方法
1.1藻株与培养杜氏盐藻UTEX LB-1644购自美国德克萨斯州大学,采用改良型液体培养基于26 ℃、4 500 Lux光照强度下在HPG光照培养箱中培养,光暗周期各为12 h。
1.2主要试剂限制性内切酶、3’RACE试剂盒、T4 DNA连接酶和pMD18-T载体均购自TaKaRa公司,逆转录试剂盒为Fermentas公司产品,实时荧光定量染料购自全式金公司。
1.3杜氏盐藻GSK3cDNA片段的克隆与鉴定
1.3.1 总cDNA模板的制备 取对数生长期盐藻细胞10 mL,1 000 r/min离心10 min收集细胞,培养基洗涤2遍后离心收集细胞于无RNase离心管中。Trizol裂解法提取盐藻总RNA,用琼脂糖凝胶电泳和分光光度计检测所提RNA的质量和浓度。最后按逆转录试剂盒说明将RNA逆转录成cDNA模板。
1.3.2 GSK3基因片段的扩增 根据转录组测序得到的2个短片段分别设计上下游引物:上游 5’-TGG TGGATGGGGACCTCA-3’(18 bp),下游 5’-GCCGCC GAAGATGAGCTC-3’(18 bp)。PCR扩增两片段间的序列,反应条件:94 ℃预变性3 min;94 ℃变性30 s,50~60 ℃退火30 s,72 ℃延伸1 min;反应30个循环后72 ℃延伸10 min,4 ℃终止反应。PCR产物经琼脂糖凝胶电泳分离后胶回收,回收片段与pMD18-T载体16 ℃过夜连接。然后转化E.coliDH5α进行蓝白斑筛选,挑取阳性单克隆扩大培养并提取质粒,EcoRⅠ、HindⅢ双酶切鉴定,取插入片段大小正确的单克隆菌液送测序,进一步进行鉴定。
1.3.3 3’RACE法扩增GSK3 3’端序列 根据已扩增的GSK3片段设计3’RACE上游引物:外侧5’-CCTGGTGCTGGAGTTCGT-3’(18 bp),内侧5’-ACTA
CACTGCTGCCATTGAT-3’(20 bp)。下游引物为3’RACE试剂盒自带的引物:外侧 5’-TACCGTCGTTC
CACTAGTGATTT-3’(23 bp)和内侧 5’-CGCGGATCC TCCACTAGTGATTTCACTATAGG-3’(32 bp)。提取盐藻总RNA,使用3’RACE 接头引物进行逆转录制备模板,按试剂盒说明上的程序进行巢式PCR反应。PCR产物经琼脂糖凝胶电泳、胶回收、连接及转化后,提取质粒,EcoRⅠ、HindⅢ双酶切鉴定正确的单克隆菌样送测序。
1.3.4 利用抑制消减杂交模板扩增5’端序列 将Tester cDNA模板PCR 引物5’-AAGCAGTGGTATCA ACGCAGAGTAC-3’(25 bp)作为上游引物。根据已知片段设计的特异性引物:外侧5’-ACTCTGCCACC ACGCAGC-3’(18 bp),内侧5’-AGGAGCACGGTAG TAACG-3’(18 bp),作为下游引物。以Tester cDNA为模板,按常规反应体系和程序进行巢式PCR,所得片段连入载体后,经蓝白斑筛选,质粒提取,EcoRⅠ、HindⅢ双酶切鉴定,最后送测序。
1.3.5 GSK3 cDNA总长的验证 根据拼接后cDNA片段两端序列设计引物,PCR扩增得到GSK3 cDNA总长,回收后连入pMD18-T载体,质粒用BamHⅠ、HindⅢ酶切鉴定后送测序。
1.4实时荧光定量PCR法检测GSK3在鞭毛再生中的变化采用pH休克法脱去杜氏盐藻鞭毛[7],在去鞭毛后的300 min内,每隔30 min取一次盐藻,将其收集于无RNase离心管中,加Trizol裂解提取RNA,逆转录为cDNA。设计引物:上游5’-GGGGCAAACAAGCAGACG-3’(18 bp),下游5’-CG GATGGTCCACCATCTT-3’(18 bp),以GAPDH作为内参,未脱鞭毛盐藻作为对照,进行实时荧光定量PCR,导出数据计算2-ΔΔCt值进行定量分析,其中Ct为扩增产物达到设定阈值所经历的循环数。
2 结果
2.1杜氏盐藻GSK3cDNA片段结果分析
2.1.1 GSK3片段的扩增及鉴定 PCR扩增得到一段长为605 bp的序列(图1A),大小与推测值相符。胶回收后与pMD18-T载体连接,转化后提取的质粒进行双酶切鉴定,结果见图1B。
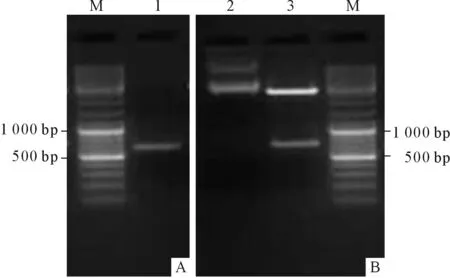
图1 GSK3片段的扩增和鉴定结果
A:PCR结果;B:双酶切鉴定结果;M:Marker;1:GSK3;2:pMD18-T-GSK3;3:pMD18-T和GSK3。
2.1.2 3’RACE和5’端cDNA序列扩增结果 3’RACE PCR扩增得到长为1 195 bp片段(图2A),测序结果显示含完整的polyA尾,回收片段与pMD18-T载体连接,质粒经双酶切鉴定,结果见图2B。5’端PCR扩增得到899 bp片段(图3A),酶切鉴定结果见图3B。
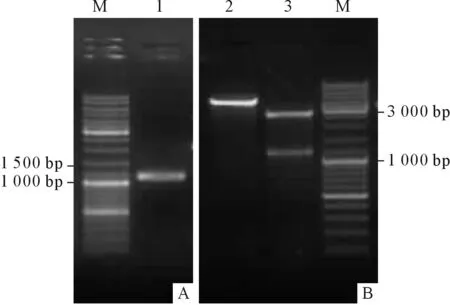
图2 3’RACE扩增和鉴定结果
A:PCR结果;B:双酶切鉴定结果;M:Marker;1:3’RACE PCR产物;2:pMD18-T-3’GSK3;3:pMD18-T和3’GSK3。

图3 5’端cDNA扩增和鉴定结果
A:PCR结果;B:双酶切鉴定结果;M:Marker;1:5’端PCR产物;2:pMD18-T-5’GSK3;3:pMD18-T和5’GSK3。
2.1.3 GSK3 cDNA总长序列分析结果 拼接后得到GSK3 cDNA总长为2 116 bp(图4),其中包括737 bp的3’UTR和1 158 bp的开放阅读框,编码386个氨基酸。氨基酸序列经比对后与莱茵衣藻的同源性为86%。

图4 GSK3 cDNA总长的扩增结果
2.2实时荧光定量PCR结果与未脱鞭毛组相比,在去鞭毛后的300 min内GSK3基因转录量明显升高,并在120 min时达到最高水平(图5)。

图5 GSK3实时荧光定量PCR结果
3 讨论
杜氏盐藻是一种无细胞壁的单细胞真核绿藻,具有一对等长的鞭毛[8],长度约为13 μm,鞭毛具有典型的“9+2”结构。因其结构简单,生长周期短,鞭毛形态易于观察,所以非常适合作为研究鞭毛/纤毛长度调控机制的模型生物。然而,由于杜氏盐藻的基因组序列尚未公布,且已得到全序列的鞭毛相关基因较少,因此给鞭毛调控机制和功能的研究带来了诸多不便。该研究通过扩增GSK3 cDNA序列,得到了完整的开放阅读框,编码386个氨基酸,经在NCBI上比对发现其氨基酸序列与衣藻、团藻等具有较高的同源性,说明GSK3在不同物种中高度保守,而这种高保守性也正是GSK3维持自身激酶活性所必需的。
一直以来,关于GSK3的研究层出不穷,作为一个多功能基因,它不仅是维持体内糖代谢的关键酶,而且还是多种细胞信号通路的关键因子。GSK3还可以通过使微管相关蛋白Tau、MAP-1B和APC发生磷酸化来调控微管的动力学作用,从而影响轴索的生长[9]。另外,GSK3可以通过分解驱动蛋白1,进而选择性地抑制顺向轴丝运输及其与货物蛋白间的相互作用[10]。虽然这些研究已经证明GSK3对维持微管的稳定性及鞭毛内运输具有一定的调控作用,但对于它在鞭毛生长中的作用以及长度调控的机制目前尚不清楚。所以,作者通过实时荧光定量PCR方法研究了GSK3在转录水平上与鞭毛再生的关系,结果显示GSK3的转录量明显升高,甚至比对照组高20多倍,表明GSK3在鞭毛再生中发挥重要作用。虽然已有研究[6]表明,GSK3的活性形式在鞭毛内大量存在且其活性抑制剂LiCl能够使衣藻鞭毛延长。但是,在杜氏盐藻中,锂处理后鞭毛长度并未出现明显的延伸,因此,作者推测杜氏盐藻GSK3可能不参与鞭毛长度的维持,只是在鞭毛生长过程中发挥调控作用。另外,GSK3是否通过参与鞭毛内某种信号通路实现对鞭毛生长的调控以及它在鞭毛再生中是否还发挥其他未知的功能,这还需要进一步的研究才能证实。
[1] Lee J, Kim MS. The role of GSK3 in glucose homeostasis and the development of insulin resistance[J]. Diabetes Res Clin Pract, 2007,77(Suppl 1):S49
[2] MacDonald BT, Tamai K, He X. Wnt/beta-catenin signaling: components, mechanisms, and diseases[J]. Dev Cell, 2009,17(1):9
[3] Forde JE, Dale TC. Glycogen synthase kinase 3: a key regulator of cellular fate[J]. Cell Mol Life Sci, 2007,64(15):1930
[4] Rayasam GV, Tulasi VK, Sodhi R, et al. Glycogen synthase kinase 3: more than a namesake[J]. Br J Pharmacol, 2009,156(6):885
[5] Pedersen LB, Rosenbaum JL. Intraflagellar transport (IFT) role in ciliary assembly, resorption and signaling[J]. Curr Top Dev Biol, 2008,85:23
[6] Wilson NF, Lefebvre PA. Regulation of flagellar assembly by glycogen synthase kinase 3 in Chlamydomonas reinhardtii[J]. Eukaryot Cell, 2004,3(5):1307
[7] Cheshire JL, Evans JH, Keller LR. Ca2+signaling in the Chlamydomonas flagellar regeneration system: cellular and molecular responses[J]. J Cell Sci, 1994,107(Pt 9):2491
[8] Oren A. A hundred years ofDunaliellaresearch: 1905-2005[J]. Saline Systems, 2005,1:2
[10]LaPointe NE, Morfini G, Pigino G, et al. The amino terminus of tau inhibits kinesin-dependent axonal transport: implications for filament toxicity[J]. J Neurosic Res, 2009, 87(2):440
猜你喜欢
广东药科大学学报(2022年3期)2023-01-04
智慧农业导刊(2022年5期)2022-11-24
生物学通报(2022年1期)2022-11-22
——一道江苏高考题的奥秘解读和拓展
中学生物学(2022年7期)2022-09-07
成都医学院学报(2022年4期)2022-08-19
宝鸡文理学院学报(社会科学版)(2021年4期)2021-12-04
江西农业学报(2021年4期)2021-04-20
三农资讯半月报(2020年11期)2020-06-21
大观(2019年1期)2019-06-28
生物学教学(2019年3期)2019-03-22
