
北京市几种绿化树种的光合特性及生态效益比较
2014-03-26 12:21胡耀升么旭阳刘艳红
西北农林科技大学学报(自然科学版) 2014年10期
胡耀升,么旭阳,刘艳红
(北京林业大学 森林培育与保护省部共建教育部重点实验室,北京 100083)
随着北京市经济的不断发展和人民生活水平的日益提高,城市生态环境越来越受到重视。城市绿化植物作为城市生态环境的重要组成部分,是优化城市环境、保证城市生态系统稳定的必要组成,在新时期建设环境友好、可持续发展的生态城市中具有非常重要的作用。近年随着城市化发展步伐的加快,引进植物在北京市所占的比例越来越大[1-2]。适度引进优良种质资源,对于丰富植物品种的生态多样性、遗传多样性和种质优异性十分重要[3-4]。而在构建城市森林的过程中,乡土植物因具有较强的适应性、抗逆性、抗病虫害能力以及易于养护管理等特点[5],同样不能被忽视。如何正确选择、配置树种以及科学引种,已成为城市森林建设过程中迫切需要解决的问题。
面对现在的城市用地紧张、绿化用地面积不断缩小的窘境,合理利用植物自身的特性,发挥其最大的生态功能就显得尤为重要。然而,以往研究过分强调美化景观,对生态效益只定性描述[6-8],缺乏对城市森林生态效益的定量评价。而对植物生态功能进行系统的量化研究,是建设城市森林必须解决的问题,它不但是研究城市绿化生态效益的一个重要基础,同时也可作为合理配置植物的主要依据。唐鸣放等[9]对墙面绿化、屋顶绿化的降温效应进行了测定,结果表明,在建筑物墙面和屋顶进行垂直绿化可以显著降低室内温度,并改善周围小气候。徐玮玮等[10]研究了扬州古运河风光带19种绿地组成植物的固碳释氧效应,并对各树种的日固碳释氧能力进行聚类分析,提出了由不同树种组成的绿地配置类型。本研究对城市环境下的4种引进树种和4种乡土树种的光合特性及其与环境因子的关系进行了分析,并对其固碳释氧和增湿降温量等进行了计算,首次从生态效益方面评价引进效果,同时将引进植物与乡土植物进行了对比分析,旨在为北京市城市绿化树种的合理选择与配置提供科学依据和参考。
1 材料与方法
1.1 研究地区概况
研究地位于北京市西北部的海淀区。地理坐标为北纬 39°53′~40°09′,东经116°03′~116°23′。东与西城区、 朝阳区相邻, 南与宣武区、 丰台区毗连,西与石景山、门头沟区交界,北与昌平区接壤,区域面积 430.8 km2。 全区地处华北平原的北部边缘地带,系古代永定河冲积的一部分。地势西高东低,西部为海拔 100 m以上的山地,东部和南部为海拔 50 m左右的平原。全区气候属温带湿润季风气候,春秋干旱;冬季寒冷干燥,盛行西北风;夏季高温多雨,盛行东南风。年均气温 12.5 ℃,年日照2 662 h,无霜期 211 d。年平均降水量 628.9 mm,主要集中于夏季的 6-8 月。
1.2 供试树种
选取海淀区东升八家郊野公园、清华大学、北京林业大学等地种植的臭椿、火炬树、刺槐等8种在北京市绿化中应用较多、长势较好的木本植物作为研究对象,具体见表1。
1.3 光合生理生态因子的测定
于树木生长旺季的7-9月份每月选择5~8 d,在晴朗、无风的天气情况下,利用Li-6400便携式光合仪(美国LI-COR公司生产)测定各树种的光合生理生态因子。从06:00-18:00,每隔1 h测量1次;测量时每个树种选择3~4株具有代表性的个体,每个个体选择3片向阳且健康的叶片进行测定,每片叶记录3个瞬时净光合速率值(Pn,μmol/(m2·s))和蒸腾速率值(Tr,mmol/(m2·s))。同时对其光合有效辐射(PAR,μmol/(m2·s))、气温(Ta,℃)、气孔导度(Gs,mol/(m2·s))、胞间二氧化碳浓度(Ci,μmol/mol)、大气二氧化碳浓度(Ca,μmol/mol)、大气水气压亏缺(VpdL,kPa)、大气相对湿度(RH,%)等生理生态因子进行同步测定。
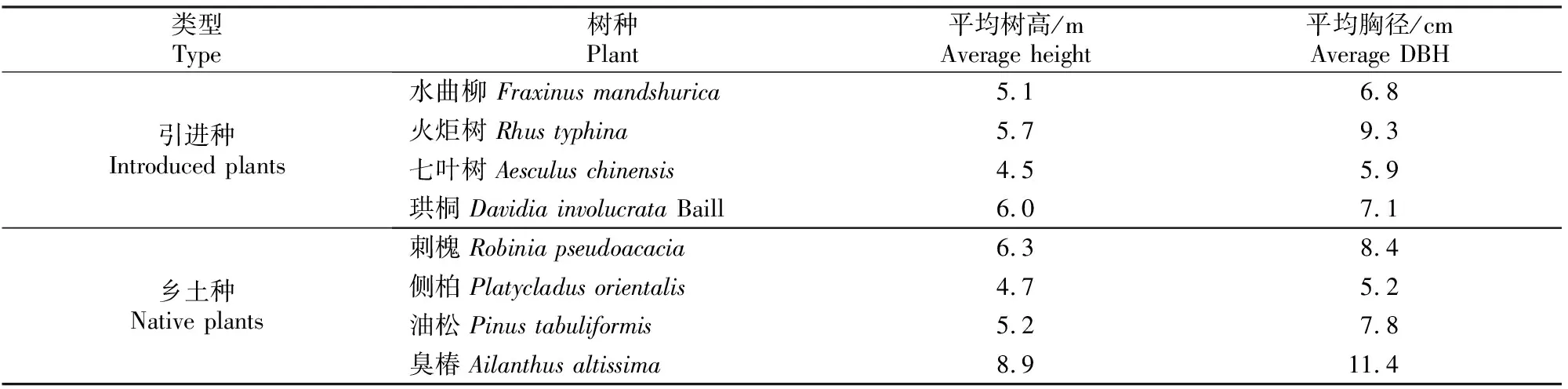
表1 8种绿化树种的基本信息
1.4 固碳释氧量和增湿降温量的计算
1.4.1 固碳释氧量 根据测定的净光合速率计算供试树种的当日同化总量[11],计算公式为:
P=∑[(Pi+1+Pi)÷2×(ti+1-ti)×3 600÷1 000]。
式中:P为测定日的同化总量,单位是mmol/(m2·d);Pi为初测时间点的瞬时光合作用速率,Pi+1为下一时间点测定的瞬时光合作用速率,两者的单位都为μmol/(m2·s);ti为初测时间点的瞬时时间,ti+1为下一时间点测定的时间;i为测试次数。
根据光合作用的反应方程式:CO2+4H2O→CH2O+3H2O+O2,依据下述公式,可计算出日固定CO2的质量和日释放O2的质量,单位均是g/(m2·d)。
日固定CO2质量的计算公式为:
WCO2=P×44÷1 000;
日释放O2质量的计算公式为:
WO2=P×32÷1 000。
1.4.2 增湿降温量 根据测定的蒸腾速率计算供试树种的日蒸腾总量[12],计算公式为:
E=∑[(ei+1+ei)÷2×(ti+1-ti)×3 600÷1 000]。
式中:E为日蒸腾总量,单位是mol/(m2·d);ei为初测时间点的瞬时蒸腾作用速率,ei+1为下一时间点测定的瞬时蒸腾作用速率,两者的单位都为mmol/(m2·s);ti为初测时间点的瞬时时间,ti+1为下一时间点的测定时间;i为测试次数。
日释水总量(WH2O),也就是日增湿量,单位是kg/(m2·d),其计算公式为:
WH2O=E×18。
根据蒸腾吸热的原理,计算出单位叶面积水分蒸腾吸收的热量,进而计算出因蒸腾吸热所导致的气温下降值[10]。
水分蒸腾吸收热量的计算公式为:
Q=WH2O×L×4.18。
式中:Q为日蒸腾吸热量,单位为J/(m2·d);L为蒸发耗热系数,L=597-0.57×T,其中T为测定日的温度。由此可计算出各种植物叶片在测定日吸收热量的值。
又因植物日蒸腾吸热量Q取自周围 1 000 m3的空气柱(10 m2×100 m),可使该气柱温度下降。 故气温下降值(以下简称降温值)[13]用下式计算:
ΔT=Q÷PC。
式中:ΔT为降温值(℃),PC为空气容积热容量,取值为1 256 J/(m3·h)。
1.5 数据统计分析
试验数据均采用Excel 2007软件及SPSS18.0软件进行分析处理,并进行One-way ANOVA分析,同时以净光合速率(Pn)为因变量,以气孔导度(Gs)、胞间二氧化碳浓度(Ci)、蒸腾速率(Tr)、大气水气压亏缺(VpdL)、气温(Ta)、大气相对湿度(RH)、光合有效辐射(PAR)为自变量进行多元逐步回归分析。
2 结果与分析
2.1 北京市4个引进树种和4个乡土树种净光合速率的日变化
植物生活在特定环境中,并与环境之间不断地进行着能量和物质交换。由于光照、温度和水分等环境因子在一天中发生明显的变化,植物的光合速率也呈现出相应的变化规律[14]。
由图1可见,在树木生长旺季,4个引进树种的光合速率日变化均呈双峰曲线,随着光合有效辐射和温度的升高,其Pn值逐渐增大,水曲柳、七叶树、火炬树在上午10:00左右达到第1个峰值(Pn分别为9.64,9.69,15.68 μmol/(m2·s)),珙桐在上午11:00左右达到第1个峰值(Pn为9.44 μmol/(m2·s));水曲柳在中午12:00出现第2次峰值(Pn为9.23 μmol/(m2·s)),火炬树和珙桐在中午14:00左右出现第2次峰值(Pn分别为14.96,9.67 μmol/(m2·s)),七叶树在中午13:00左右出现第2次峰值(Pn为8.36 μmol/(m2·s));中午12:00-13:00,温度和光合有效辐射达到最大值,植物的气孔导度开始下降,4个引进树种的Pn值均下降,出现光合“午休”现象。14:00以后,光合有效辐射逐渐下降,各树种Pn下降,且下降幅度较大,到18:00下降至全天最低值。火炬树的Pn与七叶树有显著差异(P<0.05),而其他各树种的Pn均无显著差异(P>0.05)。
由图2可看出,4个乡土树种中,臭椿和油松的Pn呈单峰曲线,刺槐和侧柏的Pn呈双峰曲线。臭椿的Pn明显高于其他乡土植物。臭椿、油松在中午12:00左右Pn达到峰值(20.10,5.88 μmol/(m2·s));刺槐、侧柏在上午10:00左右达到第1个峰值(Pn分别为14.60,7.47 μmol/(m2·s)),在下午14:00左右达到第2个峰值(Pn分别为13.70,8.71 μmol/(m2·s))。臭椿的Pn与油松有显著差异(P<0.05),而其他各树种间Pn均无显著差异(P>0.05)。
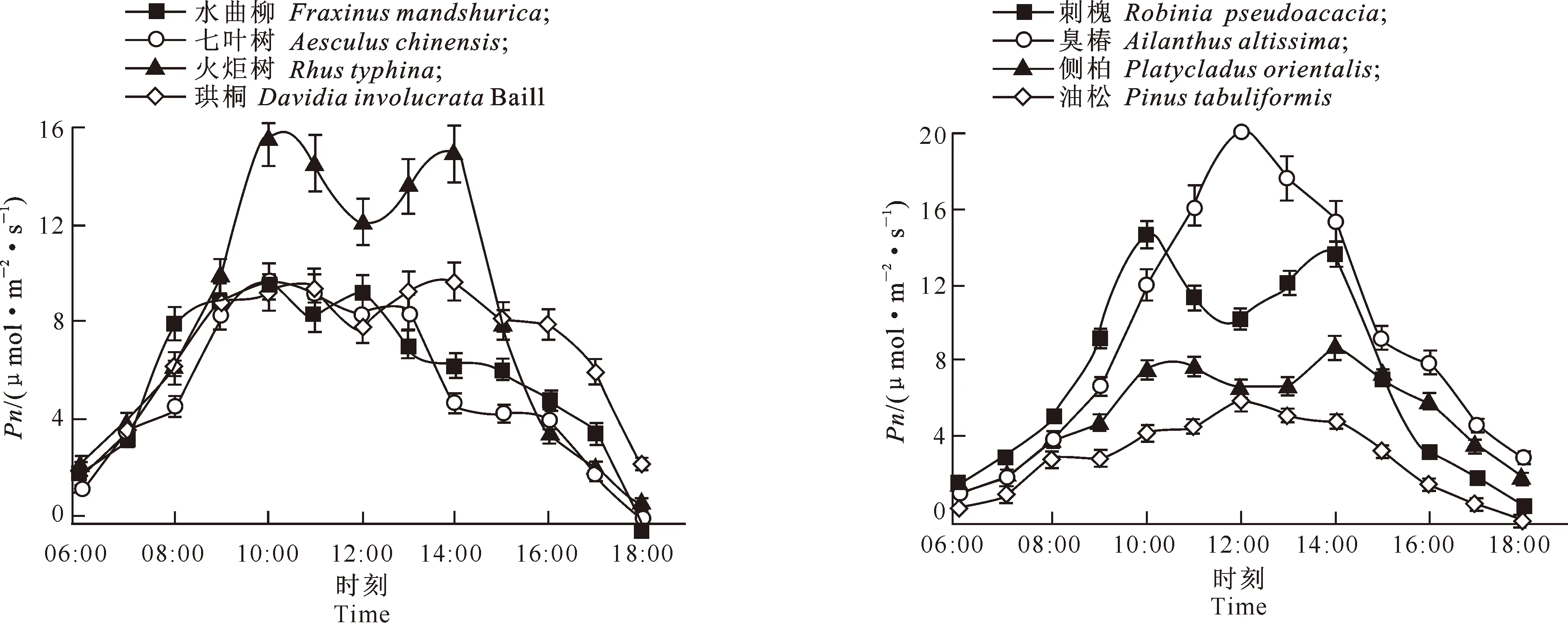
图1 北京市4种引进树种净光合速率(Pn)的日变化特征
2.2 北京市8个绿化树种净光合速率与环境因子之间的关系
植物的Pn不仅与自身特性有关,而且还受外界环境因子的影响。环境因子具有联系性和变动性,一天中光强、温度、大气湿度、CO2浓度等都不断变化,而且处于相互依赖的关系中,任何一个因子的变化都会牵动其他因子的改变,它们对植物的作用是综合性的[15]。
本研究以4个引进树种和4个乡土树种的Pn为因变量,以气孔导度(Gs)、胞间二氧化碳浓度(Ci)、蒸腾速率(Tr)、大气水气压亏缺(VpdL)、气温(Ta)、大气相对湿度(RH)、光合有效辐射(PAR)等7个生理生态因子为自变量进行拟合,确定净光合速率与环境因子关系的优化模型,结果见表2。
拟合结果表明,各模型中残差值分布是随机的,没有出现趋势性,表明回归模型有效。由表2可见,4个引进树种的复相关系数除七叶树外均大于0.95,说明回归方程拟合程度较好。从各模型的偏回归系数来看,胞间CO2浓度、气孔导度和光合有效辐射是影响Pn的主要因子。在4个乡土树种中,油松拟合度相对较差,R值仅为0.480;气孔导度、胞间CO2浓度及光合有效辐射是影响臭椿Pn的主要因子;刺槐的Pn与光合有效辐射呈正相关,与大气相对湿度呈负相关;侧柏的Pn与气孔导度呈正相关,与胞间CO2浓度呈负相关。
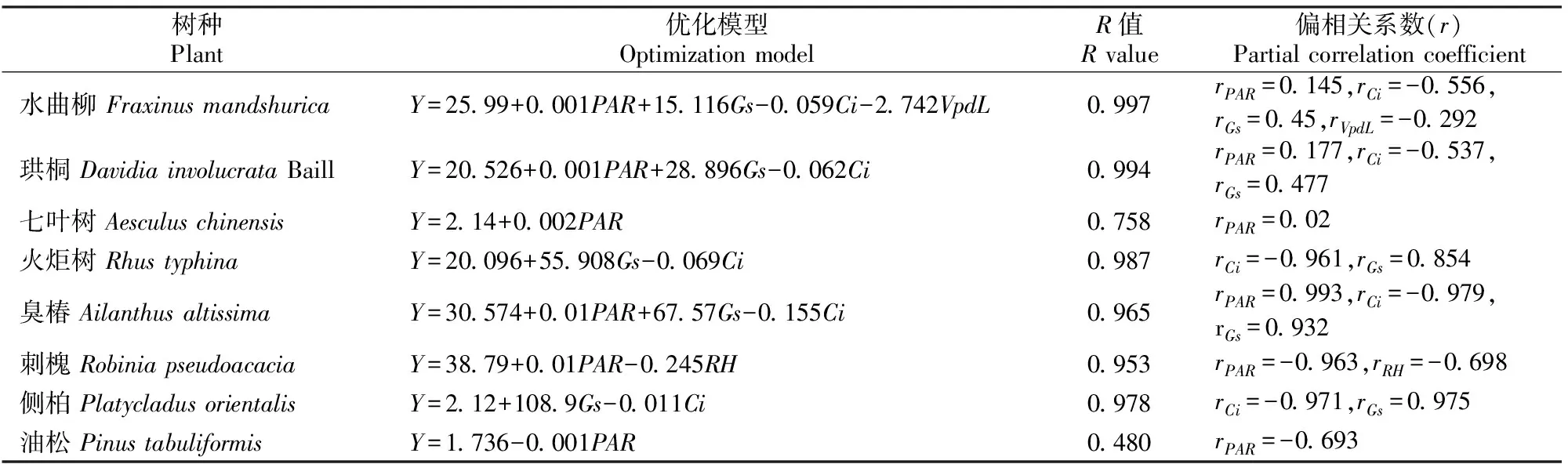
表2 北京市8个绿化树种净光合速率(Y)与环境因子(Xi)关系的优化模型
2.3 北京市8个绿化树种的固碳释氧能力
植物通过光合作用进行固碳释氧,对改善城市空气质量、实现城市生态系统良性循环发挥着重要作用[16]。
由表3可知,臭椿、火炬树的固碳释氧量较高,固碳量分别为19.36和16.99 g/m2,释氧量分别为14.08和12.36 g/m2;而七叶树、油松的固碳释氧能力均较弱,其中油松的固碳、释氧量分别为5.97和4.34 g/m2。乡土树种中,固碳释氧能力最强的是臭椿,居中的是刺槐、侧柏,较弱的为油松,其中臭椿的固碳量约是油松的4倍。引进树种的固碳释氧能力以火炬树最强,较弱的为七叶树,珙桐和水曲柳居中。就各树种固碳量而言,臭椿与七叶树、油松有显著差异(P<0.05),火炬树与油松有显著差异(P<0.05),而其他树种间的固碳量均无显著差异(P>0.05)。
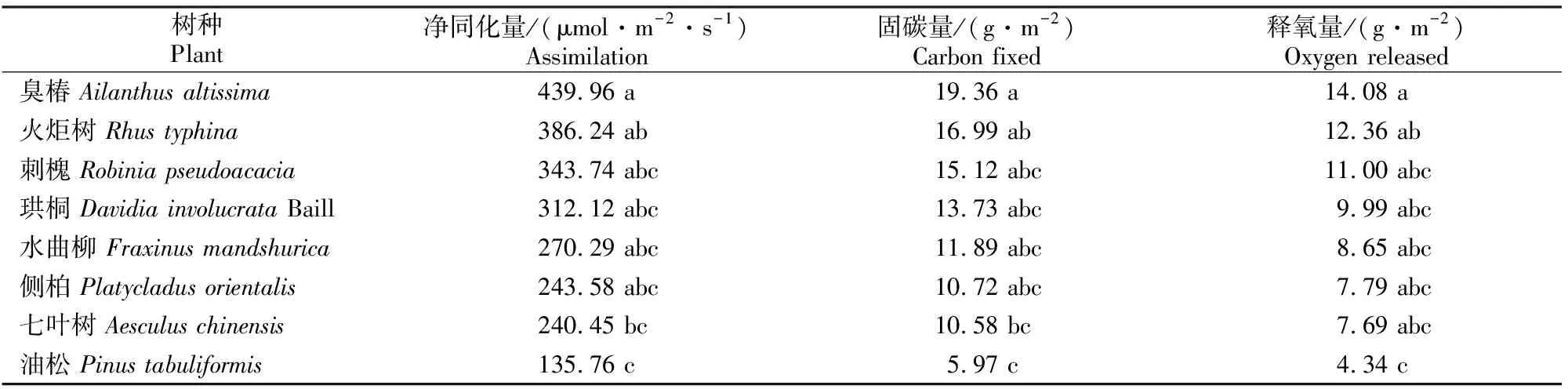
表3 北京市8个绿化树种单位叶面积的日固碳释氧量
2.4 北京市8个绿化树种的降温增湿能力
树木通过树冠减少太阳辐射和水分的蒸发,从而起到降温增湿的效果,蒸腾速率的高低直接影响着日蒸腾总量和日蒸腾吸热量的大小[17]。北京市8个绿化树种单位叶面积的日降温增湿量见表4。
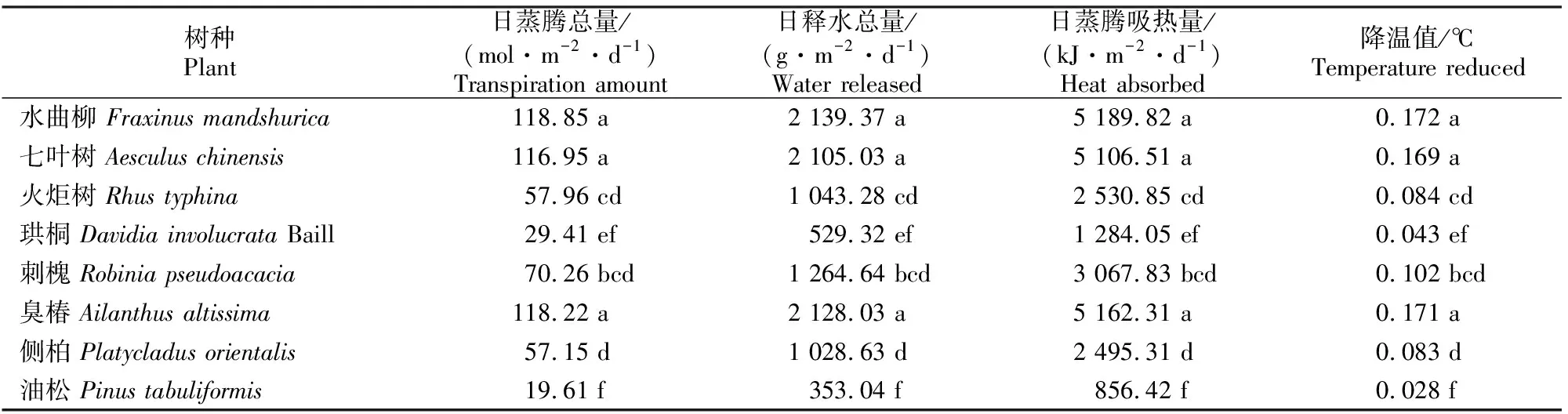
表4 北京市8个绿化树种单位叶面积的日降温增湿量
从表4可以看出,各树种日蒸腾吸热量与释水总量的顺序为:水曲柳>臭椿>七叶树>刺槐>火炬树>侧柏>珙桐>油松。其中以水曲柳的日蒸腾吸热量最高,为5 189.82 kJ/m2,日释水总量2 139.37 g/m2,降温值达到0.172 ℃。最弱的是油松,日蒸腾吸热量仅有856.42 kJ/m2,日释水总量仅有353.04 g/m2,降温值只有0.028 ℃。就各树种蒸腾量而言,除水曲柳与七叶树、臭椿,火炬树与刺槐、侧柏,珙桐与油松无显著差异(P>0.05)之外,其他树种间的日蒸腾总量均有显著差异(P<0.05)。
3 结论与讨论
本研究结果显示,引进树种和乡土树种的Pn在测定时间内呈现规律性变化,即上午09:00-11:00 逐渐增高,中午12:00-14:00达到最高值,然后逐渐降低。除臭椿和油松外,其他6个北京市绿化树种Pn均出现“午休”现象。Pn的大小是由树种本身的特性和环境条件共同决定的,光合作用是植物生长的物质基础,Pn高,则光合产物积累多,本研究中火炬树、臭椿Pn均较高,说明二者有着较强的竞争优势。
本研究采用多元回归分析发现,气孔导度、胞间CO2浓度、光合有效辐射是生长旺季影响树种Pn的主要因子,光合有效辐射的改变可以影响空气温度以及大气饱和蒸汽压差的变化,而大气饱和蒸汽压差的改变又可以影响气孔的开闭,导致气孔导度和胞间CO2浓度发生变化,从而影响树木的蒸腾作用,这些变化又将影响Pn的变化。综合来看,生态因子和生理因子相互作用,相互影响,共同作用于树木[18]。因此在植物生长旺季,应注重光照条件的改善,适时修剪,使叶片之间互不遮挡,以提高光合有效辐射,并加强水肥管理,形成适宜的小气候环境,促进植物的快速生长。
本研究结果表明,臭椿、火炬树的固碳释氧能力较强。植物的固碳释氧量取决于其白天的Pn、夜间呼吸和人工修剪量、凋落物量,其中前两者最为关键[19]。油松和七叶树的单位叶面积日固碳释氧量较其他树种低的原因可能与二者的滞尘能力有关。有研究表明,在自然条件下,大气颗粒物可能在植物叶片上形成一层物理屏障,使其光能利用率降低,从而减弱光合作用[20-21]。虽然火炬树有较强的固碳释氧能力,但因其有着极强的克隆繁殖能力[22],故在园林绿化中要慎用,使其既能为城市绿化服务,又降低其对乡土植被的影响。
与原产地(东三省)水曲柳的固碳释氧量(固碳量5.042 g/m2、释氧量3.659 g/m2)[23]相比,本研究测定数值略高,这可能是因为在试验过程中北京雨水较多,树木在进行光合作用时水分饱和,叶片较为干净,气孔未被附着物覆盖,光合速率较高所致。而对比江苏扬州刺槐的固碳释氧量(固碳量22.39 g/m2、释氧量16.28 g/m2)[10],其值高于本研究测定结果,可能是因为刺槐气孔导度较高,而气孔是植物体与外界交换气体的门户,植物体在适应干旱、半干旱环境的过程中形成的气孔运动形式也是多种多样的,在当地的环境下,树种通过改变气孔导度的大小来影响气孔限制值, 从而来调节体内的水分平衡,使其更有利于叶片光合作用。
植物通过光合作用和蒸腾作用向大气提供了大量的水蒸气,使当地的空气保持湿润,气温降低,在炎热的夏季起到了降温增湿的作用,但不同树种的这一功能具有较大差异。因此,在北京等半干旱区城市的园林绿化过程中,应主要选择七叶树和水曲柳等降温增湿能力较强的植物,将其种植于居民区,使其能够更好地发挥生态效益,而对于侧柏、油松这类通过减少蒸腾耗水来达到适应干旱目的的植物,则更适合作为山区等干旱环境的绿化树种。
[参考文献]
[1] 郎金顶,刘艳红.北京市建成区绿地植物物种来源分析 [J].植物学通报,2008,25(2):195-202.
Lang J D,Liu Y H.Study on built-up area of green sources of plant species in Beijing [J].Chinese Bulletin of Botany,2008,25(2):195-202.(in Chinese)
[2] 孟雪松,欧阳志云,崔国发.北京城市生态系统植物种类构成及其分布特征 [J].生态学报,2004,24(10):2200-2206.
Meng X S,Ouyang Z Y,Cui G F.Composition of plant species and their distribution patterns in Beijing urban ecosystem [J].Acta Ecologica Sinica,2004,24(10):2200-2206.(in Chinese)
[3] 李振蒙,李俊清.植物引种驯化研究概述 [J].内蒙古林业调查设计,2007,30(4):47-50.
Li Z M,Li J Q.Summary of studies on introduction and acclimatization of plant [J].Inner Mongolia Forestry Investigation and Design,2007,30(4):47-50.(in Chinese)
[4] 王豁然,江泽平.论中国林木引种驯化策略 [J].林业科学,1995,31(4):367-372.
Wang H R,Jiang Z P.Summary of introduction and domestication of Chinese forest [J].Scientia Silvae Sinicae,1995,31(4):367-372.(in Chinese)
[5] 唐红军.乡土树种在城市绿化中缺少利用的原因 [J].中国园林,2004,20(6):73-74.
Tang H J.The advantage of local plants and the reasons of lacking of their application in city greening [J].Journal of Chinese Landscape Architecture,2004,20(6):73-74.(in Chinese)
[6] Kabir M E,Webb E.Productivity and suitability analysis of social forestry woodlot species in Dhaka Forest Division,Bangladesh [J].Forest Ecol Manag,2005,212:243-252.
[7] Sendo T,Kanechi M,Uno Y,et al.Evaluation of growth and green coverage of ten ornamental species for planting as urban rooftop greening [J].Jap Soc Hortic Sci,2010,79:69-76.
[8] 韩焕金,周用武,柴一新.城市绿化植物生态效益研究评述 [J].通化师范学院学报,2009,30(12):47-49.
Han H J,Zhou Y W,Chai Y X.Studies on ecological benefits of urban green plants [J].Journal of Tonghua Normal University,2009,30(12):47-49.(in Chinese)
[9] 唐鸣放,白雪莲.城市屋面绿化生态热效应 [J].城市环境与城市生态,2000,13(4):9-10.
Tang M F,Bai X L.Ecological thermal effect of building roof covered by plant in urban area [J].Urban Environment & Urban Ecology,2000,13(4):9-10.(in Chinese)
[10] 徐玮玮,李晓储,汪成忠,等.扬州古运河风光带绿地树种固碳释氧效应初步研究 [J].浙江林学院学报,2007,24(5):575-580.
Xu W W,Li X C,Wang C Z,et al.Carbon fixation and oxygen release of landscaping trees along the Grand Canal in Yangzhou [J].Journal of Zhejiang Forestry College,2007,24(5):575-580.(in Chinese)
[11] 赵 萱,李海梅.11种地被植物固碳释氧与降温增湿效应研究 [J].江西农业科学,2009,21(1):44-47.
Zhao X,Li H M.Study on carbon-fixing, oxygen-releasing,temperature-reducing and humidity-increasing effects of 11 ground cover plants [J].Acta Agriculturae Jiangxi,2009,21(1):44-47.(in Chinese)
[12] 佟 潇,李 雪.沈阳市5种绿化树种固碳释氧与降温增湿效应研究 [J].辽宁林业科技,2010(3):14-16.
Tong X,Li X.Effects of release of oxygen and carbon sequestration and temperature lowering and humidity increasing by 5 afforestation tree species in Shenyang City [J].Liaoning Forestry Science and Technology,2010(3):14-16.(in Chinese)
[13] 韩焕金.城市绿化树种生态功能研究 [M].哈尔滨:东北林业大学出版社,2002:41-70.
Han H J.Studies on ecological benefits of urban green plants [M].Harbin:Northeast Forestry University Press,2002:41-70.(in Chinese)
[14] 许大全,徐宝基,沈允刚.C3植物光合效率的日变化 [J].植物生理学报,1990,16(1):1-5.
Xu D Q,Xu B J,Shen Y G.Diurnal courses of photosynthesis rate on C3 plant [J].Acta Phytophysiologica Sinica,1990,16(1):1-5.(in Chinese)
[15] 马金娥,金则新,张文标,等.蓝果树光合生理生态特性研究 [J].浙江林业科技,2007,27(6):33-36.
Ma J E,Jin Z X,Zhang W B,et al.Study on the eco-physiological characteristics ofNyssasinensis[J].Journal of Zhejiang Forestry Science and Technology,2007,27(6):33-36.(in Chinese)
[16] 陆贵巧,尹兆芳.大连市主要行道绿化树种固碳释氧功能研究 [J].河北农业大学学报,2006,29(6):51-56.
Lu G Q,Yin Z F.A research on the function of fixing carbon and releasing oxygen of afforestation trees along the main road in Dalian city [J].Journal of Agricultural University of Hebei,2006,29(6):51-56.(in Chinese)
[17] 丁振才,黄利斌.常熟虞山森林空气环境效应测定分析 [J].中国城市林业,2004(6):31-32.
Ding Z C,Huang L B.Study on determination of the effects of forest air environment in Yushan [J].Journal of Chinese Urban Forestry,2004(6):31-32.(in Chinese)
[18] 刘海荣,宋 力,张春锋,等.金银忍冬和榆叶梅春季光合特性的比较 [J].沈阳农业大学学报,2006,37(5):707-711.
Liu H R,Song L,Zhang C F,et al.Photosynthetic characteristics ofLoniccramaackiiandPrunustrilobain spring [J].Journal of Shenyang Agricultural University,2006,37(5):707-711.(in Chinese)
[19] 赵 平.退化生态系统植被恢复的生理生态学研究进展 [J].应用生态学报,2003,14(11):2031-2036.
Zhao P.Advances in plant ecophysiological studies on re-vegetation of degraded ecosystems [J].Chinese Journal of Applied Ecology,2003,14(11):2031-2036.(in Chinese)
[20] 陈自新,苏雪痕,刘少宗,等.北京城市园林绿化生态效益研究 [J].中国园林,1998,14(5):57-60.
Chen Z X,Su X H,Liu S Z,et al.Study on urban landscape ecological in Beijing [J].Journal of Chinese Landscape Architecture,1998,14(5):57-60.(in Chinese)
[21] 叶镜中.森林生态学 [M].哈尔滨:东北林业大学出版社,1991:99-101.
Ye J Z.Forest ecology [M].Harbin:Northeast Forestry University Press,1991:99-101.(in Chinese)
[22] 李传文.火炬树:一个值得警惕的危险外来树种 [J].中国水土保持,2004,17(2):31-32.
Li C W.Rhustyphina:An alarming risk of exotic species [J].Soil and Water Conservation in China,2004,17(2):31-32.(in Chinese)
[23] 王摇兵,鲁邵伟,尤文忠.辽宁省森林生态系统服务价值评估 [J].应用生态学报,2010,21(7):1792-1798.
Wang Y B,Lu S W,You W Z.Evaluation of forest ecosystem services value in Liaoning Province [J].Chinese Journal of Applied Ecology,2010,21(7):1792-1798.(in Chinese)
猜你喜欢
能源化工(2022年1期)2023-01-14
绿色科技(2022年17期)2022-09-30
汽车实用技术(2022年7期)2022-04-20
农业工程学报(2022年3期)2022-04-16
湖北农机化(2021年7期)2021-12-07
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
天津农林科技(2018年1期)2018-03-07
中文信息(2016年2期)2016-07-09
